横断山区干旱河谷川滇蔷薇种子休眠与萌发的地理空间差异
2020-11-13周志琼何其华
周志琼,何其华
中国科学院成都生物研究所,中国科学院山地生态恢复与生物资源利用重点实验室,生态恢复与生物多样性保育四川省重点实验室,成都 610041
种子休眠是植物在长期进化过程中形成的抵御不良环境的适应策略,是调节种子萌发最佳时间和空间分布的有效方法[1]。种子休眠程度在不同生态水平如物种间[2]、同一物种的种群间[3-4]、同一种群的不同个体之间[5],甚至同一果实内[6]也存在差异。种群间种子休眠差异是一种普遍存在的现象,主要是由植物自身遗传特征差异以及种子成熟的母体环境所致[7]。目前,对植物自身特征以及生长环境差异引起的种群间种子休眠程度差异的认识仍较为缺乏。
对于同一物种,影响种子休眠的因素可以分为两类:外在环境因素以及植物自身特征。目前,已有大量研究证明不同生境中种子休眠存在差异。如,高温[8-9]、长日照[7]、干旱[10]环境下以及低纬度地区植物种子具有较低程度休眠[11]。不同研究者对于种子休眠与海拔之间的关系得出的结果并不一致,其潜在的机制仍不清楚。大部分研究结果显示,种子休眠随着海拔的升高而加深[8,12];部分学者却得出了不同结论[13]。因此,种子休眠程度与海拔的关系可能与海拔梯度范围以及取样区域的气候环境条件有关。此外,种子休眠程度与植物自身特征,如种子大小、种皮厚度和开花时间有关。与大种子相比,小种子一般具有更深程度的休眠;种皮厚度增加会加深休眠程度[4]。不同种群间种子休眠的差异是由环境因子直接作用,还是通过环境因子引起种子特征变异间接作用仍需进一步的实验证实。
川滇蔷薇(Rosasoulieana)是横断山区干旱河谷广泛分布的乡土灌木,海拔范围1200—3600 m[14],具有浅性生理休眠[15],该物种在横断山区分布覆盖该物种整个海拔分布范围。此外,横断山区具有典型的山地垂直带谱,海拔梯度上环境因子差异显著[16]。因此,川滇蔷薇为研究该区域植物环境适应性提供了理想材料。相对干旱河谷其他灌草植物,川滇蔷薇植株高大,高度2—4 m,冠幅可达5—10 m2。花和果实数量巨大,可为区域昆虫和食果动物提供丰富的食物来源,是干旱河谷植被恢复的重要目标物种。前期研究结果表明:横断山区干旱河谷川滇蔷薇的生长与繁殖特征存在较强的空间差异性[14]。
本研究以横断山区4个典型干旱河谷的18个川滇蔷薇种群为对象,试图阐明在不同流域间以及不同种群间种子休眠程度与萌发的差异;揭示种子休眠与萌发在海拔梯度上的变异格局,进一步分析植物自身因素,如种子大小、种皮厚度和活力,以及环境因素,如温度、降雨、蒸发量与干燥度等对种子休眠与萌发的影响,探讨川滇蔷薇种子萌发特征的生态适应性,为干旱河谷植被恢复以及野生蔷薇资源的开发利用提供科学依据。
1 研究区域
研究区域为横断山区干旱河谷的四个主要流域:岷江流域、大渡河流域、雅砻江流域以及金沙江流域(28°51′37″—32°15′56″N,99°24′37″—103°44′48E)河谷两侧山地,海拔1200—3700 m。该区域年均气温6—16℃,降水量308—720 mm,气候干燥[17]。植被为典型干旱河谷灌草丛,不同地段植被覆盖率差异较大。大部分地段覆盖率为20%—40%之间,部分地段达60%以上。所有植物种类基本上为阳生,大多数种类具有叶片角质层发达、多刺、多毛的特点。土壤类型以山地褐土和山地棕壤为主。本研究调查范围包括岷江流域主河道:汶川绵虒至松潘镇江关,支流黑水河及杂谷脑河,约360 km,海拔1200—2460 m;大渡河流域:马尔康白湾乡至金川德威乡,约400 km,海拔1780—2260 m;雅砻江流域:雅江县的马扎乡至米龙乡,约70 km,海拔2560—3060 m;金沙江流域:乡城县沙贡乡至德荣县松麦镇,约300 km,海拔2700—3700 m[14]。
2 研究方法
2.1 样地设置与数据收集
2011年6月至7月,在川滇蔷薇盛花期开展环境因子调查。在全面踏查的基础上,针对不同流域不同海拔川滇蔷薇的典型分布区域进行样点布设,共布设18个种群样点,包含川滇蔷薇整个海拔分布范围,各样点的布局如图1。记录样点的七个地理位置参数:流域、经度、纬度、海拔、坡度、坡向和距离河道的高度(表 1)。2011年10月1日至10月20日,在川滇蔷薇果实及种子成熟期,开展繁殖性状调查,采集种子萌发实验材料。通常所说的蔷薇种子是指蔷薇瘦果,因此,本研究中所提到的瘦果为萌发单位种子。繁殖性状调查在每个样点的10丛蔷薇(已调查生长特征)中选取5丛,分别在冠丛顶端的北方、西南和东南3个方向,各选取一个当年生小枝,共计15个样本。瘦果重量的测定以小枝作为重复。将瘦果从蔷薇果中分离出来,测定瘦果数量与总重量,瘦果重量=瘦果总重量/瘦果数量。每丛蔷薇随机选取20粒瘦果,TTC染色法测定种子活力[14]。每个种群随机选取20粒瘦果,用单面刀片将瘦果在中间部位横切,测定3个不同部位的瘦果皮厚度,平均值为该瘦果皮厚度。种子萌发实验材料的采集,是在每个样点随机采集20株以上植株的蔷薇果,将瘦果从蔷薇果中分离,每个样点的所有瘦果混合成一个样品,共计18个样品。

图1 横断山区干旱河谷川滇蔷薇分布示意图[14]Fig.1 Distribution of Rosa soulieana in the dry valleys of the Hengduan mountain regionP1 镇江关Zhengjiangguan,P2 红岩Hongyan,P3 较场Jiaochang,P4 飞虹Feihong,P5 渡口Dukou,P6 杨家湾Yangjiawan,P7 巴底Badi,P8 甲居Jiaju,P9小金河Xiaojinhe,P10 孔玉Kongyu,P11 雅江北Yajiangbei,P12 八角楼Bajiaolou,P13 雅江南Yajiangnan,P14 马熊沟Maxionggou,P15 雨洼Yuwa,P16 青德Qingde,P17 正斗Zhengdou,P18 茨巫 Ciwu

表1 横断山区干旱河谷川滇蔷薇取样点的地理位置[14]
2.2 种子萌发实验
将瘦果从蔷薇果中分离、净种后在室内阴干。由于川滇蔷薇具有浅性生理休眠,为了弄清低温层积对不同种群打破休眠,提高发芽率的效果,将瘦果进行8周5℃低温层积,对照为新采集瘦果。层积方法为:将净种后的种子在蒸馏水中室温浸泡24 h,同时将水苔在蒸馏水中浸泡30 m,挤干备用。将种子与水苔按体积比1∶4充分混合,装入自封袋中,在5℃冰箱中放置8周进行层积。新采集种子萌发实验在种子阴干后2周内进行,层积种子萌发实验在层积完成后进行。用5%次氯酸钠(v/v)对萌发种子表面消毒10min,无菌水清洗三次。然后将种子均匀放入装有两层滤纸的培养皿(直径 9 cm),每个处理50粒种子,3个重复,共计5400粒种子。将培养皿放入人工气候箱(20℃,14 h有光/10 h黑暗,光强为30 μmolm-2s-1)。每两天将培养皿随机放置,以避免培养箱位置不同造成的差异。每两天检查一次种子萌发情况,当胚根达到1 mm视为萌发,并将已经萌发的种子从培养皿中去除。萌发实验共持续90 d,实验结束时,将单面刀片将种子切开,完整、白色和硬实的种子视为活力种子。
2.3 数据统计分析
样地环境因子包括年均气温、7月平均气温、年降水量、年蒸发量和干燥度。各因子通过查找样地附近气象站数据获得,对于离气象站较远的样地,结合张荣祖一书中有关气象因子空间变化规律进行推算获得[17]。
发芽率和发芽速度指数计算。
发芽率 GP=萌发种子数/活力种子数量×100%
发芽速度指数 CRG=[Σn/Σ(n×d)]×100%
式中,n为测试活力种子数量,d萌发天数,0为开始萌发时间。
将发芽率和发芽速度指数进行反正弦平方根转换。单因素方差分析ANOVA分析层积对打破休眠,促进种子萌发的作用。协方差分析ANCOVA分析流域对种子发芽率和发芽速度的影响。将发芽率和发芽速度作为固定因子,种群为协变量。偏相关分析种子特征与发芽率和发芽速度的相关关系时,将各环境因子进行控制;偏相关分析环境因子与发芽率和发芽速度的相关关系时,将种子特征进行控制。
3 结果与分析
3.1 川滇蔷薇种子萌发的流域差异
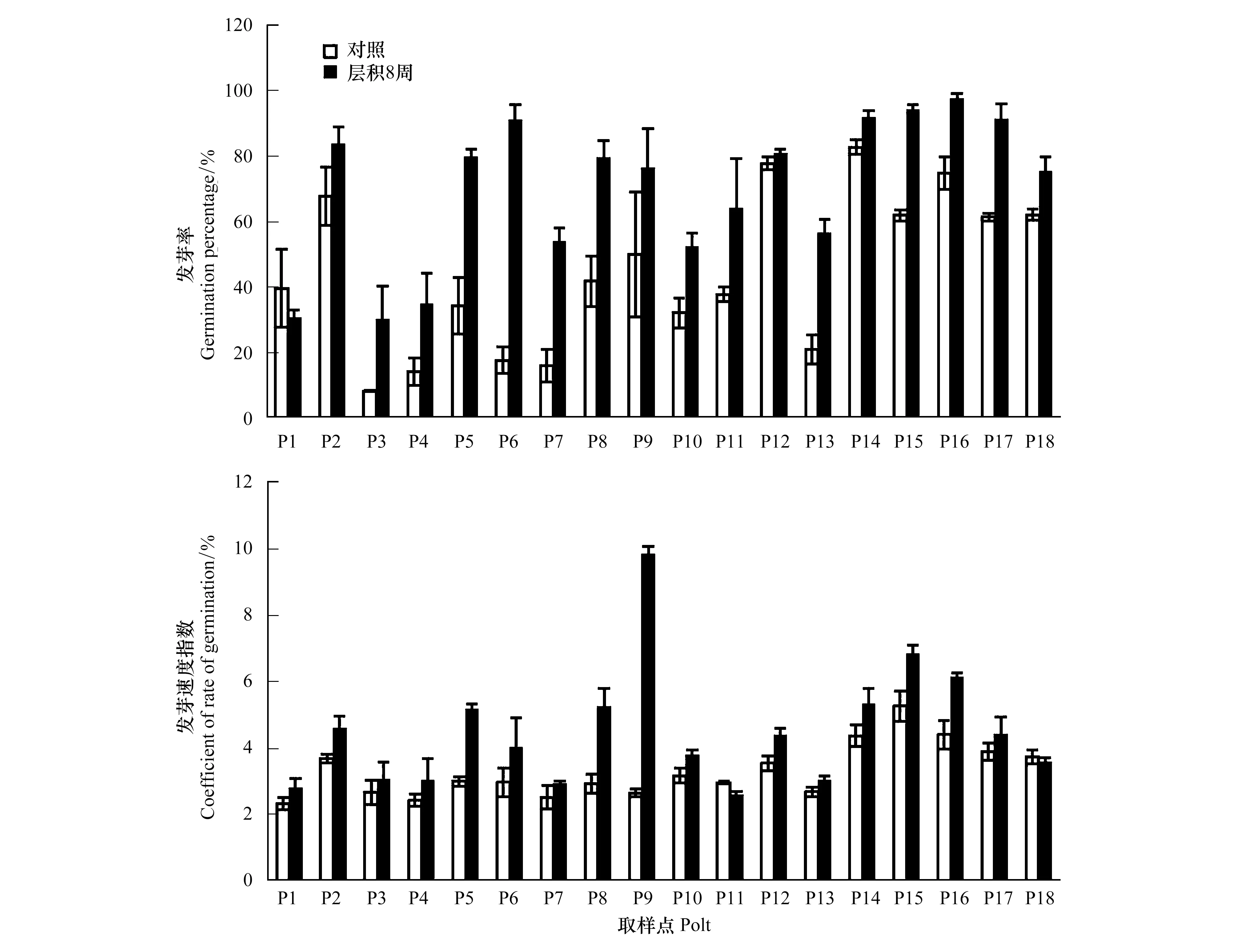
图2 川滇蔷薇在横断山区4个干旱河谷流域中的发芽率和发芽速度Fig.2 Germination percentage and the coefficient of rate of germination of Rosa soulieana in the four dry valleys of Hengduan Mountain region (mean±S.E.)岷江流域P1—P4,大渡河流域P5—P10,雅砻江河流域P11—P13,金沙江流域P14—P18
横断山区干旱河谷18个川滇蔷薇种群种子具有不同程度的休眠。新采集种子发芽率变化范围较大,为15.8%±5.0%至82.7%±2.3%,发芽速度指数2.3%±0.2%至5.3%±0.5% (图2)。不同种群种子发芽率(F=13.87,P<0.0001)和发芽速度(F=8.56,P<0.0001)差异极显著。低温层积8周显著提高了发芽率(F=42.07,P<0.0001)和发芽速度(F=9.42,P<0.007),同时降低了种群间发芽率(F=11.81,P<0.0001)的差异,但增加了种群间发芽速度指数的差异(F=21.21,P<0.0001)。新采集种子发芽率(F=8.94,P<0.0001)和发芽速度(F=3.49,P=0.022)在流域间存在显著差异,具体表现为金沙江流域种子发芽率和发芽速度明显高于其他3个流域,雅砻江流域发芽率高于大渡河和岷江流域种子发芽率,但大渡河和岷江流域种子发芽率之间没有显著差异。8周低温层积降低了流域间发芽率(F=4.06,P=0.012)的差异,层积后种子发芽速度在流域间没有差异(F=1.12,P=0.350)。
3.2 川滇蔷薇种子萌发的海拔差异
川滇蔷薇种子萌发在不同海拔间存在显著差异。新采集种子的发芽率和发芽速度随着采集地点海拔的升高而显著增加(图3),发芽率(r=0.690,P<0.0001)和发芽速度(r=0.610,P<0.0001)与海拔呈显著正相关。低温层积8周提高了休眠程度低的种群种子发芽率和发芽速度,导致发芽率(r=0.232,P>0.05)和发芽速度(r=-0.073,P>0.05)与海拔间没有显著的相关关系(图3)。

图3 川滇蔷薇采集地点海拔对种子发芽率和发芽速度的影响Fig.3 The effect of collection site altitude on germination percentage and the coefficient of germination rate of Rosa soulieana
3.3 瘦果特征对发芽率和发芽速度的影响
偏相关分析结果表明,固定环境因子的影响,分析发芽率和发芽速度与瘦果特征的关系,对于新采集瘦果,只有瘦果皮厚度与发芽速度成显著负相关,而瘦果重量和种子活力与发芽率和发芽速度没有显著相关。层积8周后,瘦果活力与发芽率成显著负相关,其他因子间相关关系不显著(表2)。

表2 川滇蔷薇18个种群萌发参数与瘦果特征的偏相关分析
3.4 环境因子对种子发芽率和发芽速度的影响
固定瘦果特征的影响,分析发芽率和发芽速度与环境因子的关系。对于新采集瘦果,年蒸发量与发芽率和发芽速度之间具有极显著正相关,而降水量与发芽率和发芽速度之间具有显著的负相关。其中年蒸发量与发芽率和发芽速度关系最为紧密。低温层积8周后,各环境因子与发芽速度都没有显著相关,而年均气温、年蒸发量和干燥度与发芽率成显著负相关。

表3 川滇蔷薇18个种群萌发参数与种子特征的偏相关分析
4 讨论
4.1 川滇蔷薇种子休眠与萌发的差异
蔷薇属植物种子一般具有不同程度的生理休眠[18]。本研究中,新采集的川滇蔷薇种子虽发芽率差异较大,但8周低温层积可有效打破休眠,提高发芽率。此外,据观察川滇蔷薇的瘦果皮透水且胚发育完全,按照Baskin和Baskin种子休眠的分类标准[19],18个种群川滇蔷薇种子的休眠类型都属于浅性生理休眠。这与我们前期对岷江干旱河谷川滇蔷薇种子休眠与萌发的研究结果一致[15]。横断山区干旱河谷广泛分布的黄蔷薇具有深度生理休眠[4],而多苞蔷薇具有中度休眠至深度休眠[20]。因此,与同一区域黄蔷薇和多苞蔷薇相比,川滇蔷薇种子休眠程度相对较浅。这可能与其种皮相对较薄,对胚发育的机械阻力较小,以及胚的休眠程度相对较浅有关[20]。
种群间种子休眠的差异是普遍存在的植物适应环境的重要生存策略[7]。本研究中,川滇蔷薇种子休眠与萌发不仅在流域之间存在差异,即使在同一流域的不同种群间也存在显著差异。我们前期研究也表明:18个川滇蔷薇种群的生长和繁殖存在显著的地理空间差异[14]。因此,川滇蔷薇在干旱河谷多变的环境条件中,可以根据自身的生存环境在生长、繁殖以及种子休眠与萌发等方面进行自我调节,以适应环境条件而获得最高适合度。这可能是该种植物适应横断山区干旱河谷多变环境条件的一种生存策略。
4.2 种子自身特征对种子休眠差异的影响
种子自身特征以及植物生长环境条件共同作用引起种子休眠的空间差异[4,7- 9]。对于生理休眠种子,种皮的机械限制以及胚生理休眠是影响种子休眠的两个最重要因素[19]。在蔷薇属植物中,坚硬的瘦果皮在种子生理休眠中起着重要作用[20-22]。本研究中,川滇蔷薇种子发芽速度与瘦果皮厚度成显著负相关,也证明了坚硬瘦果皮对种子萌发的机械阻碍作用。这也与我们前期在同一地区研究中多苞蔷薇种子休眠程度随着瘦果皮厚度增加而加深的结果一致[4]。但低温层积8周后,川滇蔷薇的发芽速度与发芽率与瘦果皮厚度之间没有显著相关性,这可能与川滇蔷薇瘦果皮较薄,休眠程度浅,层积后瘦果皮的限制作用以及胚生理休眠基本解除有关。
同一物种的不同种群,大种子种群一般具有更高的发芽率,且其发芽速度更慢。这是因为大种子需要更长时间吸收萌发所需要的水分[2,23]。但本研究中,新采集川滇蔷薇种子发芽率和发芽速度与瘦果重量没有显著相关。这可能是因为蔷薇种子的休眠机制复杂,受到蔷薇瘦果皮机械阻力、蔷薇果肉及种子内抑制物质和胚生理休眠等多种因素综合影响[4,20],而种子大小对休眠程度的影响相对较小。
4.3 环境因子对种子休眠差异的影响
多种地理空间要素,如经度、纬度以及海拔等导致种子休眠程度的空间差异。地理空间的差异归根结底是环境因子如温度、水分、蒸发、光照等差异。本研究中,对于新采集的川滇蔷薇种子,蒸发量是最重要的环境影响因素,其次为降水量,而年均气温、7月均气温和干燥度与发芽率没有显著相关。与温度、降雨以及干燥度相比,蒸发量是更为综合的指标,受温度、风和空气干燥度等共同影响。一定地段内,一般温度越高,风越大,空气越干燥,蒸发越强。本研究中蒸发量与发芽率和发芽速度成极显著正相关关系,表明温度越高、风力越强、空气干燥度越高的环境下,川滇蔷薇种子休眠程度越低。在控制实验条件下,温度和水分对种子休眠影响表现为:低温以及水分较好环境条件下产生的种子具有更深程度的休眠[24- 25]。本研究中,川滇蔷薇不同种群种子采集区域范围较广,经度范围为:99°24′36.95″—103°44′47.52″E,纬度范围:28°51′37.18″—32°15′55.56″N,气候、土壤及其他生境要素复杂多变。但本研究中川滇蔷薇种子萌发特征与蒸发量和降雨的关系与控制实验条件下得出的结果一致。本文研究结果也与Fernández-Pascual对地中海地区多年生草本植物Centauriumsomedanum不同海拔(25—1670 m)种群种子休眠与环境因子关系的结果一致,即种子休眠程度与降水量成负相关关系[9]。
海拔是影响种子休眠的重要地理环境要素。迄今为止,还未得出有关种子萌发与海拔关系的一致结论。大部分研究结果显示,高海拔种群种子休眠程度更高,需要更长时间的低温层积以打破休眠提高发芽率[3,8,12,26]。这是因为高海拔地区冬季温度更低且霜冻时间更长。较高程度的休眠可延迟种子萌发时间,避免植物在气温突变的早春萌发而受冻害,是植物对高海拔环境的一种适应策略。部分研究也表明,高海拔种群比低海拔种群具有更高的发芽率和发芽速度[13]。即使在同一区域内采集的种子,不同植物种类种子休眠与海拔的关系也表现不同。Wang 等对青藏高原东缘的5种木本植物的研究发现,Philadelphusincanus,和Berberis vernae 发芽率和发芽速度随着采集地海拔的升高而降低,而Betulautilis和Piceapurpurea却表现出相反的趋势,Berberisdubia种子休眠与萌发与海拔之间没有明显趋势[26]。本研究中川滇蔷薇种子发芽率和发芽速度与海拔成负相关关系。其原因在于:一方面种子休眠与海拔关系受多种因素协同限制而复杂多变;此外,川滇蔷薇采集区范围广,经度、纬度和海拔交互变化,进一步加强了种子休眠与海拔关系的不确定性。总的来看,种子休眠与海拔的关系是复杂多变的,与物种本身特性,种子采集区域范围大小,海拔梯度范围等密切相关。
