植物根际土壤酶对重金属污染的响应机制研究综述
2020-11-13蒲生彦王宇陈文英刘世宾
蒲生彦,王宇,陈文英,刘世宾
1. 成都理工大学,地质灾害防治与地质环境保护国家重点实验室,成都 610059 2. 成都理工大学生态环境学院,国家环境保护水土污染协同控制与联合修复重点实验室,成都 610059 3. 中国环境科学研究院,环境基准与风险评估国家重点实验室,北京 100012
随着现代工业的迅猛发展,土壤重金属污染问题日益严重。2014年,由中国环境保护部和国土资源部联合发布的全国土壤污染状况调查公报显示:土壤污染点位综合超标率为16.1%,全部超标点位中无机污染物点位占比高达82.8%[1]。土壤重金属污染在我国南部省份普遍存在,西南地区重金属背景浓度明显高于其他省份;Cd和As作为主要的污染元素,在各省的平均浓度范围分别为36.5~112.9 mg·kg-1和5.8~31.1 mg·kg-1[2]。约有1 000万hm2耕地土壤被重金属污染,每年因此造成的粮食产量损失约为1 200万t[3]。重金属污染影响范围广、危害大,具有隐蔽性和滞后性,因此,运用合适的表征手段来快速识别和评价土壤重金属毒性及其生态效应有着十分重要的意义。
土壤酶是一种高分子活性蛋白质,能催化多种生化反应[4],与土壤中物质循环和能量流动密切相关[5]。酶活性作为表征土壤质量的一项重要生化指标,对环境胁迫的响应极其敏感。土壤酶既能反映土壤生物地球化学循环过程,也能体现土壤微生物的多样性及其他土壤生物特性[6-7],目前,对土壤酶活性的研究主要聚焦于根际土壤酶。“根际”一词最先由Hiltner提出,主要指植物根周围受根功能强烈影响的土壤区域[8]。对于根际范围尚没有明确的定义,研究者普遍将其定义为植物根表面0.5~4 mm范围内[9-10]。作为植物-微生物-土壤相互作用的主要界面,植物根际是能量流动和物质代谢最活跃的部位之一,其酶活性大于非根际土壤[11]。根际的酶活性反映了植物和微生物之间的相互作用,是一项监控微生物活性、群落组成及功能变化的敏感指标[12]。由于酶活性对环境变化的敏感性,重金属与土壤酶活性的关系已成为近年来讨论的热点话题。特别地,研究植物根际土壤酶对重金属污染的响应机制对于揭示土壤及植物对重金属的响应机理有重要意义。
本文基于大量文献调研,较系统地回顾和总结了植物根际土壤酶对重金属污染响应机制相关的研究进展,在此基础上探讨了存在的问题,并对未来研究做出展望,以期对土壤重金属污染生态效应研究提供一定的参考。
1 根际土壤酶活性及主要影响因素(Rhizosphere soil enzyme activity and its main influencing factors)
根际土壤酶是微生物和植物根系向根际土壤环境中释放的蛋白质,其活性在根际的分布呈现出一定梯度。根际土壤酶活性是非根际土壤的1.3倍~2倍,大多数酶的根际范围为1~3 mm,且酶活性随着根系距离的增加而降低[9]。根际土壤酶活性的空间分布一般采用原位酶谱法[13]或土壤切片法[14]进行研究。相比破坏性取样,可视化技术能对土壤中的分子物质进行原位定量分析,对土壤的扰动较小,近些年来得到了广泛关注。根际微生物是指在根际土壤范围内生长繁殖的微生物,以根际细菌、根际真菌及古生菌为主[15]。植物根际中的微生物数量和活性均高于非根际土壤,这种现象被称为“根际效应”[16]。土壤中的生物种类能影响根际土壤酶的活性,此外,土壤理化性质、营养物质也是重要的影响因素。
1.1 生物种类
植物根际土壤酶活性的差异会因植物种类的不同而不同。例如,小麦(TriticumaestivumL.)根际土壤中蛋白酶、磷酸酶、转化酶和接触酶的活性均低于三叶草(TrifoliumrepensL.);与其他作物相比,豆科植物根际土壤中脲酶活性更高[17]。不同的根际微生物能分泌不同种类的酶,作为根际环境中的重要组成部分,植物根系也能分泌H+、酶、氨基酸以及其他有机物质[18]。因此,根际微生物和植物根系分泌物也是影响根际土壤酶活性的重要因素。
大量研究表明,根际土壤酶的主要来源包括:(1)微生物分泌。微生物细胞是土壤酶的主要来源之一[11],一些酶可由根际细菌或真菌在细胞内合成并向外分泌。土壤中90%~95%的酶由微生物直接分泌[19]。(2)植物根系分泌。根际土壤中有少部分酶来源于植物根部分泌[20]。(3)动物排泄物。经动物排泄释放的酶可转入根际土壤。表1列举了根际土壤酶的主要来源及部分酶的种类[21-29]。
1.2 土壤理化性质
温度、湿度和pH作为土壤重要的理化性质,其变化也会对根际土壤酶的活性产生影响。土壤中pH的变化与根系分泌的H+和OH-有关。植物根系分泌H+和OH-以激发养分、保持根表面电化学势平衡[18]。经植物根系分泌的H+和OH-分布于根表2~3 mm的范围以内[30],这一过程改变了植物根周围pH值,使pH值随距离的增加呈梯度分布。质子化是植物根系活动的重要过程,其产生的氢离子-三磷酸腺苷酶(H+-ATP)可为根介质提供渗出有机阴离子和氨基酸所需的反离子,pH值的变化影响了质子化过程,进而对H+-ATP酶的分泌产生影响[31]。研究表明,土壤温度的升高、土壤环境持续湿润或干燥,均将改变土壤微生物的群落组成,并可能增加微生物的生物量和酶的活性[32],低土壤湿度会明显抑制β-葡萄糖苷酶的活性,随着湿度增加,其酶活性也随之上升[33]。Ge等[34]研究发现,酶活性对温度的敏感性较低,高温通常会提高酶的活性。
可见,土壤酶活性对温度和水分较为敏感,过高或过低的温度和湿度均会影响酶的活性。pH作为土壤重要的理化性质之一,其变化也会对相应酶的活性产生影响。
1.3 营养物质
C、P、N和S是土壤中动植物生理代谢活动所需的重要营养元素,由于它们含量或比值的不同,也会给根际土壤酶活性带来不同的影响。
P的含量和C/P值均会改变植物的根际范围,进而影响根际土壤酶的活性。磷酸酶与P循环有关,Razavi等[35]利用酶谱法分析了线虫对羽扇豆(LupinusmicranthusGuss)根际酶活性的影响,结果表明,相比正常植株,受感染植株对P的需求显著增强,导致根系周围磷酸酶的根际范围扩大了1 mm。Wei等[36]发现纤维素的施加增加了酸性和碱性磷酸酶的根际影响范围,这是因为纤维素的添加使C/P值变大,从而影响土壤有机质磷的矿化过程。β-葡萄糖苷酶与C循环密切相关,添加C元素能提升其活性[37]。脲酶与N循环有关,研究表明,施肥增加了土壤中C、N、P和S元素的存储能力,使脲酶的催化活性也得到增强[38]。
综上所述,营养元素的缺乏或增加都是影响根际范围的重要因素,不同营养元素的比值也会对根际土壤酶活性造成影响,致使酶活性发生相应的变化。
2 植物根际土壤酶对重金属污染响应的研究现状(Research status of plant rhizosphere soil enzyme response to heavy metal pollution)
重金属的浓度及污染类型会影响根际土壤酶的活性,重金属在根际土壤中的过量累积会引发环境污染问题,对植物和微生物均会产生巨大危害,人体健康甚至也会受到威胁。目前,重金属污染类型主要包括:单一重金属污染、复合重金属污染、重金属-有机物复合污染、纳米重金属及其氧化物污染,不同的污染类型导致的酶活性变化也存在差异。
2.1 单一重金属污染
单一重金属对受试植物或微生物理化性质的影响是实验研究的重点问题。不同种类的重金属具有不同的生态毒理效应,根际土壤酶活性对重金属污染的响应也会因重金属浓度和种类的不同而发生改变。
根际土壤酶活性对重金属浓度的响应具有“低浓度促进,高浓度抑制”的特点。研究表明,低浓度锌的添加能增强H+-ATP酶的水解和转运活性[31]。杨良静等[39]通过根袋盆栽试验,发现低浓度的Cd对根际土壤中蔗糖酶和脲酶活性均有显著促进作用,随着其浓度的增加酶活性逐渐降低。当Cd的施加浓度为30 mg·kg-1时,蔗糖酶的活性抑制率达到7.2%。邓代莉等[40]通过研究外源Pb污染对紫色土中微生物酶活性的影响,发现森林和耕地土壤中β-葡萄糖苷酶及酸性磷酸酶的活性与Pb外源添加量和有效态Pb呈显著相关性。这说明在一定范围内,重金属浓度越高,对根际土壤酶活性的抑制作用越强。
在自然界中,单个污染物质构成的环境污染虽时有发生,但事实上绝对意义的单一重金属污染并不存在,因为污染多具伴生性和综合性。因此,仅考虑一种重金属的污染缺乏一定的现实性,我们需要对多种复合重金属污染情况进行研究。
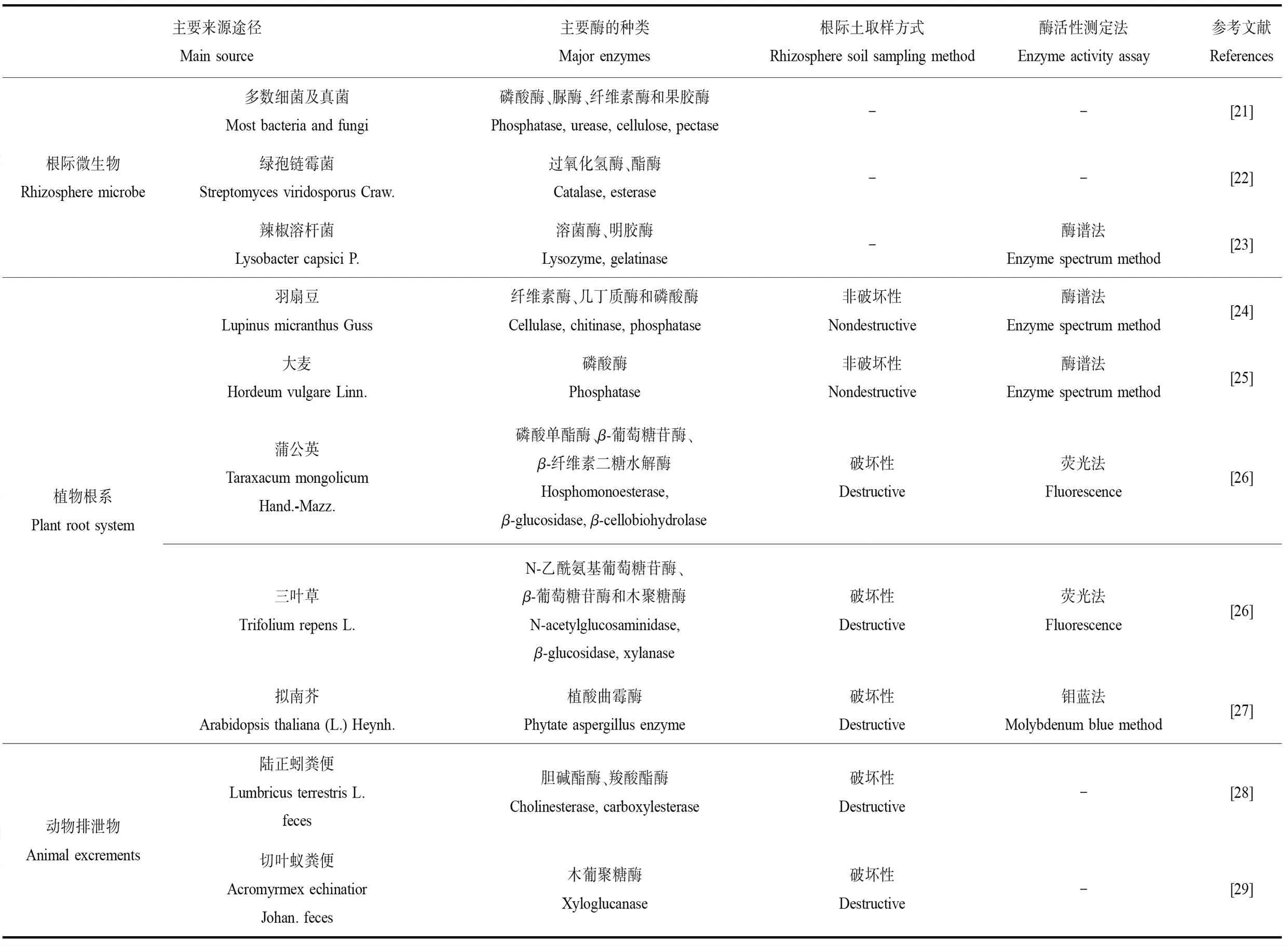
表1 根际土壤酶主要来源及酶的种类Table 1 Main sources and types of enzymes in rhizosphere soil
2.2 重金属复合污染
多种重金属之间的相互作用主要包括2种[41-43]:拮抗作用和协同作用。拮抗作用是指重金属之间通过相互作用表现出的综合毒性小于单种重金属毒性之和;与单一种类的重金属相比,多种重金属间的协同作用表现出了更高的毒性。位点竞争是重金属间产生拮抗作用的原因,这些位点主要包括:代谢系统活性部位或介质中的吸附点,如金属硫蛋白上的结合位点、土壤中的吸附点等;协同作用则是因为不同重金属作用于不同的酶或蛋白,加大了对酶分子的损伤。
研究表明,Cr2O72-和Pb2+易于形成沉淀,使Cr6+和Pb2+反应后综合毒性减弱,这说明Cr和Pb的复合污染可能产生了拮抗效应;对比单独添加Zn对水稻(OryzasativaL.)生长的影响,Cd和Zn的协同作用对水稻的毒害更强[44-45]。Bielińska等[46]将蒲公英(TaraxacummongolicumHand.-Mazz.)种植在受Zn、Pb和Cu等重金属污染的土壤中,相关性基础分析结果表明,土壤中Zn、Pb和Cu的含量越高,脱氢酶活性越低,且根际土壤中的脱氢酶活性高于非根际土壤。这说明,在复合重金属的胁迫下,根际土壤酶活性失活率小于非根际土壤。贾夏等[47]通过对冬小麦幼苗根际土壤氧化还原酶活性的研究,发现低含量的Pb能减轻Cd对多酚氧化酶活性的影响,而加剧Cd对脱氢酶活性的抑制效应。
除重金属复合污染外,自然界中也常伴随有机物污染现象,如铅和芘引发的土壤污染等。讨论重金属-有机物复合污染问题,对于土壤生态毒理学研究有着重大的意义。
2.3 重金属-有机物复合污染

研究表明,芘-铅复合污染胁迫增加了藨草(ScirpustriqueterLinn.)根系相关氧化还原酶的分泌,这些酶与污染物的降解密切相关[50]。Gao等[51]研究发现,在铯和邻苯二甲酸二丁酯(DBP)复合污染的土壤中,幼苗期小麦根际土壤脱氢酶、脲酶和酸性磷酸酶活性增加,苯酚氧化酶和β-葡萄糖苷酶活性被显著抑制,且随着DBP浓度的增加,抑制作用明显加强。后期随着小麦幼苗的生长,根际土壤酶活性得以恢复。
对复合污染的研究是目前环境科学发展的重要方向之一。因此,揭示植物根际的物质循环和能量流动等过程受复合污染影响的程度对于解决生态安全问题、充分评价各类污染物质的迁移转化行为均具有重大意义。
2.4 纳米重金属及其氧化物污染
随着在工、农业等领域的广泛应用,由纳米金属及其氧化物引起的环境问题也日益增多。土壤是人工纳米材料在环境中主要的汇,纳米材料因其独特的理化性质可能会给生态环境带来潜在影响,从而诱发人体健康风险[52]。同种纳米材料对不同土壤中酶的活性影响不同,例如,通过向玉米(ZeamaysL.)地黑土及盐碱土中分别添加纳米Fe3O4颗粒,发现盐渍化土壤中过氧化氢酶的活性显著降低,而黑土中磷酸酶的活性没有明显变化[53]。纳米ZnO能提高植株根际锌利用效率,同时增加相关根际土壤酶的活性。Raliya和Tarafdar等[54]将浓度为10 mg·L-1的纳米ZnO溶液喷施于14日龄的豆荚(LegumeL.)植物叶片上,6周后检测到根际酸性磷酸酶、碱性磷酸酶和植酸酶活性分别增加了73.5%、48.7%和72.4%,这说明,低浓度的ZnO纳米颗粒(NPs)促进了豆荚植物根际酶活性。Sillen等[55]发现低浓度Ag纳米颗粒的添加增加了玉米的生物量,根际细菌群落也显著增加,最终导致酶活性也发生了变化。
纳米重金属及其氧化物是一种新型的污染物,因其粒径尺寸较小,为其监测和治理工作带来了很多困难。目前,对于纳米材料影响根际土壤酶活性的研究还较少,很多方面还有待完善。
3 重金属对根际土壤酶活性的主要影响机制(The main influence mechanism of heavy metals on enzyme activity in rhizosphere soil)
重金属对酶活性的抑制只是一种暂时现象。研究表明,幼苗期经铯污染的小麦在生育期后,土壤酶活性逐渐恢复[51]。重金属对根际土壤酶活性的影响机制主要从酶分子、根际微生物和根系分泌物这3个方面进行研究。
重金属对酶分子的作用机制主要包括:(1)重金属与酶分子的疏基、胺基和羧基结合,或占据酶的活性中心,从而抑制酶的催化功能及破坏其结构完整性。Wang等[56]发现长期As污染会改变土壤酶功能的稳定性,轻度As污染土壤比重度As污染土壤具有更高的土壤酶活性。(2)重金属离子作为酶分子的辅基,促进酶活性位点与底物间的配位结合,使酶分子及其活性位点保持一定的专性结构,从而改变酶催化反应的平衡性质和酶蛋白的表面电荷,增强酶活性。研究表明,经外生菌根菌侵染的樟子松(PinussylvestrisLinn.)在Cd的胁迫下,其根际土壤的过氧化氢酶活性随着胁迫程度的加深呈现出先升高后降低的趋势[57]。这可能是因为低浓度Cd的存在促进了酶的活性位点与底物间的配合,使酶蛋白的表面电荷增加,让酶活性得以提升,高浓度的Cd则抑制了这些反应,降低了酶活性。(3)重金属的胁迫下,酶活性未受到影响,即两者不存在专一性对应关系。0.5 mg·kg-1Cd对TPRC2001-1型柱花草(StylosanthesguianensiasSW.)根际土壤脲酶活性有促进作用,而相同浓度的Cd对西卡柱花草根际土壤脲酶活性无显著影响[58]。
根系的代谢活动或微生物的呼吸作用引发了根际环境与外界的气体交换。根际环境中存在着大量的微生物,重金属的胁迫可诱发微生物的氧化应激机制,氧化应激的发生会抑制微生物的细胞质酶活性,进而破坏细胞结构[59-60]。这将对根际土壤酶的活性产生影响。根系渗出物(如氨基酸、有机酸、粘质物和表面活性剂等)能与重金属相互作用,使其毒性减弱[61-62]。这是因为氨基酸通过酯基(—COO)和胺基(—NH)与金属阳离子相结合形成了络合物,降低了金属离子的迁移转化能力。在极酸或极碱性条件下,根系分泌的H+和OH-也会减轻一些重金属离子的毒性,如酸性或碱性pH条件下铁锰缺乏症引起的Al3+的毒性[63]。此外,重金属的胁迫也会使酶的结构遭到破坏。综上,图1为植物根际对重金属污染的响应机制。
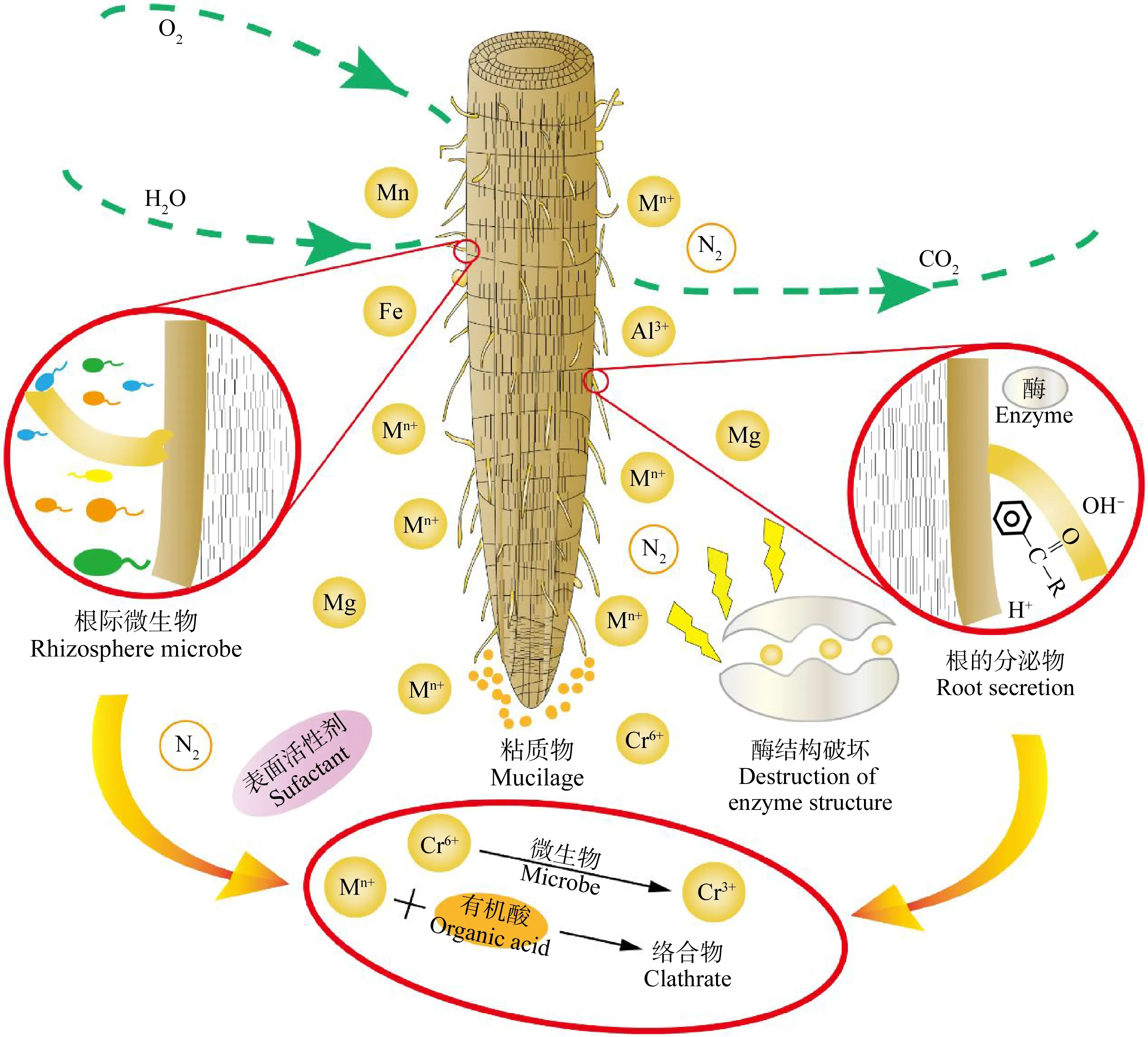
图1 植物根际环境对重金属污染的响应机制Fig. 1 Mechanism of plant rhizosphere environment in response to heavy metal pollution
在评价重金属对根际土壤酶活性影响的过程中,还需考虑多种根系分泌物联合作用的影响。特定种类的植物或微生物能在一定程度上减弱重金属对根际土壤酶活性的毒性,针对重金属污染程度较高的地区,可考虑采用植物修复或植物-微生物联合修复方式。
4 结语与展望(Conclusion and prospect)
关于重金属对土壤酶活性的影响目前已有大量研究,但重金属对植物根际土壤酶活性影响的研究相对较少。大量研究表明,根际土壤酶活性对重金属浓度的响应具有“低浓度促进,高浓度抑制”的特点。重金属-有机物复合污染是自然界中常见的污染类型,对根际土壤酶的影响随植物的生长周期而变化。纳米材料是一种新型的污染物,其对根际酶活性的影响与污染物和土壤类型密切相关。未来相关研究中应关注以下几个方面。
(1)进一步优化开发可视化技术的研究。目前的原位酶谱法在灵敏度、空间和时间分辨率以及可应用性方面具有显著的局限性,无法做到精确定量分析。今后应致力于根际土壤酶活性的量化研究,采用定点取样的方式分析影响植物根系发育和根际特性的其他因素。
(2)考虑更多的现实因素,延长试验周期,进行原位实验研究。实验室的条件无法完全模拟自然界的情况,试验周期较短,酶活性的完全恢复周期尚未明确。今后应研究真实环境下根际土壤酶活性对重金属污染的响应机制,以减少实验模拟数据带来的偏差。
(3)将酶活性与微生物联系起来,建立更多微生物指标。微生物是土壤酶的主要来源,重金属污染也会影响根际微生物的活性。应探索土壤-植物-微生物的互作关系,找出各种酶对应的敏感微生物,为土壤质量评价提供更多生物指标。
◆
