黄河三角洲芦苇根际氨氧化微生物的空间分布
2020-11-13宋延静马兰李萌付娆聂文靖李俊林王向誉郭洪恩
宋延静,马兰,李萌,付娆,聂文靖,李俊林,王向誉,郭洪恩
(山东省蚕业研究所,山东烟台 264002)
黄河三角洲由于濒临渤海土壤盐渍化严重,存在土壤结构性差、肥力低等问题,但开发利用潜力巨大。盐生植物是唯一能够在此环境下正常生长的植物,芦苇是常见的盐生植物之一。在对黄河三角洲开发利用过程中,氮素的施入不可避免。氨氧化过程是硝化作用的第一个反应步骤,也是限速步骤,是全球氮循环的中心环节。氨氧化过程由氨氧化古菌(AOA)和氨氧化细菌(AOB)两类微生物共同完成[1],二者应对各种环境变化的响应不同,因此也被认作环境变化的潜在指示。结合全球变化背景,揭示微生物驱动的氮循环过程尤其是植物根际微生物,对评价与预测河口湿地生态系统服务功能变化、管理决策等至关重要。
根际是土壤、植物、微生物三者相互作用、相互制约的界面和物质转化的活跃区域[2]。滕泽栋等[3]对野鸭湖湿地芦苇根际微生物多样性进行调查发现了磷素转化相关的微生物,但国内外有关芦苇根际微生物的研究主要集中于16S基础之上的研究[4,5],而对根际功能微生物研究较少,尤其是氮循环相关功能微生物[6]。随着微生物生物地理学的发展,距离-衰减关系被发现也广泛适用于微生物群落,空间距离越远的两个样点,其生物群落组成相似度也越低[7-9]。目前有关黄河三角洲地区不同空间尺度下的植物根际微生物研究也不多见。
本研究首次结合荧光定量PCR技术与高通量测序技术对黄河三角洲不同区域(滨州、东营、昌邑、寿光)芦苇根际氨氧化微生物(包括AOA和AOB)的丰度、多样性和群落结构进行比较,研究结果将对湿地生态环境的合理开发利用及保护提供一定的理论基础。
1 材料与方法
1.1 研究区域概况
黄河三角洲是黄河入海口处形成的冲积平原,属于温带季风型大陆性气候,全年平均气温12.3℃,降水量 542.3~842.0 mm,其中约75.5%的降水集中于 6—9月,年蒸发量 1 885.0 mm。该地区土壤以滨海盐土为主,土壤含盐量高[10]。土壤盐分主要为海水氯化物和硫酸盐类。大部分面积的土壤属于盐渍化土壤的范畴,且盐渍化程度高,此外,土壤养分匮乏,有机质含量很低,土壤黏重板结[11,12]。
1.2 样品采集
选择黄河三角洲的昌邑、寿光、滨州、东营4个采样点进行芦苇资源的调查及根际和根周土的收集(抖根法)。采集后的土壤样品存于冰盒(4℃)并立即转移到实验室,用于提取DNA的样品立即放置到-80℃冷冻,用于测定环境因子的样品于-20℃冷冻保存。
1.3 土壤理化性质测定
土壤样品利用0.01 mol/L氯化钙溶液浸提后进行pH值和含盐量测定。土样经冷冻干燥后,取 2.5 g,加入 2 mol/L的 KCl 25 mL,振荡 1 h,过滤备用,采用连续流动分析仪(AutoAnalyzerⅢ)测定铵态氮、硝态氮、亚硝态氮含量。利用元素分析仪(vario MICRO CUBE)进行土壤总氮含量的测定。利用原子吸收分光光度计测定土壤中Na+、K+含量。
1.4 土壤DNA的提取和高通量测序
按照Fast DNA©SPIN Kit for Soil试剂盒说明步骤提取DNA,用分光光度计(NanoDrop 2000c)测定DNA的浓度及纯度。利用AOB-amoA特异性引物 amoA1F/amoA2R[13]和 AOA-amoA特异性引物 arch-amoA-for/arch-amoA-rev[14]分别进行PCR扩增,然后利用Illumina PE250测序平台进行测序(上海凌恩生物有限公司)。
1.5 荧光定量 PCR
AOA和AOB的绝对丰度采用TB Green荧光定量PCR法获得,在BioRad CFX96平台上进行。稀释相应质粒至10-1到10-8,并绘制标准曲线。每个样品设3次重复,并设不加模板的反应管为阴性对照。
1.6 高通量数据分析
高通量数据分析基于Linux平台的QIIME 1.9.0流程,参考数据库为 FGR(version 7.3)。具体流程如下:过滤read尾部质量值20以下的碱基和质控后50 bp以下的read;筛除错配比率大于0.2的拼接序列;用USEARCH软件和GOLD数据库,采用denovo和reference结合的方式去除嵌合体,并选取97%的序列相似度作为阈值(cut off)划分OTU;采用 RDP classifier贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析。
1.7 数据处理
利用SPSS 20统计软件中的单因素方差分析(ANOVA)检验不同采样点芦苇根际土中amoA基因和arch-amoA基因拷贝数的差异显著性;利用mothur计算不同地域芦苇根际氨氧化微生物类群的α多样性指数;利用R语言的vegan包进行基于Weighted UniFrac距离矩阵的聚类树分析。
2 结果与分析
2.1 土壤环境因子分析
由表1可见,4个采样点的土壤 pH值为7.18~7.58,滨州和东营两点的 pH值显著高于寿光和昌邑。东营和滨州两点土壤质地为黏土,含盐量较高,均在5‰以上,尤其东营采样点达到11.22‰;昌邑和寿光采样点的土壤为砂质土,含盐量低,不足 1‰。黄河三角洲土壤总氮在0.018%~0.050%之间,东营和滨州采样点的总氮、总碳含量以及含水量均显著高于寿光、昌邑。芦苇覆盖下的土壤氮以硝态氮为主,含量在2.44~8.13 mg/kg之间;东营采样点的铵态氮含量也较高。寿光、东营、滨州土样的Na+含量高于K+,尤其东营、滨州两点的Na+含量明显高于寿光、昌邑。
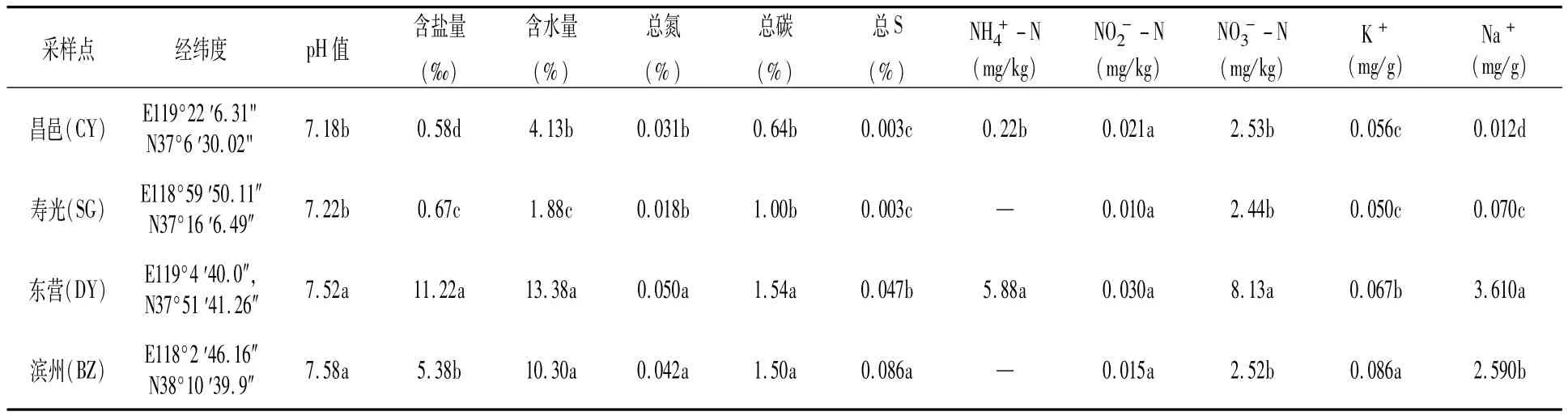
表1 土壤环境因子
2.2 基于amoA基因荧光定量PCR的AOA和AOB丰度比较
定量PCR结果(图1)显示,来源于黄河三角洲4点的芦苇根部土壤中氨氧化微生物均以AOA占主导,AOA的拷贝数比AOB要高出1~2个数量级。
对于AOA,芦苇根际土中arch-amoA基因拷贝数以东营最高,平均值为1.61×108copies/g;其次为滨州,平均值为 1.06×108copies/g;昌邑较低,为7.56×107copies/g;寿光最低,仅为5.77×106copies/g。芦苇根周土中的arch-amoA丰度要显著低于根际土,仍以东营的arch-amoA拷贝数最高,为1.14×107copies/g,昌邑、寿光、滨州 3点的芦苇根周土中arch-amoA拷贝数差异不显著,以昌邑的最低。
对于AOB,除昌邑外,寿光、东营、滨州三点的芦苇根周土中amoA拷贝数都高于根际土,东营的芦苇根周土中amoA拷贝数最高,为6.70×107copies/g;滨州的芦苇根际土中amoA拷贝数最低,仅 9.11×104copies/g。
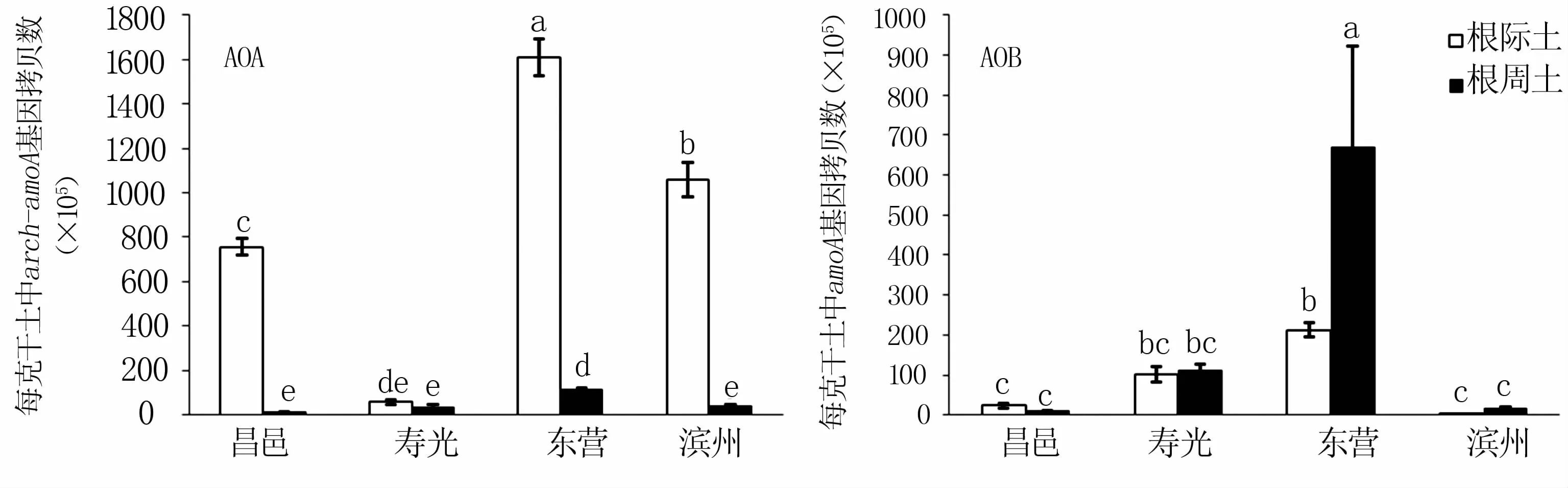
图1 黄河三角洲不同地区间芦苇根际AOA和AOB amoA基因丰度的比较
2.3 黄河三角洲不同地区芦苇根际 AOA和AOB的α多样性
对于AOA,通过质控共获得170 812条高质量的arch-amoA基因序列,划分为342个OTU,每个采样点芦苇根际AOA的OTU数量平均在100~150之间。从表2的Chao1数据可知,各样本的AOA丰富度排序为东营(DY)>滨州(BZ)>昌邑(CY)>寿光(SG);从Shannon指数和Simpson指数来看,各站点AOA多样性从高到低依次是寿光(SG)>滨州(BZ)>东营(DY)>昌邑(CY)。
对于AOB,通过质控共获得172 001条高质量的amoA基因序列,划分为443个OTU,每个采样点芦苇根际AOB的OTU数量平均在23~171之间。从表2的Chao1数据得知,AOB的丰富度昌邑最高;从Shannon指数和Simpson指数来看,AOB的多样性总体表现为昌邑(CY)>滨州(BZ)>寿光(SG)>东营(DY)。

表2 不同采样点芦苇根际氨氧化微生物(AOA和AOB)α多样性指数
2.4 黄河三角洲芦苇根际AOA和AOB群落组成分析
从图2可以看出,从属的水平来看,奇古菌门(Thaumarchaeota)的亚硝化球菌(Nitrososphaera)是昌邑和寿光两地芦苇根际土中参与氨氧化过程的主要AOA类群(>99%);对于滨州和东营,芦苇根际中的AOA类群主要为Nitrosopumilus,其次为Nitrosoarchaeum古菌类群。
昌邑和寿光两地芦苇根际土中参与氨氧化过程的主要 AOB类群为亚硝化弧菌属(Nitrosovibrio)和亚硝化螺菌属(Nitrosospira);东营芦苇根际中的AOB类群较为单一,几乎全为亚硝化螺菌属(Nitrosospira);滨州芦苇根际中除亚硝化螺菌属(Nitrosospira,86.0%)外,亚硝化单胞菌(Nitrosomonas)也占了12.7%,该菌在其它三地芦苇根际含量较低。

图2 黄河三角洲不同地区芦苇根际固氮菌群落结构(属级别)
2.5 黄河三角洲芦苇根际AOA和AOB的Beta多样性分析
基于Weighted UniFrac距离矩阵计算黄河三角洲不同地区芦苇根际AOA和AOB的Beta多样性,结果(图3)表明,不论是AOA还是AOB,东营和滨州芦苇根际氨氧化群落组成以及结构差异较小,寿光和昌邑芦苇根际氨氧化群落结构较小。

图3 基于UniFrac距离矩阵的芦苇根际AOA和AOB聚类树分析
3 讨论与结论
黄河三角洲属于典型的河口湿地生态系统,生态环境脆弱,生物多样性低。芦苇是黄河三角洲地区盐生植被的主要建群物种,占土壤面积的5.38%[15],主要生长在河口湿洼地和滨海沼泽地等地势低洼的地区,群落生境一般有积水,土壤含盐量变化较大[16,17]。
3.1 黄河三角洲芦苇根际AOA和AOB的丰度特征
本研究发现黄河三角洲湿地昌邑、寿光、东营、滨州四地的芦苇根用土氨氧化微生物均以AOA为主,这与国内外大多数河口环境的研究结果相一致[18,19]。有限的氮底物则可能是氨氧化微生物生长的限制因素,AOA在肥力低的环境中贡献较大,主要是因为AOA对底物氨的亲和力远高于AOB[20],因此在低肥力土壤中更具竞争优势。当然,也有例外,在Douro河口和Cochin河口,AOB的丰度显著高于 AOA[21,22],同样我们在昌邑采样点得到的芦苇根周土氨氧化微生物群落中也是AOB丰度较高,分析造成这种特殊现象的原因可能与盐度有关。盐度是解释AOA群落结构模式的主要因素,中等盐浓度更适宜AOA的生长。一般盐渍化土壤偏碱性,养分含量低,不利于土壤微生物的生长和繁殖[23-25]。本研究还发现,黄河三角洲4个采样点中的AOA和AOB属东营芦苇根际最丰富,分析可能与东营较高的总碳、氮含量有关,因为有研究表明总氮是影响土壤中AOA和 AOB群落结构的潜在环境因素[26]。
3.2 芦苇根际AOA和AOB在黄河三角洲不同地区的空间分布特征
本研究发现黄河三角洲4个不同地区芦苇根际的AOA几乎全部来源于奇古菌门(Thaumarchaeota),只是不同地区的所属种类不同,滨州、东营两地的AOA要比寿光、昌邑的多样性要高。其中昌邑和寿光两地芦苇根际AOA优势类群为亚硝化球菌(Nitrososphaera),滨州和东营的芦苇根际AOA优势类群为 Nitrosopumilus、Nitrosoarchaeum、Candidatus Nitrosotenuis。前人研究已证明Nitrosopumilus是海洋 AOA的主要优势种群,Nitrososphaera是土壤AOA的主要优势种群[27-29],我们分析是东营和滨州高盐度的生长环境选择了芦苇根际Nitrosopumilus这一微生物类群。
东营和滨州两地芦苇根际AOB的优势类群是亚硝化螺菌属(Nitrosospira)。Nitrosospira属于β变形菌门的一类化能自养氨氧化微生物,更容易出现在低铵的环境中,即贫瘠的土壤环境可能适宜 Nitrosospira的生长[31,32]。Kowalchuk等也发现Nitrosospira是弱碱性土壤中发挥氨氧化作用的最主要微生物。亚硝化弧菌属(Nitrosovibrio)和亚硝化螺菌属(Nitrosospira)是昌邑和寿光芦苇根际AOB的优势类群,Nitrosovibrio也是一种严格好氧的化能自养氨氧化微生物。戴九兰和苗永君在对黄河三角洲不同盐碱农田生态系统氮循环功能菌群研究中也发现,在小麦-玉米轮作和大豆种植体系土壤中,AOB中Nitrosospira和Nitrosovibrio主导着氨氧化过程[33]。
本研究还发现距离较近的寿光、昌邑两地的芦苇根际AOA和AOB群落结构类似,滨州和东营两地的AOA和AOB群落结构类似。这在一定程度上验证了微生物的距离-衰减关系[7],具体和哪种环境因子相关还需要进行更深入的分析探讨。
