高寒沙区生物土壤结皮覆盖土壤碳释放动态
2020-11-12赵河聚贾晓红李元寿岳艳鹏赵雪彬
赵河聚,成 龙 ,贾晓红,*,吴 波,李元寿,岳艳鹏, 周 虹,赵雪彬
1 中国林业科学研究院荒漠化研究所, 北京 100091 2 中国气象科学研究院, 北京 100081 3 青海共和荒漠生态系统国家定位观测研究站, 共和 813000
生物土壤结皮是由隐花植物(如蓝藻、地衣、苔藓等)和土壤中的微生物通过假根、菌丝体和胞外分泌物等与土壤表层细小颗粒胶结形成的复杂的复合体[1-2]。生物土壤结皮能够适应极端的环境条件,通过自身的代谢方式影响和改变周围环境,在稳定表层土壤、提高土壤抵抗风蚀、水蚀能力、增加土壤肥力和促进土壤发育等方面具有重要的生态功能[2-4]。干旱半干旱地区水资源极度匮乏,植被覆盖度与生物量低,但生物土壤结皮却能广泛分布其中,在部分荒漠生态系统中的覆盖度甚至可达到70% 以上[5],是干旱半干旱地区重要的地表覆盖类型之一。生物土壤结皮能够通过自身所包含的有机成分与生命体的呼吸作用向大气中释放CO2,从而影响土壤碳释放以及该区生态系统的碳循环。
国外有关生物土壤结皮的研究主要分布于美国的莫哈维沙漠、以色列内盖夫沙漠和中欧一些地区[6],国内则主要集中于腾格里沙漠[7-8]、毛乌素沙地[9-10]和黄土高原[11]等温带地区。研究内容多为生物土壤结皮的类型与功能[3,5,12]、生物土壤结皮对土壤呼吸的影响[13-16]、生物土壤结皮呼吸与水热因子间关系等[6,7,17,18]。但有关生物土壤结皮在高寒生态系统的研究相对不足,不同类型生物土壤结皮发育对土壤碳释放的影响亟待研究。共和盆地位于青藏高原东北部,青海南山和巴颜喀拉山之间,属于高寒干旱荒漠和半干旱草原过渡区,是黄河的上游地带,在保持水土、涵养水源、生态保护方面地位十分重要。本研究在具有典型高寒沙区气候特征的青海共和盆地,以广泛分布于人工植被恢复区内的藻类和藓类结皮为研究对象,采用野外原位测量实验方法,对裸地与不同类型生物土壤结皮去除和覆盖土壤碳释放速率的日和生长季动态进行观测,以期能揭示高寒沙区生物土壤结皮发育对土壤碳释放的影响规律及其变化机理,为区域碳平衡核算与建模分析提供一定的数据参考与科学依据。
1 材料与方法
1.1 研究区自然概况
研究区位于青海省共和县沙珠玉乡青海省治沙站沙珠玉试验林场(36°16′N,100°16′E),地处青海共和盆地的中西部地区,属于高寒干旱荒漠和半干旱草原过渡区。海拔高度2871 m,面积为5.28×102km2,土地沙化面积达1.74×102km2,春季风沙活动较多。该地区具有高寒、干旱、辐射强烈等气候特点,年均降水量为246.3 mm,且年内分布极不均衡,降水多集中发生于生长季,以小降水事件为主,年潜在蒸发量约为1716.7 mm。年均温度为2.4 ℃,无霜期在91 d左右,光照充足,年太阳辐射总量为6.62×105KJ/cm2。
受剧烈风沙活动影响,沙珠玉地区一直是青海省土地荒漠化的多发地区之一。为治理荒漠化问题,青海省治沙实验站已经开展了近60年的植被恢复工作,土地荒漠化问题得到了有效的治理,生态环境逐步改善,形成了以灌木为主的“乔-灌-草”复合植被恢复体系,同时大量的生物土壤结皮覆盖地表。该区乔木种主要有小叶杨(Populussimonii)、河北杨(Populushopeiensis)、青杨(Populuscathayana)等;灌木种主要有柽柳(Tamarixchinensis)、柠条锦鸡儿(Caraganakorshinskii)、沙棘(Hippophaerhamnoides)等;草本植物种主要有赖草(Leymussecalinus)、针茅(Stipacapillata)等。本研究选择结皮发育相对较好的1977年植被恢复区为研究样地。样地内土壤为棕钙土和栗钙土等。植被分布多以灌木为主,主要植物种有:柠条锦鸡儿(Caraganakorshinskii),针茅(Stipacapillata)等。样地内生物土壤结皮基本特征和土壤理化性质见表1—3[19]。

表1 研究区生物土壤结皮主要特征
1.2 研究方法
1.2.1样地设置
选取灌丛间发育较好的藻类结皮覆盖土壤和苔藓结皮覆盖土壤(两种类型生物结皮的盖度均保持在90%以上,且远离植株根系分布范围,距植株2 m左右)为研究对象,以裸地为对照。在样地内,设置五个2 m × 2 m的样方(藻类结皮覆盖土壤、藻类去除结皮土壤、苔藓结皮覆盖土壤、苔藓去除结皮土壤和裸地),每种观测类型分别设置三个重复。在每个样方内放置三个PVC土壤环用于土壤碳释放速率测定。放置时,先把生物土壤结皮表层湿润,然后将高10 cm、直径20 cm的PVC环垂直嵌入土壤中,使土壤环上端高出地表约4 cm左右,前期准备工作在实验开展前一个月完成。
1.2.2土壤碳释放速率测定
实验开展于2017和2018年的生长季(6—10月),利用LI-8100土壤碳通量自动测量系统连接土壤呼吸长期监测室8100-103(该气室使用金属材质,闭合时,密闭遮光,在完全黑暗条件下测量生物土壤结皮层呼吸作用和下覆土壤呼吸作用的总和)对裸地与两种类型生物土壤结皮去除和覆盖土壤碳释放速率进行测定。测定内容包括日变化与月变化测定(每月测定一次),日变化测定:每月选取晴朗天气(测定前3天无降水事件发生),观测时间为8:00至次日8:00,每2 h观测一次,每次观测3 min,包括测量时长90 s,测量前与测量后气体平衡时间各45 s;月变化测定:每月选取晴朗天气(测定前3天无降水事件发生),观测时间为9:00—11:00,在同一时段内,将所有土壤环内样本均观测一遍,每次观测3 min,包括测量时长90 s,测量前与测量后气体平衡时间各45 s。
R土壤呼吸增量=R生物土壤结皮覆盖土壤-R裸地
R生物土壤结皮层=R生物土壤结皮覆盖土壤-R生物土壤结皮去除土壤
1.2.3环境因子测定
近地面空气温湿度观测采用VP-3水汽压温湿度和大气压传感器,0—5 cm层土壤温湿度观测采用GS3水分、电导率、温度传感器,5—10 cm层、10—20 cm层土壤温湿度观测采用5TM 土壤水分、温度传感器,并通过EM50土壤温湿度数据采集器进行数据收集。日变化测定:所有观测装置的设定时间均为30 min记录一次数据,月变化测定:所有观测装置的设定时间均为1 min记录一次数据。降水资料通过附近气象站获得。
1.2.4数据处理与分析
日累积碳释放量计算公式如下[20]:
Cdaily=(R8+R10+…+R6)×24×3600×10-6
式中,Cdaily为日累积碳释放量(以CO2释放量计,g CO2m-2d-1),R8+R10+…+R6分别为8:00、10:00…到次日6:00的平均碳释放速率(μmol m-2s-1)。
月累积碳释放量=日累积碳释放量×每月天数
年生长季累积碳释放=生长季各月累积释放量相加
采用 Excel 2010进行数据处理, SPSS 19.0 进行双变量相关性分析和主成分分析。并在Origin 2017中绘制相关图形。
2 结果与分析
2.1 各观测类型碳释放速率日动态
裸地和不同类型生物土壤结皮去除、覆盖土壤的碳释放速率日动态变化与表层(0—5 cm)土壤温度变化规律一致,均呈“单峰”曲线(图1)。8:00开始,各观测类型碳释放速率均迅速增高,在13:00左右,生物土壤结皮覆盖土壤碳释放速率达到日最大值;在15:00左右,裸地和生物土壤结皮去除土壤碳释放速率达到日最大值;土壤温度在16:00左右达到日最大值;之后碳释放速率均迅速下降,20:00—6:00,碳释放速率均逐渐稳定,且在2:00—6:00之间出现最小值。观测期间,同一年内,各观测类型日最大碳释放速率变化规律:苔藓结皮覆盖土壤>藻类结皮覆盖土壤>苔藓结皮去除土壤>藻类结皮去除土壤>裸地。日均碳释放速率变化规律表现为:苔藓结皮覆盖土壤>苔藓结皮去除土壤>藻类结皮覆盖土壤>藻类结皮去除土壤>裸地。不同年份间,2018年各观测类型日最大碳释放速率和日均碳释放速率均大于2017年,且各观测类型日最大碳释放速率和日均碳释放速率最大值均出现在8月。

图1 裸地与不同类型生物土壤结皮去除和覆盖土壤碳释放速率(以CO2释放速率计)与表层(0—5 cm)土壤温度日动态
2.2 各观测类型碳释放速率生长季动态
各观测类型碳释放速率均存在明显的生长季动态,其年内生长季动态均呈“单峰”曲线。生长季内,各观测类型碳释放速率最大值均出现在8月,最小值均出现在10月(图2)。同一年内,各观测类型生长季平均碳释放速率变化规律为:苔藓结皮覆盖土壤>苔藓结皮去除土壤>藻类结皮覆盖土壤>藻类结皮去除土壤>裸地(图2)。不同年份间,2018年各观测类型生长季平均碳释放速率均大于2017年。

图2 裸地与不同类型生物土壤结皮去除和覆盖土壤碳释放速率(以CO2释放速率计)生长季动态
2.3 各观测类型生长季累积碳释放量
随结皮发育,同一年内,生长季各观测类型碳释放量变化规律为:苔藓结皮覆盖土壤>苔藓结皮去除土壤>藻类结皮覆盖土壤>藻类结皮去除土壤>裸地(图3)。在2017年,藻类结皮覆盖土壤碳释放量较裸地增长22.07%,增量为14.35 gCO2/m2,其中,藻类结皮层碳释放量占增量的67.60%;苔藓结皮覆盖土壤碳释放量较裸地增长85.61%,增量为55.63 gCO2/m2,苔藓结皮层碳释放量占增量的25.76%;在2018年,藻类结皮覆盖土壤碳释放量较裸地增长139.37%,增量为137.87 gCO2/m2,藻类结皮层碳释放量占增量的69.09%;苔藓结皮覆盖土壤碳释放量较裸地增长290.53%,增量为287.39 gCO2/m2,苔藓结皮层碳释放量占增量的45.59%。不同年份间,2018年各观测类型土壤碳释放量均大于2017年,2018年各观测类型(裸地、藻类结皮去除土壤、藻类结皮覆盖土壤、苔藓结皮去除土壤和苔藓结皮覆盖土壤)土壤碳释放量增长率分别为52.22%、103.28%、198.50%、140.21%、220.30%。
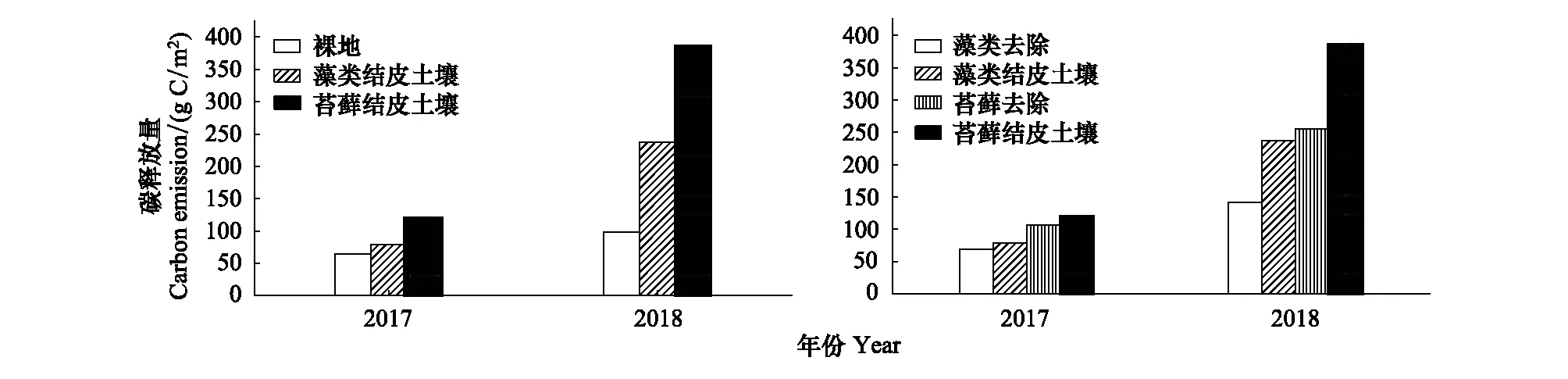
图3 裸地与不同类型生物土壤结皮去除和覆盖土壤生长季累积碳释放量(以CO2释放量计)差异
2.4 各观测类型碳释放速率与温度和水分间的关系
相关性分析表明(表4),各观测类型土壤碳释放速率与空气和不同深度土壤温度均表现出显著(P<0.05)或极显著正相关关系(P<0.01);除与近地面空气湿度呈现出负相关或极显著负相关关系外(P<0.01),各观测类型土壤碳释放速率与不同深度土壤含水量均表现出显著(P<0.05)或极显著正相关关系(P<0.01)。可得出,0—5 cm层土壤温度与含水量对各观测类型土壤碳释放速率的影响最大。通过主成分分析(图4)发现,影响本区生物土壤结皮土壤碳释放日动态的第一主成分主要为温度,第二主成分主要为土壤含水量,温度对土壤碳释放速率变化的贡献率为48.89%,水分的贡献率为31.82%。

图4 裸地与不同类型生物结皮去除和覆盖土壤碳释放速率与温度、水分间的主成分分析
3 讨论
生物土壤结皮作为一种特殊的生物与土壤的复合体,存在于土壤与大气之间的界面层,在一定程度上影响着土壤呼吸,是干旱、半干旱区生态系统CO2通量的影响因子之一[21]。本研究发现生物土壤结皮去除和覆盖土壤碳释放速率日动态和生长季动态特征与裸地一致,均呈“单峰”曲线,这与其他区域研究结果一致[11,22,23]。但生物土壤结皮的存在使土壤碳释放速率日峰值出现时间提前2 h左右,且早于土壤温度日峰值3 h左右。在毛乌素沙地的研究表明,生物土壤结皮覆被区的土壤呼吸日峰值出现时间为11:00左右,早于土壤温度日峰值1—5 h左右[9]。这可能是由于生物土壤结皮中的微生物组分对水分的响应程度不同。在严重受水分胁迫的荒漠生态系统,较低的水分含量使苔藓和藻类结皮的微生物活性更容易受到限制。相对于土壤,苔藓与藻类结皮则需要更多的水分去满足其生理活性。在毛乌素地区,日尺度土壤呼吸值的变化主要与土壤含水量有关,随土壤含水量的减少,土壤呼吸速率也相应减小[24]。故受土壤水分限制,土壤呼吸日最大值出现时间提前。但在高寒沙区,本研究发现温度是影响土壤呼吸的主要因素,温度对土壤呼吸变化的贡献率可达48.89%。研究区内昼夜温差较大,可能对表层土壤中的微生物活性与土壤酶的催化作用产生了影响,从而影响了土壤的碳释放速率。另一方面,这也可能是由于所测量的土壤呼吸值没有对应最佳深度的土壤温度,或是土壤自养呼吸与异养呼吸对环境因子的响应程度不同所导致的[18]。这还需要我们进一步去研究说明。
生物土壤结皮的存在促进了土壤碳释放,随结皮发育程度增高,促进效果越明显。但藻类结皮对土壤碳释放的促进作用主要来源于其结皮层的贡献,而苔藓结皮主要是由于其下层土壤呼吸对土壤碳释放的促进作用更大。一方面,在干旱、半干旱地区,水分有效性一直是主要限制因子。随着生物土壤结皮的发育,藓类结皮的生理活性高于藻类结皮,其表层具有密集丛生的苔藓植物和柔韧的茎叶分布,能在一定时间内,减缓水分的下渗与蒸发,增加了表层土壤水分的有效性[25]。这些表层土壤水分的有效性对生物土壤结皮生理活性的激发尤为重要[26]。另一方面,由于两种类型生物结皮下部土壤质地、养分状况、稳定性和微生物种类、数量、活性等均存在差异[27-28],发育程度越高的结皮下土壤粘粉粒含量与有机质含量均比较高(表2、表3),且微生物量的种类、数量与生物量也相应增多[29],更利于土壤的碳释放,故生物土壤结皮发育程度越高,其碳释放量增加越大。在生长季内,苔藓结皮覆盖土壤下5—10 cm的土壤含水量显著大于0—5 cm的土壤含水量,而藻类结皮覆盖土壤下5—10 cm与0—5 cm的土壤含水量没有显著差别(图5)。水分条件的改变,很大程度地影响了微生物的活性[30],改良了苔藓结皮下层土壤的发育条件,更利于下层土壤中微生物的呼吸作用,对土壤碳释放的整体促进作用更强。相反,藻类结皮的表层与浅层土壤水分却差别不大,所以微生物数量更多的结皮层对土壤碳释放的贡献率更高。本研究还发现,降雨量的增加能够促进裸地与生物土壤结皮去除和覆盖土壤的碳释放作用。在观测期内,相对于本地区近30年年均246.3 mm的降水量,2018年增加了51.16%的降水量,2017年却减少了8%,且小降水事件(降水量<10 mm)的频度均为85.90%左右(表5)。而生物土壤结皮对水分变化十分敏感。在相对干旱的环境中,能使自身脱水进入休眠,而在相对湿润的条件下,又能迅速激发其生理活性使其恢复正常[31]。因此水分条件的改变对苔藓和藻类结皮覆盖土壤碳释放的影响效果十分明显。同时,降水的增多,还会协同增多土壤中微生物的生物量,促进微生物的活动,加速底物分解,并导致土壤碳释放量的增加[32]。在本区的模拟降水研究中也发现小降水对两种类型生物结皮覆盖土壤的碳释放具有极显著的激发作用,随降水量的增加,其碳释放量也相应增加[25]。也有研究表明,中等程度的降水能够有效地增大土壤的碳释放[33]。但极端降水情况则会抑制生物土壤结皮覆盖土壤的碳释放。这可能是由于过大的降水(强度)易堵塞土壤的通透性,从而降低土壤的碳释放[7]。然而本区生长季以小降水为主,更有利于促进土壤的碳释放作用。
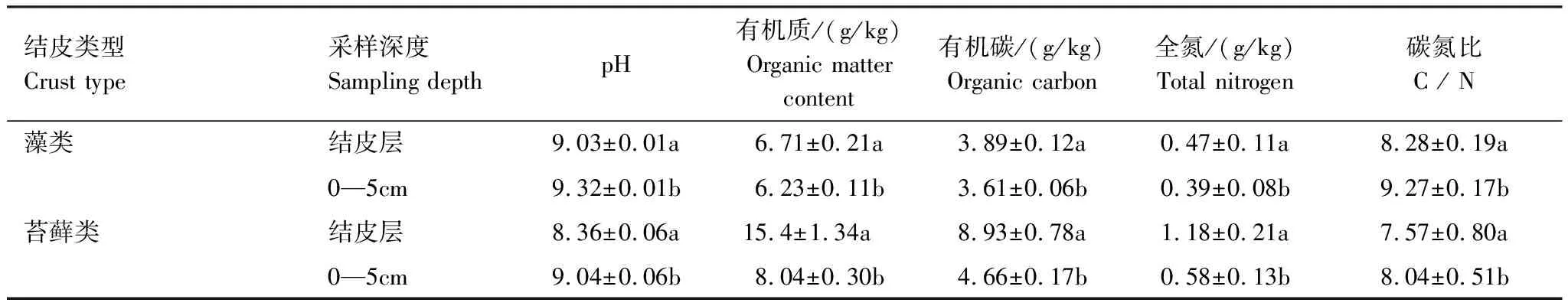
表2 研究区生物土壤结皮土壤基本特征
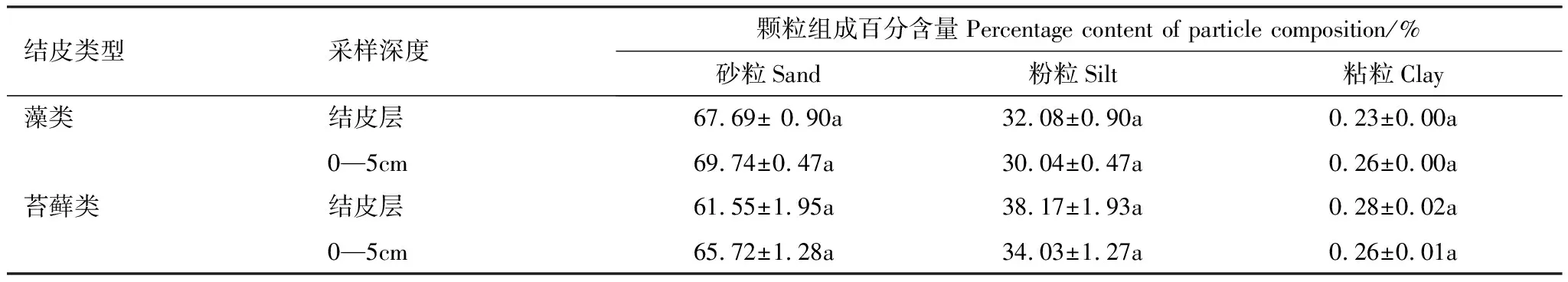
表3 研究区生物土壤结皮土壤机械组成特征

表4 裸地与不同类型生物土壤结皮去除和覆盖土壤碳释放速率与温度和水分间的关系
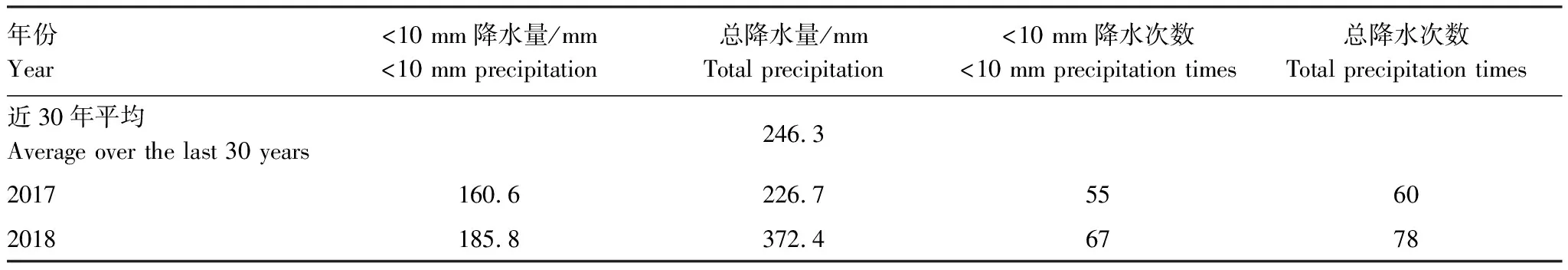
表5 生长季内降雨事件统计
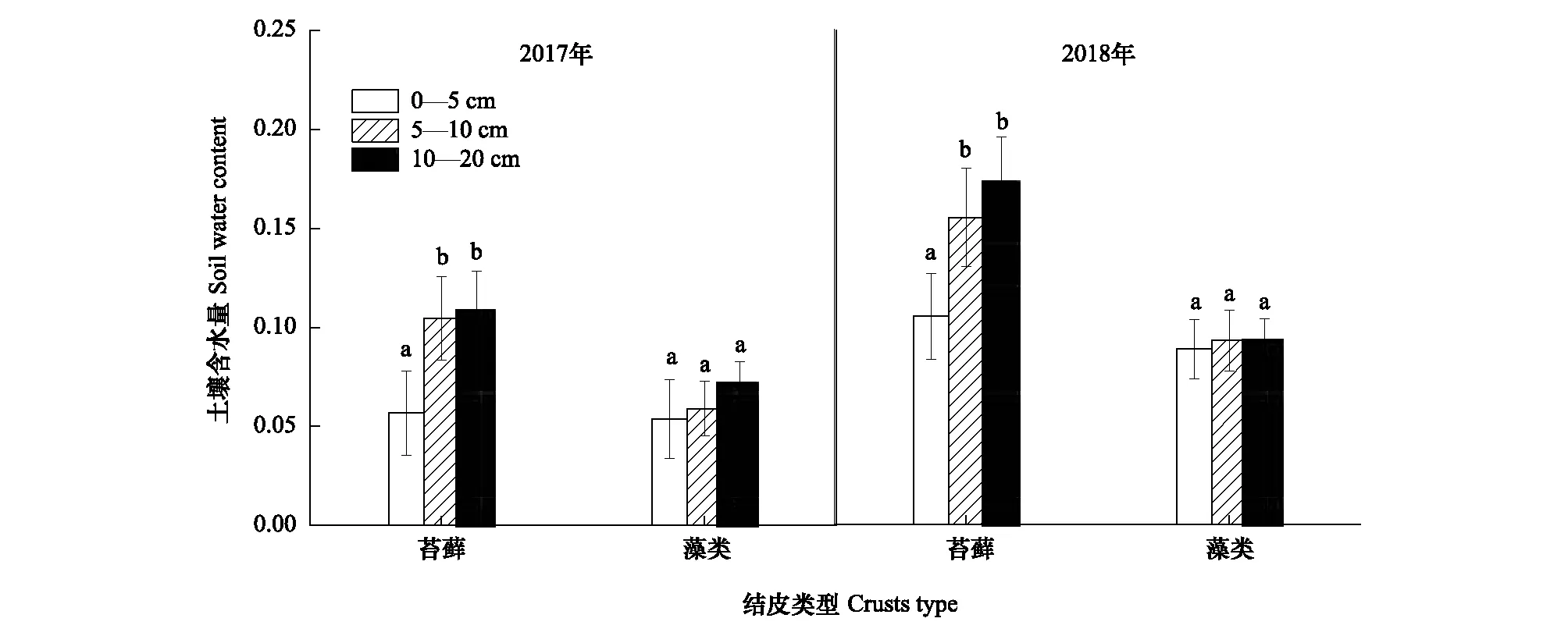
图5 生长季内(6—10月)不同类型生物土壤结皮下0—5 cm、5—10 cm和10—20 cm土壤体积含水量变化
4 结论
(1)裸地与结皮发育土壤碳释放速率日动态均呈“单峰”曲线,裸地与去除结皮土壤碳释放速率日最大值出现于15:00左右,两种类型生物土壤结皮覆盖土壤碳释放速率日最大值出现于13:00左右;日最大碳释放速率均出现于8月。
(2)裸地与结皮发育土壤碳释放速率的年内生长季动态规律一致,均呈“单峰”曲线,最大值出现在8月,最小值出现在10月。
(3)生物土壤结皮的存在增大了土壤的碳释放量,且结皮的发育程度越高,其增加量越大。藻类结皮层碳释放对土壤碳释放的促进作用更强,而苔藓结皮下层土壤碳释放对土壤碳释放的促进作用更强。土壤碳释放作用随降水 量的增加而增强。
(4)温度是影响土壤碳释放日动态的首要因素,其中土壤0—5 cm温度和含水量对各观测类型土壤碳释放速率的影响最大。
