头孢菌素C对蔬菜种子萌发的毒理效应
2020-11-11周睫雅任爱玲刘宏博吴昊马双崔思嘉王旭明田书磊
周睫雅,任爱玲,刘宏博,吴昊,马双,3,崔思嘉,4,王旭明,田书磊*
(1. 河北科技大学环境科学与工程学院,石家庄050080;2. 中国环境科学研究院固体废物污染控制技术研究所,北京100012;3. 内蒙古大学生态与环境学院,呼和浩特010021;4.东北电力大学化学工程学院,吉林 吉林132012;5.北京市农林科学院北京农业生物技术研究中心,北京100097)
我国是抗生素原料药生产和使用大国,每年生产的抗生素超过2×105t,国内消耗量约占75%,其中β-内酰胺类抗生素使用量较大,占总使用量的21%[1-4],头孢菌素C 是顶头孢霉菌产生的一种β-内酰胺类抗生素,是需求量较大的大宗类抗生素。据统计,2013年我国抗生素产量就已达到24.8 万t,依据生产1 t抗生素平均产生30~40 t 湿菌渣估算,2013 年我国抗生素菌渣产生量约为1 000 万t[4-6]。相关研究表明,在中国、美国和欧盟等多个国家的土壤[7]、地表水[8-9]、城市污水或废水[10]、沿海城市近岸海域[11]、蔬菜及肉食产品中[12-13],均能检出各类抗生素。课题组前期调研发现,不同企业或不同生产工艺产出的菌渣残留的抗生素量不同,螺旋霉素残留量为750~800 mg·kg-1[14],头孢菌素C 残留量为530~550 mg·L-1。抗生素菌渣因残留抗生素,其进入环境后将促进耐药菌的发生和传播[15],依据2016 年新版《国家危险废物名录》,抗生素菌渣被纳入危险废物名录。
按照危废管理要求,目前菌渣的安全处理处置方式主要为填埋和焚烧,这两种方式不仅安全处置难度大,而且费用高,已严重制约了抗生素制药行业的健康发展。因此,亟待开发安全、有效、经济合理的利用处置技术。根据课题组前期测样发现,经无害化处理后,头孢菌渣的含水率由90.68% 降至2.76%,而有机质、总养分(N + P2O5+ K2O)含量分别为89.00% 和10.57%,均满足《有机肥料》(NY 515—2012)要求。因此,肥料化是实现其资源化利用可行性较好的方式。抗生素菌渣因含有大量的粗蛋白、粗脂肪等有机物[16],经无害化处理技术消除残留抗生素后,可作为有机肥料基料,但缺少相关技术规范和标准。目前,中国环境科学研究院和抗生素菌渣国家工程技术中心已向国家标准化管理委员会提出制订青霉素、头孢菌素以及红霉素等典型大宗抗生素菌渣作为有机肥基料进行无害化处置技术要求、环境风险评估方法等相关技术规范的立项需求,为抗生素菌渣肥料化利用奠定基础。
菌渣中残留抗生素对植物生长毒性的影响是抗生素菌渣肥料化利用标准技术规范编制需解决问题之一。目前,国内外关于抗生素植物毒性的研究已有报道,主要通过种子发芽和幼苗生长发育实验来考察抗生素污染胁迫下植物种子发芽、生长等指标来综合评价其毒性。Hillis 等[15]对磺胺类和大环内脂类等抗生素对莴苣的生态毒性进行研究,发现金霉素、左氧氟沙星和磺胺甲恶唑生态毒性最为显著。Wang 等[16]采用水培法分别研究了土霉素和盐酸多西环素在0~90 mg·L-1、乳酸恩诺沙星在0~320 mg·L-1对油菜的生态毒性效应,发现在设定浓度范围内各种抗生素对根长的抑制作用均强于芽长。李萌等[17]通过白菜种子发芽实验,发现抗生素浓度与白菜根长抑制率之间存在剂量-效应关系。邓世杰等[18]研究发现,四环素对种子发芽率的最大无响应浓度(NOEC)为5 mg·L-1,环丙沙星和磺胺嘧啶对黑麦草种子发芽率的NOEC 为1 mg·L-1。课题组通过美国环境保护署(US EPA)毒性数据库和相关已发表的文献筛选收集了一部分β-内酰胺类(其中必须包括青霉素、头孢)等的水生和陆生生物(藻类、水生植物、鱼类、甲壳类和昆虫类等)的急慢性毒性效应数据,发现有关头孢菌素类药物的生态毒理学数据还不足以论证其对环境的具体影响。
本研究以生菜、油麦菜、白菜和油菜为研究对象,开展了不同浓度梯度下头孢菌素C 对4 种常见蔬菜种子的发芽、鲜质量、胚轴长和胚根长的毒性效应研究,对其根长进行了剂量-效应关系拟合,推导不同蔬菜种子对头孢菌素C的响应敏感程度,以期为头孢菌素类抗生素菌渣肥料化利用的环境基准生态阈值建立与生态风险评估提供科学依据。
1 材料与方法
1.1 实验材料
供试蔬菜种子:生菜(Lactuca sativaLinn. var. ramosa Hort.),油 麦 菜(Lactuca sativavar. longifoliaf.Lam),白菜(Brassica pekinensis(Lour.)Rupr.),油菜(Brassica campestrisL.),购于北京派得伟业科技发展有限公司。
供试头孢菌素C(cephalosporin C)标准品,由伊犁川宁生物技术有限公司提供(从菌渣中提炼纯化),纯度大于99%,分子式为C16H21N3O8S,结构式如图1。
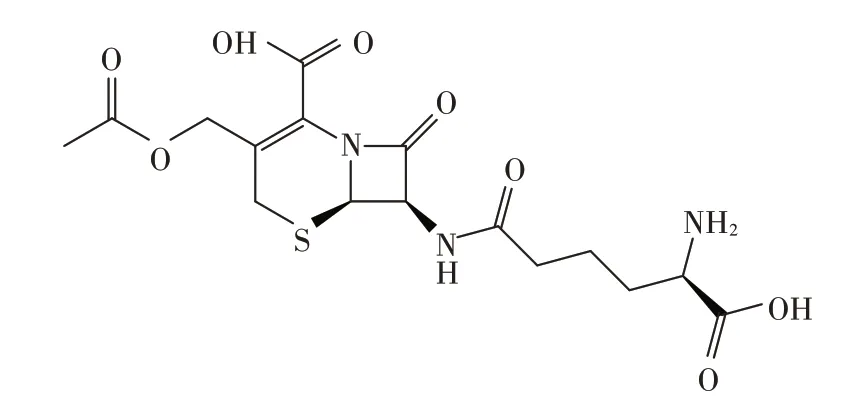
图1 头孢菌素C的结构式Figure 1 Constitutional formula of cephalosporin C
1.2 实验方法
参考国际种子检验协会(ISTA)的种子监测标准[19],采用保湿培养法进行试验。
用体积分数为10% 的H2O2溶液对种子消毒15 min,再用去离子水冲洗3 遍,滤干,于直径90 mm 无菌培养皿中放置1 张滤纸,将15 粒种子均匀排列在培养皿中,向培养皿中加入3 mL 不同浓度梯度的头孢菌素C 溶液,浓度梯度设定为:10、100、500、1 000、1 500、2 000 mg·L-1,以加入等量的去离子水为对照,试验设3 组平行。将培养皿置于(25±1)℃智能型光照培养箱(GXZ-280,宁波江南仪器厂)中黑暗培养,试验周期4 d。每日定时记录种子发芽数,通过恒质量法保持滤纸和种子湿润。试验期间,直到对照组的发芽率趋于稳定不变(发芽率≥85%);当对照组胚根长>20 mm 时,结束培养并测定幼苗鲜质量,胚根长和轴长。
1.3 数据处理
1.3.1 发芽率、胚根长生长率的计算
发芽率和根生长率的计算公式如下:
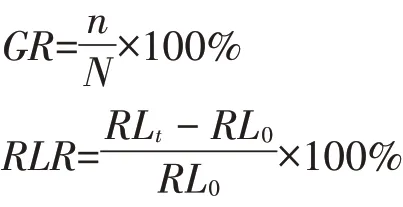
式中:GR为发芽率,%;n为发芽种子数,个;N为供试种子数,个;RLR为胚根生长率,%;RL0为对照组胚根长,mm;RLt为处理组胚根长,mm。
1.3.2 最大无响应浓度(NOEC)及半抑制浓度(IC50)的计算
建立以头孢菌素C浓度与根生长率的剂量-效应关系图,并进行拟合。当根生长率为0 时,对应的头孢菌素C 浓度即为该种抗生素的NOEC 值;当根生长率为-50% 时,对应的头孢菌素C 浓度即为该种抗生素的IC50值。
1.3.3 统计分析
采用IBM SPSS Statistics 25 统计分析软件对数据进行单因素(One-way ANOVA)方差分析和差异显著性检验,P=0.05 为显著性指标,用Origin 2019 对实验数据进行作图。图表中各指标的数据为3 组平行实验的平均值,或平均值±标准差。
2 结果与分析
2.1 不同浓度头孢菌素C胁迫下对种子发芽率的影响
如表1 所示,头孢菌素C 处理下生菜、油麦菜、白菜及油菜种子的发芽率与对照组相比无显著差异(P>0.05),即在本试验浓度范围内,头孢菌素C 对供试的4 种蔬菜种子的萌发无明显抑制或促进作用。这与Derek 等[20]进行的10 种抗生素对苦菊、莴苣、紫花苜及胡萝卜的植物毒性试验的结果相似。其主要原因,一是种子发芽期较短,抗生素还未对种子产生明显影响就已经萌发,在种子萌发初期,植物幼嫩细胞中未形成液泡,根也还未破皮而出,此时种子对水分的吸收是沿着水势梯度进行的吸胀吸水物理过程;二是种皮的保护作用,抗生素吸附在种皮表面,阻碍了胚根与外界环境直接接触,使得种子萌发过程受到的侵害较不明显[21]。种子萌发是种子从吸水到胚根突破种皮期间一系列酶催化的生理生化过程。种子内各类参与萌发的酶在此阶段激活,并作用于种子子叶储存的淀粉、蛋白质和脂肪的分解活动,为后期幼苗植物细胞的合成提供能量来源。这就意味着,从表观上看种子正常发芽,但并不能断定抗生素的胁迫对植物细胞无毒性效应[22]。
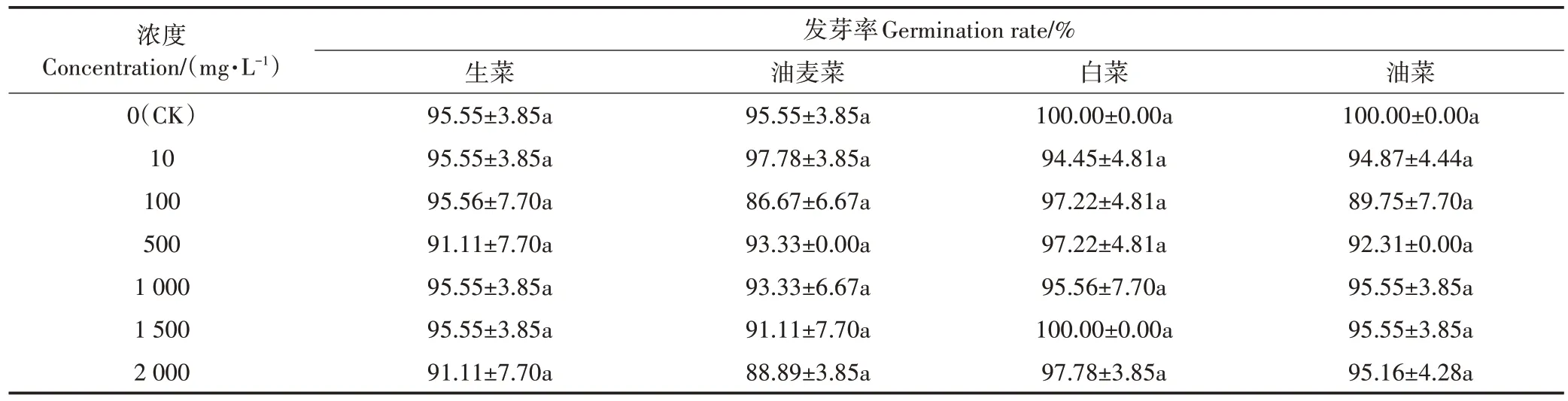
表1 不同浓度头孢菌素C处理下蔬菜种子的发芽率Table 1 Germination rate of seeds under cephalosporin C stress
2.2 不同浓度头孢菌素C 胁迫下对幼苗胚轴和鲜质量的影响
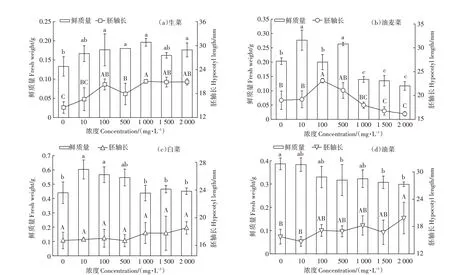
图2 不同浓度头孢菌素C溶液对4种蔬菜幼苗胚轴和鲜质量的影响Figure 2 Effects of cephalosporin C on fresh weight and hypocotyl of different seedlings
由图2 可见,处理组生菜的胚轴长均高于对照组,而油麦菜胚轴长则随着头孢菌素C浓度的增加呈现先增长后下降的趋势。油菜的胚轴长与头孢菌素C 浓度≤1 500 mg·L-1时,与对照差别不大,当浓度为2 000 mg·L-1时,则略高于对照。而白菜的胚轴长在不同浓度之间差异不显著。相对于胚轴长,头孢菌素C 对4 种蔬菜幼苗鲜质量的影响更为明显,但也存在一定的差异。不同浓度头孢菌素C溶液胁迫下,生菜及白菜的鲜质量均高于对照组,油菜则低于对照组,油麦菜鲜质量在头孢菌素C 浓度≥1 000 mg·L-1时,也低于对照组。随着头孢菌素C 的浓度提高,4 种蔬菜幼苗的鲜质量均呈现不同程度下降趋势,尤其是白菜和油麦菜,而生菜和油菜则不太显著。当浓度为10 mg·L-1时,白菜鲜质量最高,比对照组高37.11%;当浓度为1 000 mg·L-1时,生菜鲜质量最高,比对照组高47.04%。当头孢菌素C 浓度为1 000~2 000 mg·L-1时,油麦菜鲜质量则显著低于对照。
与发芽率和幼苗胚轴相比较,生菜、油麦菜、白菜及油菜幼苗鲜质量对头孢菌素C 的胁迫敏感性更显著,其原因是在胚根向胚轴传递运输过程中,头孢菌素C的浓度沿水势方向递减,使得在幼苗生长发育过程中,胚轴与头孢菌素C 溶液接触的浓度较低,受到的影响小。
2.3 不同浓度头孢菌素C胁迫下对幼苗胚根的影响
胚根是种子萌发后首先出现的器官,具有固着、贮存合成氨基酸和植物激素等有机物质的作用,其主要功能是吸收环境中水分及养分,并通过维管组织输送给茎。幼苗生长过程根尖与抗生素溶液接触非常紧密,对其中的抗生素响应较为显著。Alessandro等[23]研究了4 种抗生素(氯霉素、螺旋霉素、大观霉素及万古霉素)对番茄种子萌发过程中根系发育的影响,证明了4 种抗生素对番茄顶端根系细胞分裂造成明显损害。
由图3 可知,随着溶液中头孢菌素C 浓度的提高,4 种蔬菜胚根长整体呈现出先促进后抑制的现象,当浓度为100 mg·L-1时,基本均达到生长峰值。随着头孢菌素C浓度逐渐增大,其对胚根长的抑制作用越趋明显。4种蔬菜对头孢菌素C的响应程度各不同,但趋势相似。与发芽率、幼苗鲜质量及胚轴长相比,胚根对头孢菌素C 胁迫的敏感性最为显著,选取胚根长作为4 种蔬菜受损程度的判断指标。葛成军等[24]在进行四环素类抗生素对白菜种子发芽实验时也得出了类似结论,试供作物对金霉素和土霉素的生态敏感性表现为:根伸长>轴伸长>发芽率。张乙涵等[25]也报道根长可作为诊断抗生素植物毒性的敏感指标。
3 讨论
3.1 不同毒性指标与头孢菌素C之间的关系
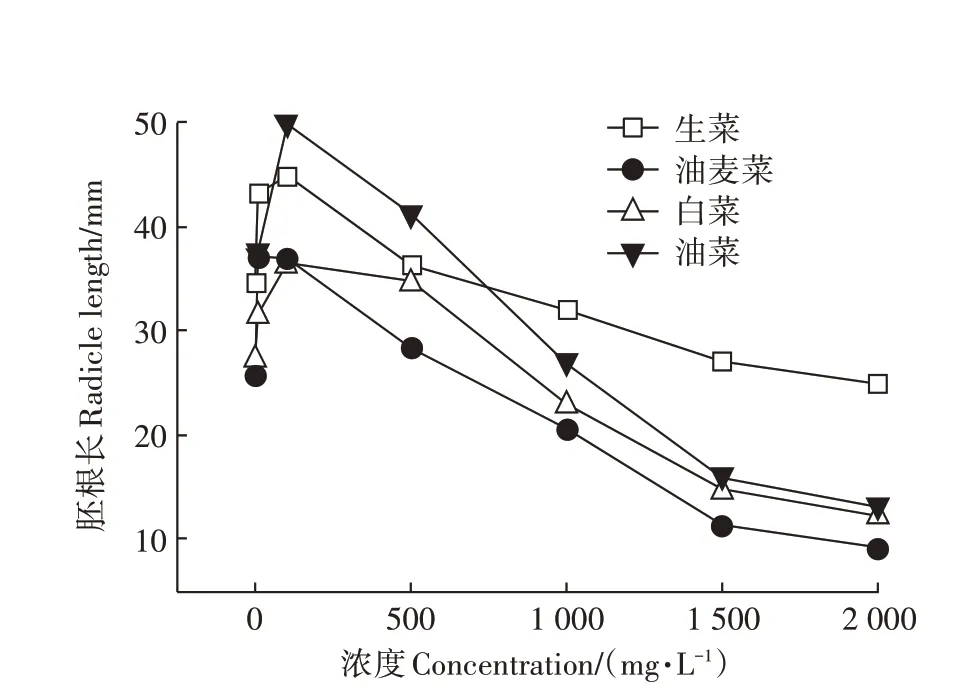
图3 不同浓度头孢菌素C溶液对胚根影响Figure 3 Effects of cephalosporin C on radicle length under stress
对发芽率、幼苗鲜质量、胚轴长及胚根和头孢菌素C 浓度及蔬菜种类进行了主成分分析(PCA),见图4。方差贡献率第一主成分为38.0%,第二主成分为29.6%,累计方差贡献率为67.6%,共同解释所有样品的差异。第一主成分的特征指标有发芽率、鲜质量和胚根,其中头孢菌素C 浓度对胚根长呈负相关,说明头孢浓度变化会影响胚根的生长状态。根是蔬菜主要的吸收和代谢器官,同时也是最容易受毒物分子影响的部位,且与头孢菌素C 直接接触,通过影响蔬菜根部的抗氧化防御系统和细胞分裂来影响根长[26-27],也有研究指出,种子萌发发生在植物暗生芽期,此时胚根是快速生长的器官,对抗生素敏感[27],使胚根长在头孢菌素C 存在下受到显著抑制(P<0.05)。第二主成分的特征指标是发芽率和鲜质量,种子发芽率与鲜质量呈正相关,种子的发芽数,直接对幼苗鲜质量产生影响。
由图4 可见,生菜组与油麦菜组之间的样品相似性高,且有部分重叠,油菜和白菜之间也有相同的规律。本试验的供试蔬菜中,油麦菜和生菜均属于菊科莴苣属,油菜和白菜均属于十字花科芸薹属,造成不同属蔬菜在头孢菌素C 溶液胁迫下呈现出不同的聚集状态,这可能与蔬菜中营养物质的储存状态有关[28]。与莴苣属蔬菜相比,芸薹属蔬菜种子中不具有淀粉粒,使得抗生素作用在不同属蔬菜中的靶分子不同[29],但其具体机制还需进一步研究。
不同浓度头孢菌素C处理下,发芽率、鲜质量、胚轴及胚根毒性指标都会受到不同程度的影响。图4的相关性分析可见头孢菌素C 浓度对胚根的影响最为显著,其次是鲜质量和胚轴,影响最不显著的是发芽率。抗生素对胚轴的作用机制主要是种子胚根与抗生素接触后,将吸收抗生素并沿着胚轴向上输送,所以与其他部位相比,根中抗生素累积量最多[30],多位学者的研究结果[31-33]指出,随抗生素浓度的升高,蔬菜胚轴长和胚根长生长逐渐受到抑制,且受抑制程度胚根长>胚轴长。
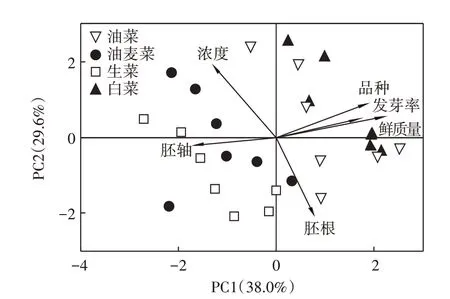
图4 不同毒性指标与头孢菌素C浓度之间的关系Figure 4 The principal components analysis between different toxicity indexes and cephalosporin C
3.2 基于胚根长的不同蔬菜敏感程度分析
试验胚根长为重要毒性指标,以头孢菌素C 浓度为自变量,根生长率为因变量,建立了的根生长率与头孢菌素C溶液浓度响应曲线(图5a),获得回归方程(表2)。从表2可知,4种蔬菜根的生长率与头孢菌素C 浓度间均呈现出极显著的线性关系(P<0.01)。由图5b 及表2 可知,生菜、油麦菜、白菜和油菜的NOEC分别为868.10、727.25、796.27 mg·L-1和629.27 mg·L-1。当头孢菌素C 浓度小于不同蔬菜各自对应的NOEC 时,头孢菌素C 对4 种蔬菜胚根的伸长具有明显的促进作用,当浓度超出了幼苗耐受范围,则出现了抑制效应,但根伸长抑制率存在差异。刘娣[34]进行的小白菜水培实验,同样发现低浓度四环素类抗生素促进了小白菜种子根的伸长,而高浓度的四环素类抗生素则抑制了根伸长。蔡嘉颖等[35]也发现,土霉素在低浓度(≤5 mg·L-1)对苦菊根伸长有促进作用,高浓度(≥10 mg·L-1)时抑制。这说明抗生素类物质对试验幼苗生长具有双向调节功能,其主要原因可能与植物的逆境生理相关[36],当抗生素浓度在幼苗体内超出其生理承受范围,胚根就受到影响,同时高浓度抗生素会阻止抗氧化酶系统的信号传递[37],使植物正常代谢受到影响。
目前,常用植物的IC50值来评价污染物的生态毒性强弱[38]。IC50值与植物对头孢菌素C 的敏感程度负相关。从图5b 可见,头孢菌素C 对4 种蔬菜的敏感度依次为:油菜>油麦菜>白菜>生菜。Pan 等[39]通过小油菜、番茄、胡萝卜和黄瓜的根伸长试验评价四环素(TC)、磺胺甲嘧啶(SMZ)、诺氟沙星(NOR)、红霉素(ERY)和氯霉素(CAP)的毒性,得到上述抗生素对小油菜的IC50值分别为14.4、157、49.4、68.8 mg·L-1和204 mg·L-1,均小于本研究头孢菌素C 对油菜的IC50值,即1 538.36 mg·L-1。头孢菌素C 对小油菜的毒性远小于上述抗生素,这主要是由于抗生素毒性与其分子结构及其稳定性具有相关性,头孢菌素类分子具有不稳定、高应变和反应性的β-内酰胺键,易于水解,半衰期较短,因此对胚根毒性影响相对较小。由此可见,针对不同种类的蔬菜对头孢菌素C 的耐受程度各不相同,头孢菌渣的肥料化利用过程,应充分考虑菌渣中头孢菌素C 的残留情况,合理种植蔬菜,保证农产品安全,减少头孢菌素C 沿食物链进入人体的风险。
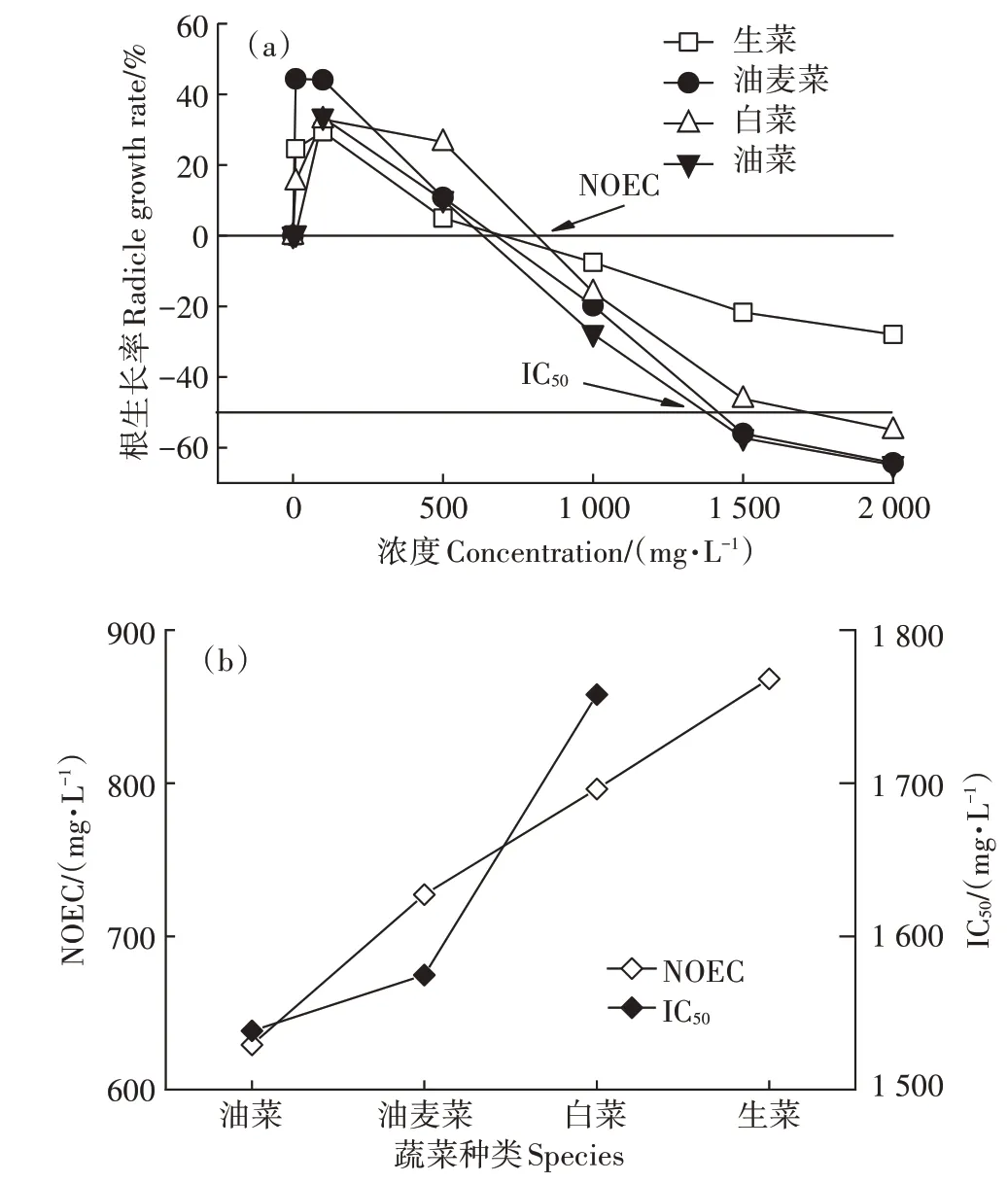
图5 浓度-根生长率的剂量-效应曲线Figure 5 Dose-response curve of concentration-radicle growth rate
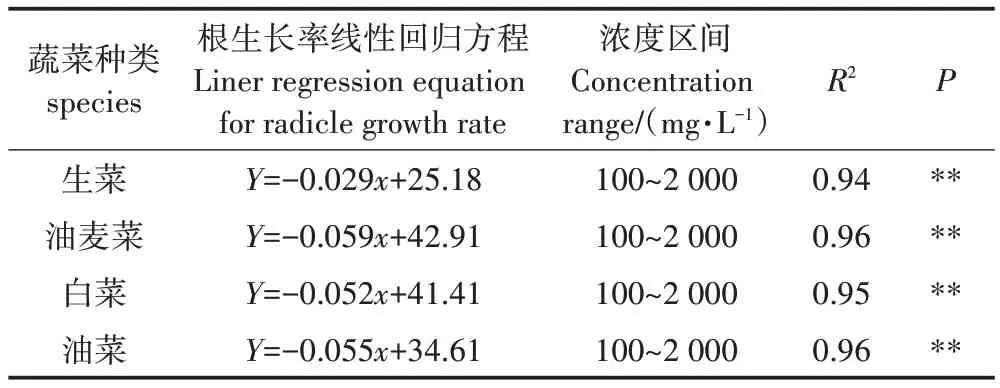
表2 头孢菌素C与根生长率的相关性Table 2 Correlation between cephalosporin C and radicle growth rate
4 结论
(1)4 种蔬菜种子的毒性指标对头孢菌素C 的生态敏感性均表现为:胚根>鲜质量>胚轴>发芽率。胚根长作为诊断头孢菌素C生态毒性的敏感指标,可较好反映头孢菌素C的污染状况。
(2)生菜、油麦菜、白菜及油菜胚根的NOEC 分别为868.10、727.25、796.27 mg · L-1和629.27 mg · L-1。NOEC 可为头孢菌渣残留量的环境安全阈值的设定提供基础数据。
(3)4种蔬菜对头孢菌素C的敏感分布顺序为:油菜>油麦菜>白菜>生菜。结果表明,油菜在所测试的蔬菜中是最为敏感的,其IC50为1 538.36 mg·L-1。油菜将作为后续探究施用头孢菌渣肥对土壤抗性基因及微生物群落影响较为理想的毒性指示作物。
