LncRNA HULC通过miR-372/CXCR4轴来调控肝癌细胞的增殖、凋亡与上皮-间质转化
2020-11-09刘远光刘怀海
刘远光刘怀海
(1.河北省唐山市工人医院肝胆外科,河北唐山 063000; 2.河北医科大学第二医院普外科,石家庄 050000)
肝细胞癌(hepatocellular carcinoma,HCC)是高风险、高度有害的恶性肿瘤,对人类健康构成威胁[1]。此外,肝癌的发病机理尚未完全了解。据报道,环境和饮食因素影响肝癌的发生。目前,手术仍是肝癌治疗的首选。但是,肝癌的复发率仍然很高,其5 年生存率一般低于50%[2]。因此,探索肝癌的发病机制以筛选有效的诊断标志物和治疗靶点对于肝癌患者非常迫切。
长非编码RNA(LncRNA)是一类RNA 转录物,长度大于200 个核苷酸[3]。LncRNA 可以调节蛋白质编码基因、转录和转录后,并在生物学过程中发挥重要作用[4]。研究表明,已LncRNA 在生物学过程中的许多功能,包括心血管疾病、炎症反应和癌症[5]。这些作用由表观遗传调控、转录和转录后调控介导。在 HCC 中已鉴定出许多 LncRNA,例如H19、HOTAIR、MALAT1 和 MEG3[6]。肝癌中高度上调(highly upregulated in liver cancer,HULC)是一种长的非编码RNA(lncRNA),最近已被确定为肝细胞癌,神经胶质瘤和胃癌进展中的关键调节剂[7]。研究表明,HULC 在肝癌发生过程中发挥积极作用,例如,HULC 调节肝癌细胞中的脂质代谢[8]。因此,LncRNA 可以作为肝癌诊断、预后和药物基因组学的潜在标志物。
microRNA(miRNA)是小的非编码RNA,其长度约为22 个核苷酸,实际上参与了细胞所有过程,例如增殖、分裂分化、凋亡、蛋白质分泌和病毒感染过程[9]。通常,miRNA 通过抑制翻译或诱导目标信使RNA(mRNA) 降解来负调控基因表达。但是,miRNA 也可以通过调节启动子活性或激活翻译来积极调节基因表达[10]。近几年的研究表明miRNA在肝病发展和进展中的特定作用[11],提示miRNA可能与肝癌的发生发展有关。因此,miRNA 可以作为肝癌诊断、预后和药物基因组学的潜在标志物。
C-X-C 趋化因子受体 4 (C-X-C chemokine receptor 4,CXCR4)是趋化因子配体12(CXCL12)特异受体。大量研究表明,CXCR4 与HCC 的发展有关,包括细胞增殖、血管生成、侵袭和转移[12-13]。例如,HCC 细胞中EphA1 表达的升高增强了内皮祖细胞(EPC)向肿瘤部位的募集,从而通过激活SDF-1/CXCR4 轴来促进血管生成[14]。肿瘤内皮细胞中高水平的CXCR4 可以诱导肿瘤血管生成,从而促进肿瘤转移并与HCC 患者的不良预后相关[15]。因此,CXCR4 在肝癌的发生发展中起着重要作用。
综上所述,本研究分析了 LncRNA、miRNA 和mRNA 在HCC 组织和肝癌细胞系中的表达,研究分析了 HULC、miR-372 和 CXCR4 的潜在关系以及HULC/miR-372/CXCR4 轴对肝癌细胞增殖、凋亡与上皮-间质转化的影响。我们的研究结果可能有助于增进对肝癌细胞分子机制的了解,LncRNA HULC可能作为HCC 的潜在诊断生物标志物和治疗靶标。
1 材料和方法
1.1 实验材料
1.1.1 临床组织
所有肿瘤组织(20 例)和配对的癌旁正常组织(20 例)均来自接受肝癌切除手术的患者。所有患者均未接受任何栓塞治疗或在手术前进行化学疗法,并在术后进行病理诊断。所有患者均签署了知情同意书,所有实验均获得本医院伦理委员会的批准与监督(IACUC2018011505)。每个组织标本都保存在-80℃直至使用。
1.1.2 实验细胞
六种 HCC 细胞系 Huh7 (批号:SAc0188)、Hep3B(批号:I110643)、MHCC-97L(批号:YCL-0497)、MHCC-97H(批号:CL-H153)、SMMC-7721(批号:L4821)、HepG2(批号:YS-C3688)和正常肝细胞系(LO2,批号:0158),均购自中国科学院上海细胞生物学研究所。
1.2 主要试剂与仪器
RPMI-1640 培养基 (批号: CD-100043GM)、DMEM 培养基(批号:BH-S3208)和胎牛血清(批号:164210-500)(美国 Gibco 公司);CCK-8 试剂盒(批号:IC-CCK8-Hu)、青霉素(批号:B25911)和链霉素(批号:S66602)(美国 Sigma 公司);TRIzol 试剂(批号:CD102523GM)、cDNA 逆转录试剂盒(批号:4368814)和 SYBR Premix EX TaqTM试剂盒(批号:RR420A)(日本 TaKaRa 公司);阴性对照、miR-372 mimics、miR-372 inhibitor ( Genepharma 公 司 );PcDNA3.1-control、pcDNA3.1-HULC、si-control、si-HULC 和 LipofectamineTM3000(批号:L3000015)(美国Invitrogen 公司);pmirGLO 载体(批号 GN1439)、双荧光素酶活性检测试剂盒(批号:C-8304)(美国Promega 公司);Annexin V-FITC/PI 试剂盒(批号:120248)、RIPA 裂解液(批号:R21235)(英国 Abcam公司);DAB 显色剂(批号:SNM477-ARB,北京百奥莱博科技有限公司);双氧水(批号:R23672,扬州市天泰化工有限公司); E-cadherin 抗体(批号:3195T)、Vimentin 抗体(批号:5741T)、β-actin 抗体(批号:4970T)和山羊抗兔IgG 二抗(批号:2985S)(美国 Cell Signaling Technology 公司)。SDS-PAGE电泳仪(型号:DYY-7C,北京六一生物科技有限公司);凝胶扫描仪(型号:4466611,美国BIO-RAD 公司);酶标仪(型号:SMR16.1,上海科华实验系统有限公司);流式细胞仪(型号:1026,美国BD 公司);StepOnePlus Real-Time PCR System(型号:401411,日本 TaKaRa 公司);倒置相差显微镜(型号:CKX53,日本 Olympus 公司)。
1.3 实验方法
1.3.1 细胞培养
LO2 细胞用含10%胎牛血清和100 IU/mL 青霉素、100 IU/mL 链霉素的 RPMI-1640 培养基培养;Huh7、Hep3B、MHCC-97L、MHCC-97H,SMMC-7721和HepG2 细胞用含10%胎牛血清和100 IU/mL 青霉素、100 IU/mL 链霉素的DMEM 培养基培养,置于37℃、5% CO2的培养箱,2 d 更换一次培养基,以保证细胞所需的营养成分。取对数期、状态良好的细胞进行后续实验。
1.3.2 Quantitative real time RT-PCR(qRT-PCR)
用TRIzol 试剂提取组织样本或者细胞总RNA。用Nanodrop 2000 仪器测量样品总RNA 浓度,并将每个配对的样品调节至相同浓度。然后按照cDNA逆转录试剂盒将总 RNA 反转录成 cDNA。使用SYBR Premix EX TaqTM试剂盒进行 qRT-PCR。U6和 β-actin 分别用作 LncRNA、miRNA 和 mRNA 的内部参考。使用FTC-3000p 实时PCR 系统完成实验后,采用 2-△△Ct法分析数据。其中使用的引物见表1。
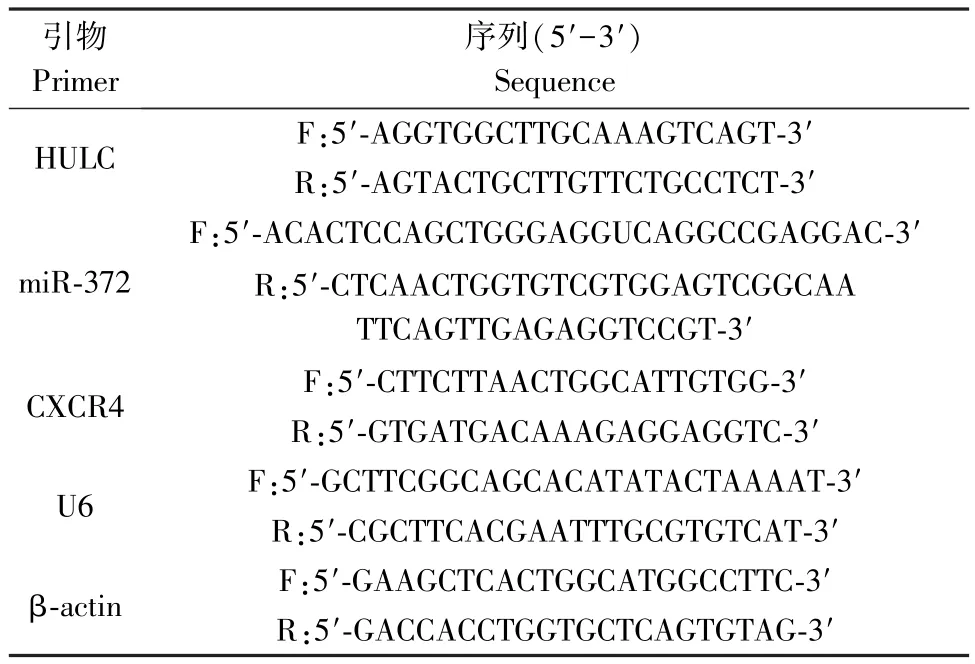
表1 引物信息Table 1 Primer information
1.3.3 细胞转染
按照LipofectamineTM3000 转染试剂说明书将质粒(阴性对照、miR-372 mimics、miR-372 inhibitor、pcDNA3.1-control、pcDNA3.1-HULC、si-control 和 si-HULC)分别转入至 MHCC-97L 和 HepG2 细胞中。在37℃、5% CO2的培养箱中培养6 h 后,更换新鲜的培养基继续培养。48 h 后结束培养,收集各组细胞进行后续实验。
1.3.4 双荧光素酶报告基因检测
通过 TargetScan 数据库分析 miR-372 分别与HULC 和CXCR4 的非编码区区域的结合位点。将HULC 3′UTR wt/mut 或 CXCR4 3′UTR wt/mut 克隆到pmirGLO 载体中,然后将阴性对照、miR-372 mimics、miR-372inhibitor、pmirGLO-HULC-wt、pmirGLO-HULC-mut、pmirGLO-CXCR4-wt 或pmirGLO-CXCR4-mut 转染至 MHCC-97L 或 HepG2细胞中。48 h 后,用双荧光素酶活性检测试剂盒测定MHCC-97L 细胞和HepG2 细胞的荧光素酶活性。
1.3.5 CCK-8
将转染后的细胞接种到96 孔板上,培养24、48、72、96 h。然后,将 10 μL CCK-8 溶液添加到每个孔中,并在37°C 下孵育2 h。最后,使用自动酶标仪检测在450 nm 波长的吸光度。
1.3.6 细胞集落形成实验
将MHCC-97L、HepG2 细胞分别接种到6 孔板中,并用指定的质粒转染。48 h 后,收集各组细胞并铺在60 mm 的培养皿中。3 d 更换一次培养基。14 d 后,用结晶紫染色,并计数含有≥50 个细胞的集落,并进行拍照。
1.3.7 细胞流式术
收集转染的各组细胞,离心并重悬于结合缓冲液中。将细胞依次用异硫氰酸荧光素(Annexin VFITC)溶液(10 μL)和碘化丙啶(PI)溶液(10 μL)染色。将染色的细胞在室温下黑暗中孵育30 min,然后使用FACSCaliburTM流式细胞仪进行分析。
1.3.8 免疫组织化学染色
组织经固定、包埋、石蜡切片、脱水、缓冲液清洗,3%过氧化氢溶液去除内源性过氧化物酶,滴加非免疫动物血清20 min,滴加一抗、PBS 冲洗、滴加二抗、冲洗后加 ABC 复合物、DAB 显色,在400 倍显微镜下观察并拍照。
1.3.9 Western blot
各组细胞用RIPA 裂解提取总蛋白,并通过BCA 方法进行定量,样品经变性后进行上样。按照实验步骤进行电泳-转膜-封闭(5%脱脂奶粉)-一抗孵育(4℃过夜)-二抗孵育(室温1 h)-显影曝光。Image-Pro Plus 图像分析系统对蛋白条带进行分析。
1.4 统计学方法
统计学分析用SPSS 19.0 软件进行,至少取3次独立实验结果,以平均数±标准差(±s)表示,多组间数据比较采用单向方差分析,组间两两比较采用LSD-t检验P<0.05 表示差异有统计学意义。
2 结果
2.1 HULC 在肝癌组织和肝癌细胞中高表达
qRT-PCR 结果表明HULC 在肝癌组织和肝癌细胞中高表达:与癌旁非肿瘤组织组相比,HULC 在肝癌组织中表达上调(P<0.01,图1A);与LO2 细胞相比,在六个HCC 细胞系中均观察到HULC 高表达,其中,MHCC-97L 细胞中 HULC 表达最低(P<0.05),而 HepG2 细胞中 HULC 表达最高(P<0.001,图1B),因此,因此这两种细胞系分别用于功能增强和功能缺失实验。总之,研究人员得到结论:HULC 在肝癌组织和肝癌细胞中高表达。

图1 HULC 在肝癌组织和肝癌细胞中的表达Figure 1 HULC expression in liver cancer tissues and liver cancer cells
2.2 HULC 对HCC 细胞增殖和凋亡的影响
pcDNA3.1-HULC 转染 MHCC-97L 细胞,si-HULC 转染 HepG2 细胞。qRT-PCR 分析显示,pcDNA3.1-HULC 和 si-HULC 在 MHCC-97L 和HepG2 细胞中显著调节了 HULC 的表达(均P<0.01,图2A、2B)。从功能上看,CCK-8 分析表明,过表达HULC 明显增强MHCC-97L 细胞的活力(P<0.01;图2C),而低表达HULC 明显减弱HepG2 细胞的活力(P<0.01,图2D)。细胞集落形成实验表明,过表达HULC 明显增强MHCC-97L 细胞的增殖能力(P<0.01,图2E),而低表达HULC 明显减弱HepG2细胞的增值能力(P<0.01,图2F)。细胞流式术结果表明,过表达HULC 明显抑制MHCC-97L 细胞的凋亡能力(P<0.01,图2G),而低表达 HULC 明显增强HepG2 细胞的凋亡能力(P<0.01,图2H)。
2.3 HULC 对 HCC 细胞 EMT 的影响
免疫组织化学染色揭示了HULC 和EMT 标记物(E-cadherin 和Vimentin)之间的关联,结果表明,与低表达HULC 组相比,过表达HULC 组的HCC 组织具有较低的E-cadherin 和较高的Vimentin 表达(P<0.01,图3A)。Western blot 结果表明,过表达HULC 可以明显抑制MHCC-97H 细胞中E-cadherin的表达并提高Vimentin 的表达(均P<0.01);低表达HULC 可提高 HepG2 细胞中 E-cadherin 的表达并抑制 Vimentin 的表达(均P<0.01,图3B)。综上所述,HULC 促进HCC 细胞的增殖、抑制HCC 细胞凋亡以及促进HCC 细胞EMT 的进展。
2.4 miR-372 是 HULC 的靶标
TargetScan 数据库揭示了 miR-372 与 HULC 的潜在结合位点(图4A)。qRT-PCR 的数据表明,miR-372 在肝癌组织和肝癌细胞中的表达明显下降(均P<0.01,图4B、4C)。Pearson 相关分析显示,miR-372 与 HCC 组织中的 HULC 表达相关分析表明,在肝癌组织中观察到miR-372 与HULC 表达之间呈负相关(r=-0.56,P<0.001,图4D)。接下来,分别将 MHCC-97L 和 Hep2G 细胞转染了miR-372 抑制剂和 miR-372 模拟物,qRT-PCR 证实miR-372 的低表达和过表达(均P<0.05,图4E)。双荧光素酶报告基因测定表明,miR-372 明显抑制了携带HULC 3′UTR wt 的荧光素酶活性,但当报告质粒携带HULC 3′UTR mut 时未观察到明显的抑制作用(均P<0.01,图4F)。此外,HULC 可以负调控HCC 细胞中miR-372 的表达(均P<0.05,图4G)。另一方面,通过调节HCC 细胞中的miR-372 水平,HULC 表达明显升高或降低(均P<0.01,图4H)。综上所述,HULC 可直接与 HCC 细胞中的 miR-372 结合,且两者具有相互抑制的作用。

图2 HULC 促进HCC 细胞的增殖、抑制HCC 细胞凋亡Figure 2 HULC promotes HCC cell proliferation and inhibits HCC cell apoptosis
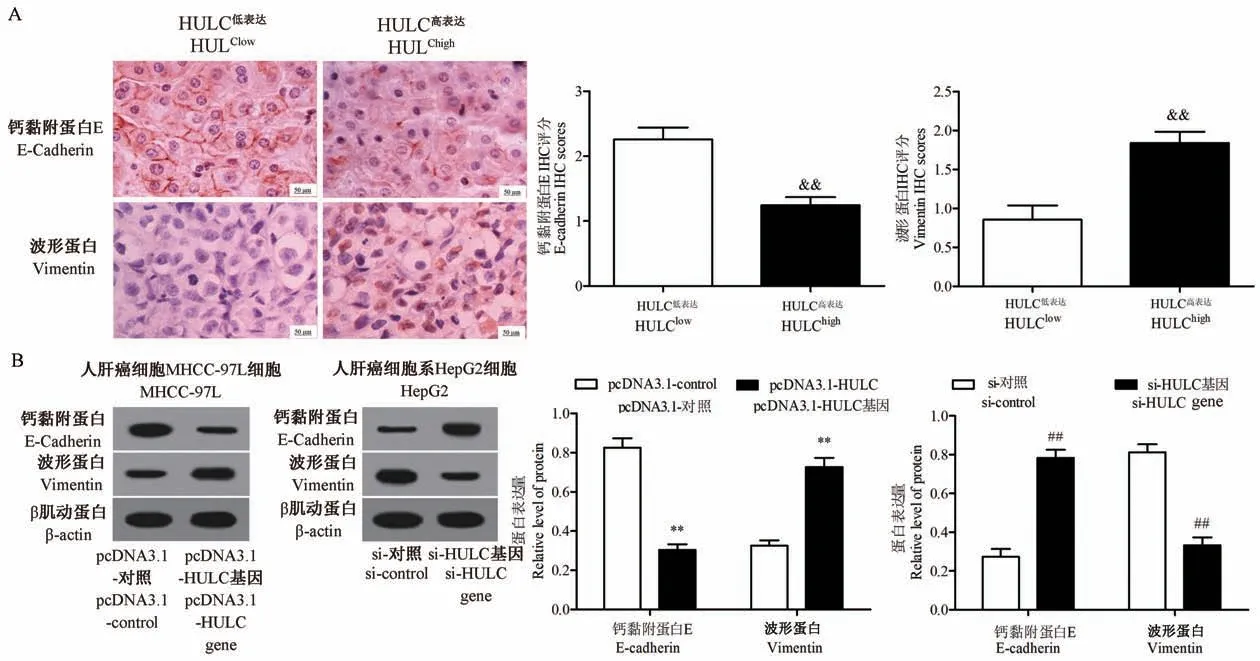
图3 HULC 促进 HCC 细胞 EMT 进展Figure 3 HULC promotes the EMT progression of HCC cells
2.5 miR-372 对HCC 细胞的增殖和凋亡的影响
CCK-8 分析表明,低表达miR-372 明显增强了MHCC-97 的活力(P<0.01),而过表达 miR-372 明显降低HepG2 细胞的活力(P<0.01,图5A)。细胞集落形成实验表明,低表达miR-372 明显增强了MHCC-97 的增殖能力(P<0.01),而过表达miR-372明显降低HepG2 细胞的增殖能力(P<0.01,图5B)。细胞流式结果表明,敲低 miR-372 明显抑制了MHCC-97 的凋亡能力(P<0.05),而过表达miR-372明显增强HepG2 细胞的凋亡能力(P<0.01,图5C)。
2.6 miR-372 对 HCC 细胞 EMT 的影响
免疫组织化学染色以探讨miR-372 和EMT 标记物(E-cadherin 和Vimentin)之间的关联,结果表明,与高 miR-372 组相比,低 miR-372 组的 E-cadherin 表达明显降低,且Vimentin 表达明显升高(均P<0.01,图6A)。此外,Western blot 分析显示,低表达 miR-372 可降低 MHCC-97L 细胞中 E-cadherin 的表达并增加 Vimentin 的表达含量(P<0.01),而过表达 miR-372 抑制了 HepG2 细胞的EMT 过程(均P<0.01,图6B)。因此,我们得出的结论是miR-372 可以抑制HCC 细胞的增殖、促进凋亡以及抑制HCC 细胞EMT 的进展。
2.7 miR-372 直接靶向HCC 细胞中的CXCR4
TargetScan 预测了 miR-372 与 CXCR4 的结合位点(图7A)。qRT-PCR 的数据表明,CXCR4 在肝癌组织和肝癌细胞中的表达明显上调降(均P<0.05,图7B、7C)。Pearson 相关分析显示,miR-372 与HCC 组织中CXCR4 表达相关分析表明,在肝癌组织中观察到miR-372 与CXCR4 表达之间呈负相关(r=-0.415,P<0.01,图7D)。使用双荧光素酶报告基因检测CXCR4 的表达是否受miR-372 调节,结果表明,当报告质粒携带CXCR4 3′UTR wt 时,miR-372 抑制了HCC 细胞中的荧光素酶活性,但当报告质粒携带CXCR4 3′UTR mut 时未观察到明显的抑制作用(均P<0.01;图7E)。qRT-PCR 分析结果表明,miR-372 可以直接靶向 CXCR4,并负调节CXCR4 在 HCC 细胞中的表达(均P<0.01,图7F)。接下来,我们探讨了HULC 是否可以调节CXCR4 的表达,数据表明,CXCR4 表达可以在肝癌细胞中受到 HULC 的正调控(均P<0.01,图7G)。因此,我们得出的结论是,CXCR4 是miR-372 的直接靶标,并且在HCC 细胞中受到HULC 的正调控。

图4 miR-372 在肝癌组织和肝癌细胞中的表达以及与HULC 的关系Figure 4 miR-372 expression in liver cancer tissues and liver cancer cells and its relationship with HULC
2.8 HULC/miR-372/CXCR4 轴可影响肝癌细胞的增殖、凋亡与EMT
qRT-PCR 结果表明,在 MHCC-97L 细胞中,过表达HULC 使CXCR4 表达上调,而过表达miR-372下调了 CXCR4 的表达(P<0.01);在 HepG2 细胞中,低表达HULC 使CXCR4 的表达下调,而低表达miR-372 上调了 CXCR4 的表达(P<0.01,图8A)。CCK-8 分析表明,过表达 HULC 使MHCC-97L 细胞活力增强,而过表达miR-372 降低了细胞活力(P<0.01);低表达HULC 降低了HepG2 细胞活力,而低表达 miR-372 增加细胞活力(P<0.01,图8B)。细胞集落形成实验表明,过表达HULC 使MHCC-97L细胞的增殖能力增强,然而过表达miR-372 降低了细胞的增殖能力(P<0.001);低表达HULC 降低了HepG2 细胞的增殖能力,而低表达miR-372 增强细胞的增殖能力(P<0.001,图8C)。细胞流式术结果表明,过表达HULC 使MHCC-97L 细胞凋亡能力减弱,然而过表达miR-372 提高了细胞的凋亡能力(P<0.01);低表达HULC 提高了HepG2 细胞的凋亡能力,而低表达miR-372 减弱了细胞的凋亡能力(P<0.001,图8D)。Western blot 结果表明,与 pcDNA3.1-HULC 组相比,过表达 miR-372 可增加 MHCC-97L 细胞中E-cadherin 的表达并减少Vimentin 的表达(均P<0.01);与 si-HULC 组比,低表达 miR-372 可减少HepG2 细胞中E-cadherin 的表达并增加Vimentin 的表达(均P<0.01,图8E)。因此,我们证明了 HULC可通过HULC/ miR-372/CXCR4 轴发挥其对HCC 增殖、凋亡和EMT 的影响作用。

图5 miR-372 抑制HCC 细胞的增殖、促进HCC 细胞凋亡Figure 5 miR-372 inhibits HCC cell proliferation and promotes HCC cell apoptosis

图6 miR-372 抑制 HCC 细胞 EMT 进展Figure 6 miR-372 inhibits the EMT progression of HCC cells
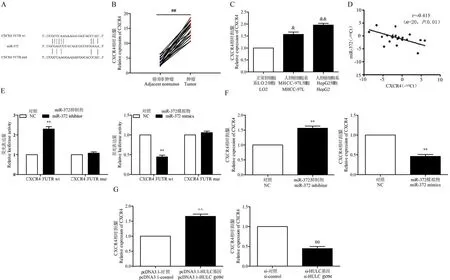
图7 CXCR4 在肝癌组织和肝癌细胞中的表达以及与miR-372 的关系Figure 7 CXCR4 expression in liver cancer tissues and liver cancer cells and its relationship with miR-372

图8 HULC/miR-372/CXCR4 轴调节肝癌细胞的细胞增殖、凋亡与EMTFigure 8 HULC/miR-372/CXCR4 axis regulates cell proliferation,apoptosis and EMT of liver cancer cells
3 讨论
在全球范围内,肝细胞癌(HCC)占所有肝癌病例的75%~85%[16]。据估计,2015 年中国有46.61万新诊断的肝癌患者,占全球所有肝癌病例的50%以上[17]。显然,肝癌对中国人民而言是沉重的公共卫生威胁[18]。因此,进一步研究肝癌进展的机制,为肝癌治疗提供更多的靶标。
人类基因组计划表明,大约90%的基因组被转录为非编码RNA[19]。据报道,非编码RNA,包括长非编码RNA(LncRNA,长度超过200 个核苷酸)和microRNA(miRNA,长度为 18 ~25 个核苷酸),是癌症和其他疾病的关键调节因子[20-21]。另外,LncRNA 和miRNA 能够调节几乎所有与癌症发生和发展有关的过程。LncRNA HULC 与癌症进展有关[7]。研究表明,HULC 在肝癌发生过程中发挥积极作用,例如,HULC 调节肝癌细胞中的脂质代谢[8]。而且 HULC 增强上皮-间质转化(EMT),促进肝癌细胞的转移[22]。本实验中,我们发现HULC 在肝癌组织和肝癌细胞中高表达,且HULC 可促进肝癌细胞增殖和EMT、抑制细胞凋亡。因此,HULC 有可能作为一类新型的潜在生物标志物或癌症的治疗靶标。
miRNA 是一类内源性、高度保守的、小的非蛋白质编码RNA,长度约为22 个核苷酸[23]。miRNA在许多生物中表达,并通过复杂的过程在调节靶基因表达中发挥作用[24]。miR-372 属于miR-371-372基因簇,位于染色体19q13.42。研究表明,miR-372可调节许多类型人类癌症中的细胞周期、凋亡、侵袭和增殖[25]。Lai 等[26]研究表明,miR-372 可能在非小细胞肺癌患者中转录后下调大肿瘤抑制基因2,从而导致肿瘤的发生和增殖。但是,miR-372 在肝癌中的作用尚不清楚,有研究表明 miR-372 在HCC 中低水平表达,并且通过负控制其靶基因ATAD2 发挥抗癌基因作用[27]。我们的实验结果表明miR-372 在肝癌组织和肝癌细胞中低表达,与上述文献一致。我们还发现,过表达miR-372 可能会显著抑制肝癌细胞的增殖和EMT,促进其凋亡。因此,miR-372 可能作为具有足够敏感性和特异性的一系列生物标志物的候选物,用于临床诊断HCC。
CXCR4 在20 多种肿瘤类型中过表达,并且在肿瘤生长、肿瘤侵袭性、癌细胞微环境相互作用和转移中起关键作用[28]。CXCR4 的存在与多种不同肿瘤实体的不良结局有关,包括血液系统恶性肿瘤、乳腺癌、肾细胞癌、妇科恶性肿瘤、胰腺腺癌和肝细胞癌[29]。本实验研究发现,CXCR4 在肝癌组织和肝癌细胞中高表达,低表达CXCR4 可能会抑制肝癌细胞的增殖和EMT,促进其凋亡。
因此,了解 LncRNA、miRNA 和 mRNA 之间的相互作用关系将有助于深入了解疾病。
本实验中发现 miR-372 可能靶向 HULC 和CXCR4,并且通过靶向关系进行负调控,进一步影响HCC 细胞的增殖、凋亡和 EMT。因此,HULC/miR-372/CXCR4 轴可能促进 HCC 细胞的增殖和EMT 进程、抑制细胞凋亡,这可能是将来HCC 有价值且有希望的治疗靶点。
综上所述,这项研究说明了肝癌中HULC、miR-372 与CXCR4 之间的分子相互作用。低表达HULC通过miR-372/CXCR4 轴抑制肝癌的发生和发展。这些结果表明,HULC/miR-372/CXCR4 轴可以用作缓解肝癌进展的有希望的治疗靶标。
