泡果沙拐枣种子萌发对模拟降雨间隔的响应
2020-11-09朱成林李晓梅阿依帕夏居麦赵晓英成鹏
朱成林,李晓梅,阿依帕夏·居麦,赵晓英,成鹏
(1.新疆师范大学生命科学学院,新疆 乌鲁木齐 830054;2.山东省潍坊第七中学,山东 潍坊 261021;3.乌鲁木齐市气象局,新疆 乌鲁木齐 830002)
泡果沙拐枣(Calligonumjunceum)是蓼科沙拐枣属的超旱生灌木,主要分布在新疆精河、托克逊、吐鲁番等地,苏联中亚、蒙古也有分布。在达坂城荒漠的有些地段,泡果沙拐枣为优势种或共建种,对维持荒漠生态系统的稳定性具有重要作用,它也是干旱荒漠地区防风固沙和植被恢复的重要植物[1]。
水分是种子萌发的先决条件,在干旱荒漠区,降雨量(水分)是植被能否成功建立的主要限制因子[2,3]。种子成熟后严重脱水,只有在合适的水分条件下经过吸胀,才能启动萌发。有些植物启动种子萌发的最低土壤水分含量较高,而有些植物启动种子萌发的最低土壤水分含量很低,这一机制被认为是其种子适应生境特殊气候和地形的一种生态对策[4]。降雨的频率和降雨量的大小会影响种子的萌发率和萌发速率[5,6]。种子萌发对降雨变化的响应以及其适应特征会直接影响后续的幼苗建立与补充,进而影响种群的更新动态。因此,加强干旱区降雨格局变化对种子萌发影响的研究,对干旱区植被的发展趋势以及种质资源保育更新均具有重要的意义[7]。在干旱荒漠中,土壤表面不能长期保持湿润,湿润和干燥状态交替出现,这可能有利于种子快速萌发。植物为了在干旱区生存,种子必须适应这种吸湿—回干循环[8]。种子附属物的形成是植物生活史长期进化的重要特征,能够在种子受到机械损伤或者其他风险时保护种子。种子附属物除了对种子传播和扩散进行调控外,还对适宜环境条件下种子的萌发起着重要作用[9,10]。附属物的吸水保水作用是荒漠植物对环境适应的一个重要特征[11]。
有关泡果沙拐枣的抗旱性[12],抗逆性[13],种子催芽处理[14],种子休眠的解除[15],形态、遗传、物候以及分布格局[16,17]、群落多样性[18]等方面的研究已有报道。吸湿——回干处理对播种前提高种子活力十分有益[19]。一些研究中,通过干湿交替来模拟降雨的频率和降雨间隔。研究表明,吸湿—回干处理的种子比未经过处理的提前萌发[8],而且随着回干时间的延长萌发率增高[20],我国西北地区旱生植物红砂(Reaumuriasoongorica)种子,通过增加单次降雨量以及延长降雨间隔(10 d),其萌发率和出苗率也增高[7]。但浸泡次数过多时银合欢(Leucaenaleucocephala)种子活力降低,从而影响萌发率[21]。对于有些物种吸湿—回干处理时间的延长也有可能诱导种子休眠[5,22],如鄂尔多斯荒漠植物赖草属的赖草(Leymussecalinus)种子[5],达坂城荒漠地区霸王(Zygophyllumxanthoxylon)种子[22]和羊草(Leymuschinensis)种子[23]。黑麦草(Loliumrigidum)种子在夏季和秋季经雨水吸湿—回干后,种子休眠得以解除虽然促进了种子的萌发,但种子活力会受到一定的影响[24]。
本研究在实验室模拟自然环境中的降雨间隔时间和土壤水分含量,对泡果沙拐枣种子进行处理并观察其萌发特性,为植被更新过程中种子适应极端干旱环境的生态机制提供理论依据。
1 材料与方法
研究区位于达坂城柴窝堡林场后的泡果沙拐枣自然群落(43°31′37.11″ N;87°53′23.6″ E),海拔1 103 m。该地区属于温带大陆性干旱气候,年平均气温6.6 ℃,年平均降雨量71.7 mm,年平均蒸发量212.2 mm[25]。达坂城的植被为荒漠植被,群落组成主要有霸王、泡果沙拐枣、刺木蓼(Atraphaxisspinosa)、膜果麻黄(Ephedraprzewalskii)、木本猪毛菜(Salsolaarbuscula)等。研究区的气温和降雨量数据来自乌鲁木齐气象局柴窝堡站(图1)。
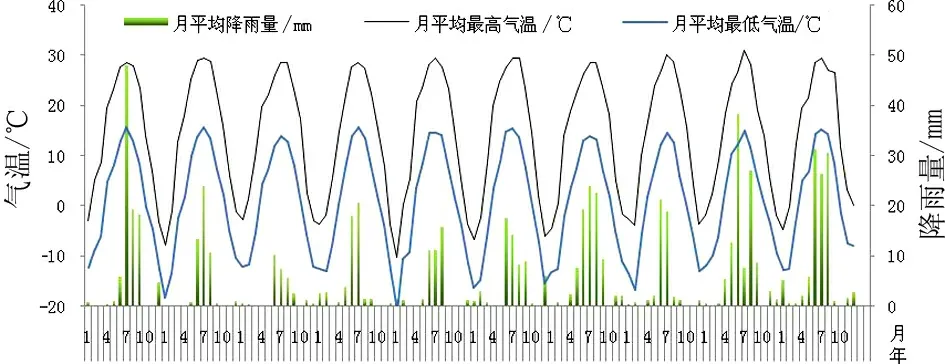
图1 达坂城地区2007—2016年月平均降雨量、最高和最低气温
于2015年6月中旬种子成熟期,在自然群落中每隔10 m随机选取30丛以上泡果沙拐枣植株,采集成熟的种子,将采集的种子充分混匀,置于通风干燥的室内晾干,选取健康饱满的种子备用。每个培养皿(直径为100 mm)底部放25粒种子,4次重复。分别用100 g沙子覆盖(约1 cm厚)。沙子取自原生境,过0.5 mm孔径的筛子,洗净,105 ℃下烘干,置于不同的实验条件下,以种子长出2 mm胚根视为萌发,每天检测1次,并且将已萌发的幼苗捡出,持续观测30 d。实验结束后对未萌发的种子用TTC法检测生活力。萌发率=萌发种子数/有活力的种子数×100%[26]。
1.1 种子的物理特性
随机选取800粒种子,经测量表明,种子百粒质量为3.24±0.06 g,长、宽、高分别为12.16±0.34 mm、10.21±0.22 mm和10.21±0.19 mm。
1.2 不同降雨间隔种子的萌发
于2017年7月,将种子低温层积7个月,解除休眠进行萌发试验,每个培养皿第1次加14 mL蒸馏水,降雨间隔设为5、10、20、30、60 d(模拟夏季降雨之间的干旱期),之后每过一个降雨间隔加1次水,置于室温下培养。
1.3 不同降雨量种子的萌发
采用ECH2O土壤水分检测系统(Decagon ,USA)记录地表和土中3~5 cm的土壤水分含量。每2 h记录1次,取当日的平均值作为当日的土壤含水量(图2),根据测量结果设置土壤水分含量的梯度。
选取试验田埋藏5个月的泡果沙拐枣种子,晾干11周,进行萌发试验。每组培养皿分别加入0.5、1、2、5、8、11、14、17、20、23、26、29 mL蒸馏水,计算出沙子的含水量分别为0.5%、1%、1.5%、2%、5%、8%、11%、14%、17%、20%、23%、26%、29%。将培养皿置于25/15 ℃(适温)的变温培养箱内培养,每天称质量,补充因蒸发散失的水分,保持恒定的湿度,每次检出已萌发的种子时立刻称质量记录,下次加水时总质量减去已萌发的幼苗的质量。

图2 2015—2016年研究区生长季种子埋放生境地表和土中(3~5 cm)的土壤水分含量
1.4 种子附属物与种子吸水脱水的关系
实验室条件下,选取室内贮藏6个月的完整泡果沙拐枣果实360粒。设置3组处理,附属物完好120粒,用针刺破附属物120粒,完全去除附属物120粒。每皿30粒,4次重复。将处理好的果实称质量,分别置于装有蒸馏水的培养皿中,检测附属物对种子的吸水量和吸水时间。吸水开始0、0.5、1和2 h后时将果实分别从培养皿取出,用电子天平称质量,之后每隔4 h称质量,直到恒质量为止,检测附属物对果实的脱水量和脱水时间。
种子累积吸水率(%)=种子吸水量(g)/吸水前种子质量(g)×100%
种子累积失水率(%)=种子吸水量(g)/失水前种子质量(g)×100%
1.5 数据处理与分析
利用SPSS 19.0统计分析软件和Excel 2010软件对数据进行处理和绘图,结果以平均值(Mean)±标准误(SE)表示。采用单因素方差分析(One-way ANOVA)、最小显著性差异法(LSD)和新复极差法(Duncan)检验分析试验数据。
2 结果与分析
2.1不同降雨间隔种子的萌发
由图3可知,泡果沙拐枣种子经低温层积7个月后(无降雨间隔)在室温条件下萌发率为69%。降雨间隔5、10、20、30、60 d后萌发率分别为47%、31.2%、24.4%、24.3%、16.5%,显著低于未经回干处理的种子(P<0.05),其降雨间隔60 d萌发率最低,为16.5%,显著低于降雨间隔5 d和10 d(P<0.05)。降雨间隔0~30 d,从第2天开始有种子萌发,降雨间隔60 d,从第3天开始有种子萌发,集中萌发时间为1周,之后的30 d内无种子萌发。

图3 泡果沙拐枣不同降雨间隔(0~60 d)的种子萌发率(平均值±标准误)
2.2 不同土壤水分含量种子的萌发
泡果沙拐枣在不同土壤水分含量条件下的萌发情况见图4,在土壤水分含量0.5%时无种子萌发,1%时萌发率最低,为2.4%,土壤水分含量1.5%、2%、5%、8%、11%、14%、17%、20%、23%、26%和29%时的萌发率,分别为13.5%、42.4%、75.3%、73%、83.2%、95.2%、91.7%、77.6%、76%、73.6%和75%,从1%极干旱条件到所测沙子的饱和含水量29%的条件下均有种子萌发。土壤水分含量≥5%时萌发率均≥73%。14%和17%具有相对较高的萌发率,分别为95%和91%,其差异显著(P<0.05)。土壤水分含量在5%~11%和20%~29%范围内的种子萌发率差异不显著(P>0.05)。当土壤水分含量1%~1.5%时,从第4天开始有种子萌发,萌发持续时间为10 d,当土壤水分含量5%~29%时,从第2天开始有种子萌发,第4天萌发指数最高,集中萌发时间为1周,之后无种子萌发或只有5%的种子萌发。

图4 不同土壤水分条件下(0.5%~29%)泡果沙拐枣种子萌发率(平均值±标准误)
2.3 种子吸水脱水特性
由图5可见,刺破附属物的果实在0~12 h期间吸水量快速增加,24 h后吸水量达到饱和状态;完整果实在0~36 h内吸水缓慢,之后吸水量快速增加,50 h后达到饱和状态;无附属物种子前12 h快速吸水,之后达到基本饱和状态。具附属物果实的吸水量显著高于无附属物果实,二者之差代表了种子附属物吸水量的变化特征。
吸水饱和之后,完整果实、刺破附属物果实和无附属物果实在室温下脱水的速率存在显著差异。由图6可见,随着时间的增加,3种处理的果实本身的含水量均呈下降趋势,但是下降的速率不一,依次为完整果实<刺破果实<无附属物果实。其中无附属物的果实约12 h就从吸水饱和状态恢复到原重,完整果实和刺破果实0~12 h缓慢脱水,之后迅速脱水,质量到48 h后才恢复到吸水前状态。
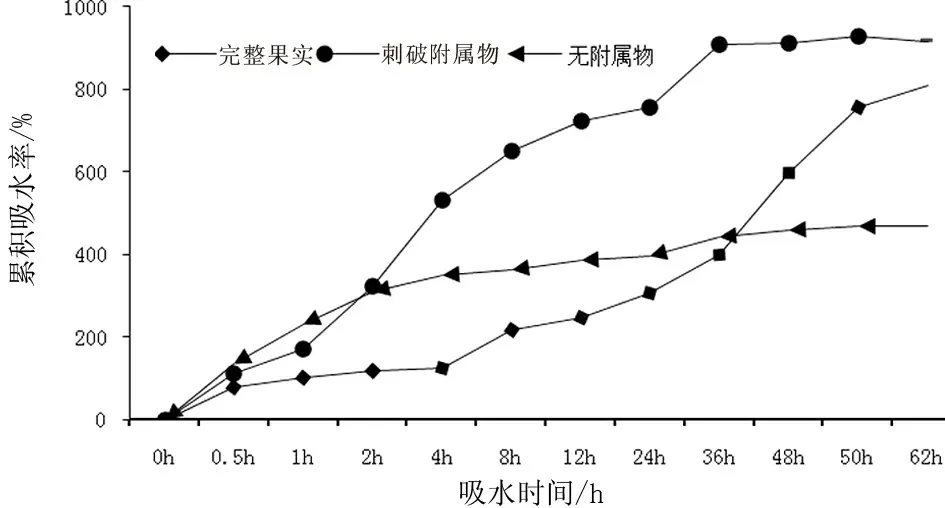
图5 泡果沙拐枣完整果实、刺破附属物和去除附属物后果实吸水质量变化和累积吸水率(平均值±标准误)

图6 泡果沙拐枣完整果实、刺破附属物和去除附属物后果实脱水质量变化和累积脱水率(平均值±标准误)
3 讨论与结论
3.1 附属物对种子吸水脱水的影响
干旱荒漠生境中土壤种子库主要集中在0~2 cm表层。由于春季风多,水分和温度波动性大,大多数表层土壤种子库处于干湿交替的环境中,种子萌发受到干旱胁迫。因此,作为种子特殊结构的附属物具有一定吸水保水功能被看作是干旱荒漠植物种子适应环境的一种生态机制[11]。有附属物的泡果沙拐枣种子吸水率均高于无附属物的种子吸水率。二者之差代表了种子附属物吸水率的变化特征。这表明,附属物的吸水能力远高于种子自身的吸水能力,其吸收的水分在泡果沙拐枣果实的吸水过程中占主导作用。而脱水率和脱水速率低于无附属物的果实,这表明,附属物具有较强的保水能力,在干旱脱水的环境中能使种子保持湿润,缓慢脱水。附属物的保水作用可能减缓干旱胁迫对种子萌发的不利影响,使种子在干旱少雨的荒漠环境中有效利用有限的降雨而不至于快速脱水死亡,保持种子活力。西北地区草本植物欧夏至草(Marrubiumvulgare)具有附属物的集合繁殖体能保护种子被捕食,附属物比种子吸收更多的水分并能更长时间的保留水分防止种子迅速干燥[27]。小车前(Plantagominuta)种子表面上的黏液物质具有吸水保水的特性,在干湿交替剧烈的干旱荒漠区通过调节水分来起到调节种子萌发时机的作用[11]。
3.2 降雨间隔长,仍有种子萌发
本研究中,降雨间隔延长到60 d时仍有部分种子萌发,说明种子在吸胀的状态下可长时间保持活力,远长于同一群落中旱生灌木霸王种子,霸王的降雨间隔大于14 d萌发率已明显降低[22]。任君等在腾格里沙漠所做的3次短时间(最长回干时间10 d)吸湿回干处理对泡果沙拐枣种子萌发无促进作用[6]。水分是荒漠植物种子萌发的主要限制因素,泡果沙拐枣种子在1%的低土壤含水量条件下能启动萌发(图4),而在2%时萌发率高于锦鸡儿属等其他旱生植物[4]。达坂城地区一次降雨过后土壤水分含量可达到2%~13%(图2),这种低水分条件完全满足休眠已解除的泡果沙拐枣种子的萌发。这表明长期进化过程中,泡果沙拐枣种子具备了能在长期干旱环境下生存下来的机制策略。在长期干旱条件下泡果沙拐枣种子仍能持续产生幼苗,可能与其泡状瘦果的形态特征有关,其种子外有一层淡红色泡状膜,包围整个瘦果的外部,而且种皮木质化,这种特殊的结构有利于种子的吸水和贮水。本试验表明,泡果沙拐枣种子吸水率很高,带附属物的果实吸水约是自重的11倍,可见其附属物的保水作用,以此可以适应达坂城2个月无降雨的干旱环境。种子的特殊形态结构和萌发机制确保了许多干旱荒漠植物在合适的时间与地点下萌发与幼苗建成[2]。
本研究中,无降雨间隔时泡果沙拐枣种子萌发率较高为70%,试验开始的第2天就有种子萌发,萌发集中在第1周,后续再无种子萌发,说明种子一旦解除休眠,只要温度适宜,之后的降雨会推动种子萌发。可能因为泡果沙拐枣种子种皮坚硬,具有休眠现象,而解除休眠需要较长时间的湿冷贮藏,因此种子休眠解除后会立即萌发,避免之后的干旱使幼苗灭绝。
3.3 降雨间隔长,萌发率降低
在达坂城地区降雨量少(年平均降雨量71.7 mm),蒸发量大(年平均蒸发量212.2 mm)[25],一次降雨可能很快蒸发掉,因此不足以使土壤中的种子萌发,本试验表明,短期吸水—回干(降雨间隔5 d)对种子萌发的影响不大,这可能是因为回干后的泡果沙拐枣种子能够保持着短期吸水后的生理变化,从而保证再次吸水后的快速萌发。类似的现象在降雨偏少,蒸发量大的黄土丘陵地区旱生植物杠柳(Periplocasepium)种子中也有发现[28]。然而降雨间隔延长20~60 d时萌发率显著降低,其未萌发的种子活力高于90%,说明种子进入休眠。这表明大部分种子通过进入休眠来适应这种持续干旱的环境保持活力。鄂尔多斯荒漠植物赖草种子经过8 d的回干处理后萌发率降低,种子进入休眠[5]。然而,我国西北地区旱生植物红砂种子,经增加单次降雨量以及延长降雨间隔(10 d),其增高萌发率和出苗率[7]。还有研究指出,延长降雨间隔时间可以增加旱生植物红砂的幼苗忍耐干旱胁迫持续的时间[29]。本研究中延长降雨间隔时间种子进入休眠,可能的原因是,随着降雨间隔时间(干旱)的延长其土壤水分含量减少,种子脱水,新陈代谢缓慢,种子萌发率降低,进入休眠。另外,随着降雨间隔时间的延长,已变软的泡果沙拐枣种皮重新坚硬,阻碍胚根突出种皮,从而抑制萌发。泡果沙拐枣种子野外埋放以后,经过秋季、冬季和早春季的冷湿环境,种子会解除休眠,而经过夏季的高温和干旱环境种子会进入休眠[30],本研究结果进一步验证了这一点。种子进入休眠的好处在于,泡果沙拐枣多分布于干旱半干旱的荒漠化砾石地以及沙地。达坂城是干旱荒漠地区,降雨量很少(图1),第1次降雨和第2次降雨之间的间隔较长,而且风大,水分蒸发量高,土壤水分的有效性低。在此环境条件下,若种子不能利用有限的水分抵抗干旱,可能导致幼苗全部死亡。因此植物通过休眠对策尽量分摊这种环境条件的风险,在干旱荒漠区种子通过这种适应策略可以避免种子在炎热和长期干旱环境下萌发之后幼苗的死亡。这种萌发对策被认为是长期生活在干旱地区的植物适应环境的必然结果[31]。
3.4 低土壤水分含量下仍然有种子萌发
本研究中,土壤水分含量1%时仍有种子萌发,这说明启动泡果沙拐枣种子萌发所需的土壤水分含量很低,这比启动旱生植物白皮锦鸡儿(1.25%)、甘蒙锦鸡儿(3.75%)和中间锦鸡儿(2.50%)种子萌发的最低土壤含水量低。土壤水分含量2%时萌发率达到40%,这说明泡果沙拐枣种子萌发对土壤含水量的要求不是很严格。较低土壤含水量能够启动萌发的原因可能是,第一、与种子形态结构和遗传因素有关,泡果沙拐枣种子具附属物,包围整个瘦果的外部,而且种皮木质化,这种特殊的结构有利于种子的吸水和较长时间的保水;第二、与泡果沙拐枣种子所生存的环境条件有关,干旱地区水分的补充主要靠降雨并且降雨的时间和空间分布不均匀,年降雨波动性较大,在这样一个选择压力下,泡果沙拐枣种子萌发对策需要对降雨做出响应;第三、达坂城荒漠降雨量很少,降雨时间集中在夏季(图1),但是夏季土壤温度高(65 ℃)、风速较大(7 m·s-1)、蒸发量高(889.7 mm)[25]、土壤水分含量的波动性大,降雨后土壤含水量只能达到2%~13%(图2),在这样的生存环境条件下一场降雨过后,解除休眠的种子能够在较低土壤含水量下启动萌发,且萌发速度较快,以适应降雨量少的干旱荒漠环境。在干旱地区,休眠解除的种子当水分适宜时,种子快速萌发出土是该物种能否成功建立种群和自然更新的关键。较早萌发的物种具有很大的竞争优势,它们能够提前获得有限的营养和水分资源等来提高存活率,从而减少物种灭绝的风险。
3.5 土壤水分含量的高低与萌发速率的关系
种子萌发时响应不同的环境暗示,萌发对水分的适应主要体现在对土壤湿度的响应。干旱荒漠植物种子在应对水分亏缺的环境压力时,表现出不同的生态适应对策[2]。本研究中土壤水分含量1%~1.5%时,从第4天开始有种子萌发,萌发持续时间为10 d,说明土壤水分含量极低时种子萌发速率受到限制,萌发策略倾向于缓慢萌发,这可能是因为种子为了避免在达坂城极端干旱条件下萌发之后长期干旱导致幼苗全部死亡的措施。水分亏缺条件下霸王种子萌发率,低萌发速率缓慢,这是适应极端干旱环境的一种策略[32]。种子萌发速率的调控作用确保了更多的泡果沙拐枣种子在适宜的土壤水分含量条件下萌发。土壤水分含量5%~29%时,从第2天开始有种子萌发,第4天萌发指数最高(暴发),集中萌发时间为1周,之后无种子萌发或只有5%的种子萌发,这说明土壤水分含量适宜时种子萌发策略倾向于快速萌发,避免由萌发持续时间延长而导致幼苗死亡的风险。这确保大部分种子只有在土壤水分含量适宜时启动萌发,为种群在风险环境中稳定发展打下坚实的基础。
