RecA蛋白介导同源重组的步进式链交换
2020-11-06黄星榞隋明宇侯文清李明陆颖徐春华
黄星榞 隋明宇 侯文清 李明 陆颖 徐春华†
1) (中国科学院物理研究所, 北京 100190)
2) (中国科学院大学, 北京 100049)
3) (兰州大学物理科学与技术学院, 兰州 730000)
1 引 言
DNA是生命体中重要的遗传物质, DNA的正常复制、转录和翻译是细胞得以稳定繁殖的基础.当DNA受到细胞内部或外部因素的威胁时, 会产生诸如DNA断裂等结构性损伤, 损伤严重时可导致癌变或细胞凋亡[1]. 细胞有一系列机制能够修复这类损伤[2], 其中有一种以同源DNA为模板, 进行严格而精确修复的方式, 称之为同源重组. 同源重组由重组酶介导发生, 原核生物中的RecA和真核生物中的Rad51, DMC1是重组酶的代表, 它们在其他蛋白共同作用和促进下开展同源重组过程.下面以本文的研究对象RecA重组酶为例对同源重组的过程进行简单说明:当DNA的磷酸骨架断裂, 首先RecBCD会处理断链末端, 沿一定方向剪切DNA断链位置, 暴露出一段有3’末端的单链.在腺苷三磷酸(ATP)存在时, RecA与单链DNA形成核蛋白丝结构 (nucleoprotein filament), 搜寻游离双链DNA上的同源序列并进行同源匹配, 匹配成功后核蛋白丝入侵双链DNA并进行链交换形成三链复合体. 最后由RuvABC处理三链复合体, 完成同源重组. 除了修复 DNA 的损伤, 保证遗传信息的稳定性, 同源重组也能促使同源染色体上的同源碱基互换, 引入基因多样性促进生物更快适应环境的变化.
RecA蛋白是大肠杆菌中recA基因的表达产物, 分子量 38 kD, 共含有 383 个氨基酸残基. RecA蛋白的中心区域是包含240个氨基酸残基的ATP水解酶核心, 除此之外, 还有带30个残基的N端和82个残基的C端, N端和C端均带有正电, 它们可以与DNA骨架发生相互作用. RecA蛋白上这两处带有大量正电的区域, 称为第一结合位点和第二结合位点. 单链DNA与多个RecA单体的第一结合位点结合, 形成具有周期螺旋结构的核蛋白丝. 核蛋白丝中, 每个RecA单体能够结合三个碱基形成基本结构单元, 这三个碱基称作一个triplet. 根据Chen[3]等的X射线晶体结构结果, 一个triplet内部的碱基间距为0.34 nm, 与B-DNA结构中的碱基距离相同, 但两个triplet的间距为0.78 nm, 核蛋白丝包裹的单链呈现拉伸状态, 平均的碱基间距为0.51 nm. 核蛋白丝的一个周期包含6.2个RecA单体和18.5个碱基, 呈右手螺旋结构. 在同源重组产生的三链结构中, 我们称核蛋白丝包裹的单链为入侵链(incoming strand), 同源双链中与入侵链配对的链为互补链(complementary strand), 与入侵链序列相同的链为被置换链(outgoing strand).
同源重组过程中的序列识别和链交换的分子机制近年来一直是研究热点[4−6], 而链交换的步长是链交换过程中的一个重要表象, 它可能会是链交换分子机制的一种体现. 因而对链交换步长的测定是研究RecA分子机制的重要一环. Ragunathan等[7]使用荧光共振能量转移(FRET)方法,通过统计不同长度同源DNA链交换反应完成的时间, 推算出链交换是以每步3 bp进行的. Lee等[8]使用 DNA 帘 (DNA curtain)方法, 通过统计标记荧光的DNA双链在同源或错配序列上的驻留时间, 计算出无论是RecA, Rad51还是DMC1体系,换链步长均是3 bp. 以3 bp为步长的说法很容易被人们接受, 因为一个RecA蛋白单体正好结合3 bp DNA, 但也有研究人员对步长提出其他看法.Prentiss课题组[9−12]参考核蛋白丝的晶体结构, 对链交换过程进行了数学建模, 发现在链交换发生前的同源序列匹配过程中, 互补链处在能同时与入侵链和被置换链配对的位置, 当配对长度为9 bp时体系处于最稳定状态, 配对成功后发生换链, 同源序列识别和链交换均是以9 bp进行的. 我们前期工作首次使用单分子磁镊, 结合DNA 发卡结构(DNA hairpin)的底物设计, 将反应信号放大[13],直观测量了链交换的反应步长. 经统计发现每一步链交换的步长均为3 bp的整数倍, 其中最概然长度约为9 bp, 与Prentiss等的理论结果相同, 但与3 bp的结论矛盾. 磁镊实验可以得到最直观的实验结果, 但 9 bp的步长也可能是由 3个 3 bp构成的,只是受到仪器的时间分辨率和空间分辨率的限制而没有观察到. 因此, 目前同源重组的链交换步长依然没有定论. 我们认为掌握了链交换的步长, 对于发掘同源识别和链交换的机制有着重要的意义,可以进一步完善对同源重组过程的理解. 因此, 本文用一系列实验来探索RecA介导的同源重组链交换步长, 以期从多方面、多角度验证9 bp为链交换的最概然长度, 而非3 bp. 首先运用酶切保护实验, 证明了链交换的步长不是3 bp, 再使用单分子磁镊测量了一定错配程度序列下的换链总长度, 侧面证实了链交换的步长为9 bp. 最后通过局域连续错配序列, 再次证明链交换步长不是3 bp, 同时也分析了这9 bp内错配碱基的分布是如何对链交换造成影响的. 本文是使用单分子实验对同源重组过程探究的新的尝试, 运用错配碱基和酶切保护等手法, 为研究提供了新思路.
2 实验材料、设计与操作
2.1 RecA、内切酶及反应溶液
RecA 蛋白购自 New England Biolabs (NEB)公司. 在磁镊和荧光实验中, 核蛋白丝的链交换反应在 RecA reaction buffer中进行, 具体成分为70 mM Tris-Ac (1 M=1 mol/L), 10 mM Mg(Ac)2,100 mM NaAc, pH=7.4. 为了保持核蛋白丝的稳定, 每次进液时需要额外补充 5 mM的ATPgS.酶切时使用了NEB公司的DNaseI内切酶, 酶切环境为 DNaseI reaction buffer.
2.2 DNA的制备
磁镊实验中所使用的所有DNA序列均购自生工生物工程(上海)股份有限公司, 核苷酸链均稀释在 TE buffer中. Hairpin DNA 的制备过程参考我们之前的工作[13].
2.3 磁镊方法的原理和操作
磁镊是一种单分子力谱方法, 可以通过追踪磁球的高度直接观测蛋白质或DNA分子的结构变化[13−17], 实验中使用磁镊结合 DNA hairpin结构,对链交换长度进行直接测量. DNA hairpin是两条具有回文序列的单链, 在小力下能够自然退火形成B-DNA双链. 用来代表同源重组过程中的互补链和被置换链. 链交换的发生导致互补链和被置换链打开, 根据打开的长度计算可计算链交换碱基数.
首先将盖玻片用丙酮/甲醇超声清洗30 min,再浸泡在浓硫酸∶双氧水体积比为7∶3配制的食人鱼洗液中, 于 95 ℃ 水浴 2 h, 接着放置在 1% 的APTES 溶液中修饰 2 h, 具体步骤可参考[18]. 冲净APTES后, 在1%的戊二醛溶液中修饰8 h, 取出冲洗吹干后密封保存. 在实验前取出玻片, 用双面胶和麦拉膜制成封闭的反应池, 反应池下表面用地高辛抗体修饰. 玻片的表面处理和钝化可见参考文献[19].
DNA连接完毕后, 将核蛋白丝加入到体系中使反应进行, 通过软件记录磁球位置和时间的关系, 绘制反应曲线, 使用数据处理软件收集换链长度等信息.
2.4 酶切保护实验的原理和操作
酶切保护实验涉及单分子荧光成像和酶切. 为了避免激光将反应池内所有荧光激发, 增加实验的背景噪声, 使用TIRF场仅激发玻片几百纳米范围内的荧光分子[4], 通过软件处理实验图像统计酶切前后荧光分子的数量.
实验所用盖玻片的清洗方法直到修饰APTES均与磁镊相同, 在APTES处理后加入PEG钝化玻片表面, 其中掺入1%的biotin-PEG用来与链亲和素修饰的DNA进行特异性链接. 玻片修饰完毕后制成反应池, 连接单链DNA, 加入RecA单体和ATPgS共同孵育15 min制备核蛋白丝[20]. 随后在体系中加入末端标记Cy3荧光的双链DNA.在打开激光录像时加入抗淬灭体系延长荧光持续时间[20].
双链首先与核蛋白丝反应 15 min, 15 min 内大部分核蛋白丝都已参与反应. 随后在反应体系中加入DNaseI内切酶进行酶切处理. 虽然ATPgS环境下链交换结束后核蛋白丝不易水解, 但为了保证核蛋白丝的稳定性, 酶切时间不宜过长, 我们发现30 min的酶切已能产生明显的对比效果, 因此酶切时间选定为30 min. 使用软件对剩余荧光点数进行计数. 对比酶切前后的荧光点数, 可计算出发生完全链交换反应的比例.
2.5 核蛋白丝的制备
磁镊实验中加入的核蛋白丝在反应池外完成制备. 在RecA 反应buffer中将RecA蛋白∶核苷酸以摩尔比1∶3的比例混合均匀, 随后补充5 mM ATPgS, 在 37 ℃ 下孵育 5 min, 取出混合液后即可放入反应池中进行实验. 在荧光实验中, 核蛋白丝在反应池中合成, 取 RecA 原液 2 µL, 与 100 µL的RecA 反应buffer充分混合, 补充5 mM ATPgS后加入到反应池中, RecA与反应池中的单链DNA会自组装形成核蛋白丝结构.
3 实验结果与分析
3.1 验证链交换反应是否以3 bp发生
首先要验证的是链交换是否以3 bp进行. Lee等[8,21]的 DNA curtain 实验表明, 在 ATPgS条件下链交换可以跨过1个错配碱基, 但会被两个连续错配碱基阻挡. 如果3 bp是换链的基本长度, 既然可以跨过一个错配碱基, 那么如果每一个triplet内都含有一个错配碱基时, 也应该可以成功换链.以此思路我们设计了带有部分错配碱基的DNA序列, 序列全长69个碱基, 前45个碱基为同源序列, 后24个碱基为33%错配的序列, 错配碱基均匀分布在每个triplet中, 即每个triplet包含一个错配碱基, 如图1(a)所示, 观察链交换反应是否可以跨过所有错配. 同源序列长度大于18 bp, 满足同源重组链交换反应发生的条件[22]. 同源碱基提供DNA与核蛋白丝的初始反应位点并促使换链反应进行, 形成三链结构.
荧光分子标记在双链中的互补链上, 位于错配一端的末尾位置. 根据原子力显微镜结果[23], 在同源重组过程中, 如果链交换过程遇到阻碍, 剩余的双链会以接近垂直的角度从核蛋白丝上脱离, 尾部暴露在外部环境中, 可以被内切酶处理. 荧光分子随着被水解的核苷酸链脱离三链结构, 荧光点数减少. 如果链交换反应完全发生, 互补链与入侵链形成的杂合DNA被核蛋白丝包裹, 可以保护末尾的核苷酸不被水解, 荧光点数不变. 根据该原理设计了酶切保护实验, 观察核蛋白丝的链交换反应能否容忍错配碱基, 完成整条链的交换. 为了验证核蛋白丝对酶切保护的效果, 使用了完全匹配的序列作为对照组, 示意图如图1(b)所示. 酶切实验整体分为全匹配对照组和33%周期性错配实验组.

图 1 全匹配酶切保护实验的示意图及酶切结果 (a) 包含错配序列的酶切保护实验示意图, 蓝色部分代表同源序列, 红色部分代表错配序列; (b) 在全匹配序列的情况下酶切保护实验示意图; (c) 酶切实验结果, 分别为全匹配序列和33%错配序列的酶切情况(误差线代表统计误差), 酶切时间为30 minFig. 1. The enzyme protection assay: (a) The diagram of the enzyme protection assay with mismatch sequence, blue strands represent homologous sequence and red strand represents mismatched sequence; (b) same as (a) when using completely homologous sequence;(c) the results of enzyme protection assay.
酶切保护实验结果如图1(c)所示, 在全匹配对照组实验中, 酶切后荧光点数为酶切前的96%,基本没有发生变化. 而33%错配的实验中, Cy3荧光点数在酶切后降为酶切前的19%, 说明链交换反应在中途停止, 互补链和被置换链以B-DNA的形式脱离核蛋白丝, 暴露在溶液中的DNA末端被DNase I切除, Cy3分子脱离三链结构扩散到了溶液中, 导致荧光点数减少. 与对照组对比, 33% 错配与全匹配的反应现象完全不同, 错配序列没能像全匹配实验一样彻底完成链交换. 结果说明, 当每个triplet内平均分布一个错配碱基时, 链交换过程无法克服错配碱基带来的阻碍而中止, 得到链交换可能不是以3 bp为步长发生的初步结论.
3.2 使用单分子磁镊观察不同错配程度下的链交换长度
由于酶切保护方法只能验证换链反应是否完全进行, 无法得到链交换终点的准确位置, 观测的现象有一定局限性. 我们使用磁镊方法来定量测量33%错配序列下的链交换停止的位置时[13], 意外得到了步长的信息. 磁镊实验的设计如图2(a)所示. 为了保证信号的信噪比, 同时防止较大的力对DNA hairpin的稳定性造成影响, 实验中选择了7 pN的拉力. 在使用错配序列进行实验前, 先用全匹配序列验证DNA长度变化与换链碱基数的对应关系. 全匹配序列长度为69个碱基, 根据FJC模型[24,25]计算在7 pN下单链DNA的平均碱基间距为0.31 nm, 即hairpin岔口位置每有一个碱基完成链交换, 磁球升高 0.51+0.31=0.82 nm.全匹配序列的实验结果如图2(b)所示, 磁球的高度升高 57 nm, 根据换算对应约 69 bp 的换链, 同源序列对照实验说明长度与碱基的换算关系正确,接下来的磁镊实验同样使用此对应关系进行换算.
33%错配实验所用的DNA序列与酶切保护实验的序列相同, 前45个碱基为同源序列, 后24个碱基为错配碱基均匀分布的33%错配序列, 定义第45个碱基位置为界面, 作为同源序列与错配序列的分界线. 在ATPgS条件下, 假如换链沿着入侵链的 3'到 5'方向进行, 即从 DNA hairpin岔口位置开始向另一侧换链, 将得到如图2(c)所示的实验曲线. 对链交换信号处理后, 获得了换链总长度的统计图, 如图2(d)所示. 从散点图(图2(d))可以看出, 换链总长度集中分布于界面位置, 少部分分布于界面之前 9 bp (–9)的位置, 说明在 33%错配程度的情况下, 链交换反应无法跨过错配碱基, 导致链交换中止. 磁镊方法定量的给出了换链总长度, 使我们发现了链交换完全无法跨过33%周期错配序列这一新的现象. 如换链以3 bp为单位进行, 结合链交换可以跨过1个错配碱基的结论, 应有部分反应可以跨过界面, 但实验结果显示几乎所有换链均停止在界面之前, 这与酶切保护实验的结果是一致的, 说明链交换可能不是以3 bp为步长发生的. 而另一个链交换终点的峰值位于界面之前9 bp(–9), 说明这部分链交换被阻挡在了界面之前 9 bp, 而–9 到界面之间并没有明显的峰, 这很可能说明链交换的步长即为9 bp, 终点在–9位置的换链正好比终点在界面位置的换链少了一步.
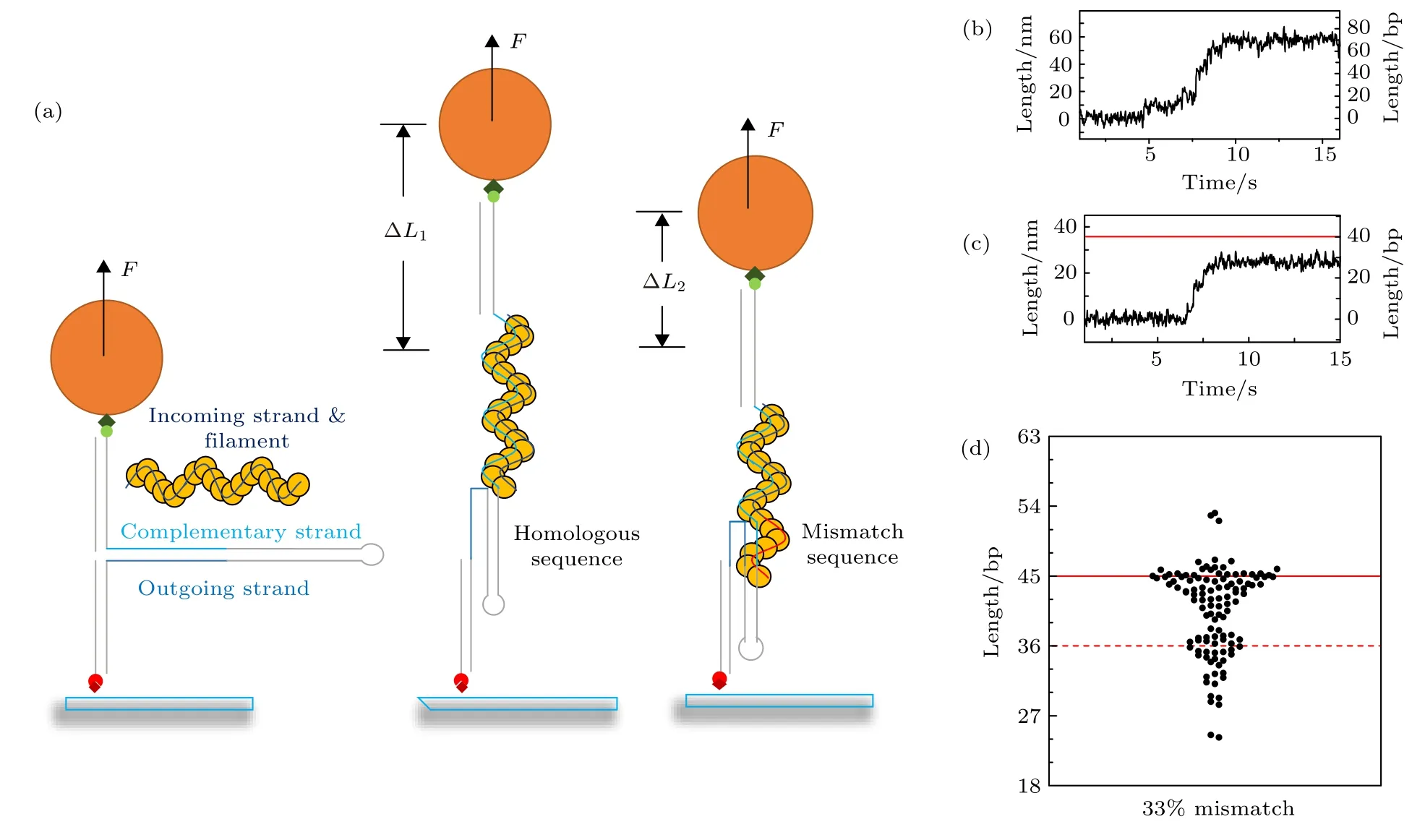
图 2 用磁镊定量测量不同匹配程度序列下的换链长度 (a) 磁镊实验的示意图, 红色链代表错配序列, 蓝色链代表同源序列,从左到右分别对应链交换反应发生之前, 全匹配序列的链交换反应, 和链交换被错配序列阻挡三种现象; (b) 全匹配序列下的换链曲线, 换链可以进行到末尾; (c) 后 24 个碱基为部分错配时的换链曲线, 红色为界面位置; (d) 33% 错配时的换链长度, 红色实线代表界面位置, 红色虚线为界面前9 bp的位置Fig. 2. Using magnetic tweezers to measure the strand exchange length under different periodic mismatch sequence: (a) From left to right are diagrams of experiments when strand exchange is not happened, strand exchange process under homologous sequence and periodic mismatch sequence, respectively. Red strands represent mismatch sequence and blue strands represent homologous sequence;(b) trace of fully homologous sequence; (c) trace of periodic mismatch sequence; (d) strand exchange length distributions of 33%periodic mismatch sequence, red line marks the interface and red-dashed line marks the position which is 9 bp before the interface.
上述现象说明核蛋白丝链交换的步长倾向于以 9 bp 发生而不是 3 bp, 与 Zhang 等[13]的实验结果相符合. 结合同源比对过程是以9 bp进行的理论结果, 推测在同源比对完成后, 紧接着发生了同样长度的链交换. 至于界面到–9 bp之间的序列,完全匹配但是为什么仍然不进行链交换, 还需更多的实验来探究, 这可能涉及到同源识别和链交换更深层次的分子机理.
3.3 连续错配碱基对换链的影响
前面的错配实验提示链交换步长倾向为9 bp而不是3 bp, 将错配碱基由周期性均匀分布调整为局域连续分布. 实验使用全长69个碱基的同源序列, 从第40号碱基开始, 插入3个连续错配碱基, 通过磁镊方法判断链交换能否跨过不同长度的连续错配碱基.
实验模型图如图3(a)所示, 蓝色部分为同源序列, 红色部分为连续错配碱基序列, 通过测量磁球高度的变化, 观察链交换反应能否跨过不同长度的局域错配碱基. 为了更直观地表现错配碱基对链交换的影响, 使用柱状图统计了不同错配碱基数时链交换能够跨过界面位置的比例, 结果如图3(b)所示, 同样的反应条件和浓度下, 假设全匹配的链交换事件发生的概率是100%, 所有的链交换行为可跨过2个连续错配, 完全发生链交换反应; 7成的链交换行为可以跨过连续3个错配碱基; 83%的链交换会被4个连续碱基阻挡, 停在界面之前. 当连续错配碱基数目为5个时, 链交换反应则完全无法跨过错配碱基.
连续3个错配碱基的链交换结果进一步说明了链交换的步长并非3 bp, 如果链交换的步长是3 bp,是没有理由跨过100%错配的triplet的. 同时, 如图 3(c)所示, 当链交换步长为 9 bp 时, 3 个错配碱基的均匀分布和连续分布在9 bp内的错配程度同样为33%, 却表现了不一样的结果. 对比二者的错配碱基分布, 33%错配序列中每3个碱基中有一个错配碱基, 界面后的每个triplet均受到影响, 而3个错配碱基影响的triplet数量有限, 所受影响的triplet数量对链交换有着重要影响.
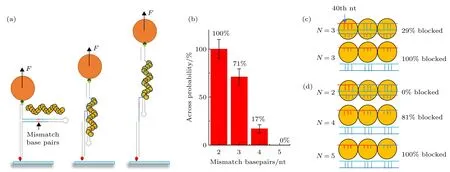
图 3 使用磁镊方法探究换链是否能够跨过连续错配碱基 (a) 磁镊实验的示意图, 红色线段位置为错配碱基序列, 蓝色线段为同源碱基序列, 从左到右分别是链交换反应之前, 链交换被错配碱基阻碍和链交换跨过错配序列三种现象; (b) 不同连续错配碱基数目时, 越过错配碱基事件数量的百分比(误差线代表统计误差); (c) 33%错配序列和连续3 bp错配序列的碱基位置分布,N为错配碱基数目, 红色代表错配碱基, 蓝色代表同源碱基; (d)同(c)一样, 用来表示N = 2, 4和5时的错配碱基分布Fig. 3. Using magnetic tweezers to investigate if strand exchange could stride across continuous mismatch base pairs: (a) From left to right are diagrams of experiments when strand exchange is not happened, strand exchange process blocked by continuous mismatch base pairs and strand exchange process stride across continuous mismatch base pairs, respectively. Red segments represent mismatch base pairs and blue segments represent homologous base pairs; (b) across probability when facing different continuous sequence; (c) the mismatch base pairs distribution of 33% periodic mismatch sequence and 3 bp continuous mismatch sequence, N represents the number of mismatch base pairs; (d) same as (c) when N = 2, 4 and 5.
如图3(d)所示, 随着连续错配碱基数量的增加, 9 bp范围内序列的同源度逐渐降低, 跨过错配碱基的链交换事件减少, 较低的同源度给同源重组过程增加了阻力. 除此之外, 较多的连续错配碱基也使受错配碱基影响的triplet数量增加, 在低同源度和受影响triplet数量的共同作用下, 5个连续错配碱基彻底使同源重组反应中止. 实验说明序列同源度和受影响的triplet数量均会对链交换产生影响, 但它们影响的具体机制和各自的影响效果还需要实验进一步探索.
4 讨 论
本文结合了磁镊和酶切保护的方法, 通过不同错配序列的链交换实验, 验证了链交换的步长为9 bp的结论. 结合理论与实验结果, 我们认为在同源序列比对过程中, 同时检测9 bp序列同源度时的自由能最低[26], 与链交换步长倾向于以9 bp发生存在关联. DNA在与核蛋白丝进行同源比对时,被置换链与第二结合位点结合, 第二结合位点提供的结合能, 使被置换链与互补链均能被剧烈拉伸,但该体系并非处在一个稳定的状态, 趋向于发生链交换形成自由能更低的状态. 因此在同源比对成功后会紧接着发生链交换反应, 链交换的长度与同源比对长度相等, 均为 9 bp. 同时, 通过这些实验还发现了均匀分布的错配序列把链交换的终点挡在了界面之前9 bp, 这个现象可能与链交换的同源识别机制密切相关, 但还需进一步的实验来探索.我们还发现错配碱基的数量和分布都会对链交换产生影响, 简单来说就是错配程度越高, 错配涉及的triplet数目越多, 越难发生链交换. 我们推测,错配碱基数的增加使发生同源序列比对的9 bp内碱基的同源度不断降低, 互补链与入侵链形成杂合双链所降低的自由能无法弥补因含有错配碱基的两条链形成杂合双链产生的不稳定性, 使链交换无法自发进行. 相比于换链步长为3 bp的理论, 换链步长为9 bp的同源检测机制可以容纳更多的错配碱基, 也能蕴含更多的调控手段, 使同源重组过程具有更灵活的可控性, 从而使其对生物生存的有利程度最大化. 至于同源比对是如何整体考虑9 bp内的错配数量和分布, 从而决定是否换链, 其更深层次和更细致的机理, 可能会十分复杂, 还需设计更多的实验来得到确切的结论. 至于其他人3 bp链交换步长的研究结果, 也不能说是完全错误的.我们认为3 bp应该是基本的结构单位, 而9 bp是主要的功能单位, 亦或是有更深层的分子机制可以将3 bp与9 bp的结果统一起来, 这些都需要进一步的实验去证实.
在实验中使用酶切保护实验验证链交换反应是否完全进行, 使用了磁镊观测链交换碱基数的方法, 并通过改变碱基序列进一步发掘了同源重组的链交换机制. 酶切保护实验看到的现象有限, 但是实验数据量多, 适合对猜想进行提前验证; 磁镊实验数据量少但观测的现象丰富, 可以作为实验的主要研究方法. 两种手段相辅相成, 二者亦可结合起来, 拓宽实验设计的范围, 得到新的信息, 进一步推动同源重组过程的研究.
5 结 论
本文使用酶切保护实验证明了同源重组链交换的步长可能不为3 bp, 用单分子磁镊方法, 通过周期性错配序列, 验证了同源重组的链交换步长倾向为9 bp. 通过连续错配序列, 也倾向于否定步长为3 bp的说法, 同时还发现了9 bp内序列的同源度和错配碱基的分布共同对链交换造成影响. 得到以上结论的同时, 我们也得到很多启示, 提示同源重组分子机理的复杂性. 这些尚不能解释的现象促使我们设计更多的实验来一步步深入探索, 逐步解开同源重组分子机制这一大难题.
