条斑紫菜丝状体不同发育时期对光照和温度的光合适应能力*
2020-11-03魏家慧李国梁汪文俊梁洲瑞鲁晓萍刘福利孙修涛张朋艳
魏家慧 李国梁 汪文俊 梁洲瑞 鲁晓萍 刘福利 孙修涛 张朋艳
(1. 中国水产科学研究院黄海水产研究所 农业农村部海洋渔业可持续发展重点实验室 青岛 266071;2. 上海海洋大学水产与生命学院 上海 201306;3. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071)
条斑紫菜(Pyropia yezoensis)是一种较为原始的 大型红藻,主要分布在我国浙江舟山群岛以北的东海和黄、渤海沿岸及朝鲜半岛和日本沿海等地(许璞等,2013)。条斑紫菜具有很高的营养、经济和生态价值,是我国重要的大型栽培海藻(李晓蕾等, 2017)。条斑紫菜生活史包括宏观配子体(叶状体)和微观孢子体(丝状体) 2 个异形世代(马家海等, 1996)。丝状体分为丝状藻丝(营养藻丝)、孢子囊枝、壳孢子形成(张学成等, 2005) 3 个阶段。环境适宜时,成熟的孢子囊枝放散壳孢子,继而发育生成新的叶状体。条斑紫菜叶状体生长于潮间带,藻体每天随潮汐的变化,经受温度、光照、盐度及水分等环境因子的胁迫(Davison et al,2010),是研究植物抗逆性及其机理的良好实验材料。
目前,有关条斑紫菜的研究主要集中在适宜丝状体生长发育的环境生态条件(郭文竹, 2012; 张美如等,2009)、叶状体抗逆性及机制(侯和胜等, 2008; 周向红等, 2013)与分子生物学等(贺剑云等, 2010; Yang et al,2011; Cao et al, 2018)。丝状体是紫菜生活史的重要阶段,作为种质资源保存形式,调控其生长发育对生产和科研十分重要。孢子囊枝和营养藻丝是紫菜丝状体发育最典型的2 个阶段,受环境因子影响显著(Green et al, 2015)。高温促进丝状体由营养藻丝发育为孢子囊枝。研究表明,温度达到12.5℃以上,才能形成孢子囊枝(任国忠等, 1979);15℃~25℃之间,壳孢子囊枝的增长率与温度成正比(陈翠芬等, 2008)。光是植物进行光合作用的能量来源,对植物体生长、发育具有重要影响(卢晓等, 2014; 夏云捷等, 2016)。光强过高或过低均不利于紫菜丝状体的生长和发育。骆其君等(1999)研究表明,15℃、60 μmol/(m2·s)是条斑紫菜丝状体适宜的温度和光照。陈翠芬等(2008)研究发现,8 L ∶16 D的短日照光周期、57 μmol/(m2·s)光强下,壳孢子囊枝形成率最高。Zhong 等(2016)对长紫菜(Pyropia dentata)丝状体发育研究表明,较高温度有利于孢子囊枝的形成;在不高于12 h 日照情况下,孢子囊枝形成率随日照时间的延长而增加;在一定的光照范围内,孢子囊枝的形成随光强的增加而增加。
紫菜发育和有性生殖的调控与环境紧密关联,而这个过程藻体光合作用也会发生相应变化(Katz et al,2000; Jiang et al, 2013)。紫菜丝状体发育阶段,营养藻丝和孢子囊枝在形态上存在巨大差异(Shen et al,2000; Varela-Alvarez et al, 2004; 周文君等, 2006),对环境因子需求也有所不同,但二者在不同环境条件下的光合生理差异尚未明确。本研究通过叶绿素荧光技术和液相氧电极技术,对条斑紫菜自由丝状体营养藻丝与孢子囊枝阶段在不同温度与光强下的光合生理进行比较研究,以比较二者光温适应能力,尤其是对高温和高光胁迫的光合耐受性,旨在为揭示条斑紫菜丝状体发育光温调控机制、建立苗种繁育精准调控技术奠定基础。
1 材料与方法
1.1 实验材料
采用课题组培育的条斑紫菜“黄优1 号”品系为实验材料。营养藻丝和孢子囊枝暂养条件:20℃,光强30 μmol/(m2·s),光周期12 L ∶12 D,灭菌海水中添加营养盐(5 mg/L-N,1 mg/L-P)为培养液。培养适应3 d 后进行实验。
1.2 实验方法
1.2.1 温度和光强对丝状体Fv/Fm的影响 实验于DGXM-508B 光照培养箱中进行,称取等量营养藻丝和孢子囊枝分别放于培养皿中。设置3 个温度(15℃、25℃和35℃)和3 个光强[40、100 和300 μmol/(m2·s)]。分别测定不同光温组合培养6 h、1 d、2 d 和3 d 的Fv/Fm。Fv/Fm测定采用叶绿素荧光仪(Imaging-Pam,WALZ, 德国),方法:取少量藻体于暗盒中,分别进行20 min 暗适应,每个样品取6 个平行,测定其Fv/Fm。
1.2.2 温度和光强对丝状体Pg、Rd和Pn的影响实验设置同 1.2.1,采用液相氧电极(Hansatech Oxygraph, 英国)测定,反应介质为灭菌海水,光源为500 W 碘钨灯,通过控制灯的远近,将光强控制在40 μmol/(m2·s)。每个条件下反应稳定5 min,测试时间控制在10 min 内,取较为平滑曲线计算。由氧电极自带软件得出数据,每个处理组设置3 个平行。
1.3 数据处理
采用Excel 分析数据并绘图,采用SPSS 18 软件对各组数据进行单因子方差分析(One-way ANOVA)及Duncan 多重比较,P<0.05 为差异显著。
2 结果
2.1 温度和光强对营养藻丝和孢子囊枝 Fv/Fm 的影响
15℃时,除40 μmol/(m2·s)光强组,其他实验组孢子囊枝的Fv/Fm均显著高于营养藻丝(P<0.05),此现象随光强增加而越明显,且二者Fv/Fm均随光强增加而下降。光强为40 μmol/(m2·s)时,6 h~2 d 内,孢子囊枝与营养藻丝的Fv/Fm均呈上升趋势,3 d 时显著下降(P<0.05),2 种藻丝的Fv/Fm在最优时相当。100 μmol/(m2·s)光照下,孢子囊枝的Fv/Fm呈线性下降趋势;当光强达到300 μmol/(m2·s)时,第3 天时,孢子囊枝的Fv/Fm显著降低(P<0.05);在光强为100~300 μmol/(m2·s)时,营养藻丝的Fv/Fm一直处在非常低的水平(P<0.01)。

图1 15℃下,不同光强对条斑紫菜丝状体Fv/Fm 的影响Fig.1 Fv/Fm of P. yezoensis under different light intensities at 15℃
温度为25℃时,孢子囊枝的Fv/Fm均显著高于营养藻丝(P<0.05),但不同光强下情况不同:光强为40与100 μmol/(m2·s)组情况相同,营养藻丝Fv/Fm变化不大,孢子囊枝的Fv/Fm先下降(P<0.05),后有所恢复。当光强达到300 μmol/(m2·s)时,营养藻丝Fv/Fm趋近于零,藻体发绿变白,接近死亡;孢子囊枝Fv/Fm先上升后有所下降(P<0.05);但二者Fv/Fm均显著低于40~100 μmol/(m2·s)组(P<0.05)。
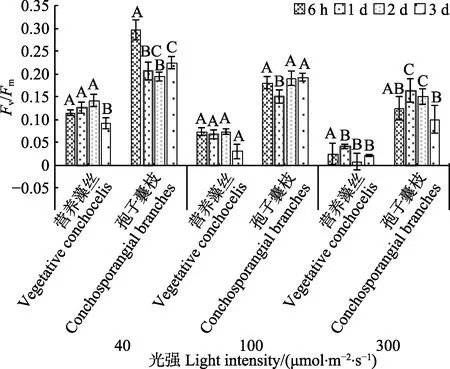
图2 25℃不同光强对条斑紫菜丝状体Fv/Fm 的影响Fig.2 Fv/Fm of P. yezoensis under different light intensities at 25℃
35℃时,营养藻丝与孢子囊枝的Fv/Fm显著低于15℃和25℃组。不同光强下,营养藻丝的Fv/Fm几乎检测不到(P>0.05)。40~100 μmol/(m2·s)光强下,孢子囊枝在培养初期(6 h),Fv/Fm随光强增加而急剧下降;当光强为300 μmol/(m2·s)时,孢子囊枝在6 h 的Fv/Fm几乎为0。营养藻丝在6 h 时,藻体变绿发白,孢子囊枝在1~2 d 后,藻体也变绿发白、死亡。
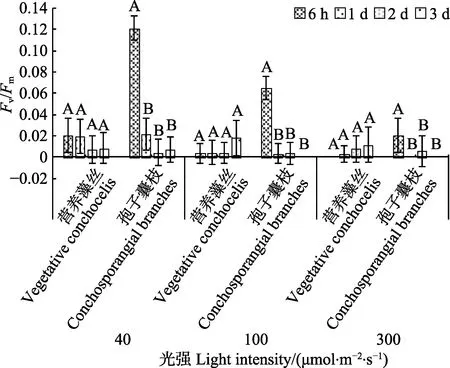
图3 35℃不同光强对条斑紫菜丝状体Fv/Fm 的影响Fig.3 Fv/Fm of P. yezoensis under different light intensities at 35℃
2.2 温度和光强对丝状体总光合速率的影响
15℃时,孢子囊枝的总光合速率均显著高于营养藻丝(P<0.05)。随光强的增加,二者Pg逐渐降低。40 和100 μmol/(m2·s)光照下,孢子囊枝Pg呈递增趋势,营养藻丝则呈先上升后下降的趋势(P<0.05)。300 μmol/(m2·s)时,第2 天,营养藻丝出现显著上升后又急剧下降,其余各组间无显著差异(P>0.05)。

图4 15℃不同光强对条斑紫菜丝状体总光合速率的影响Fig.4 Gross photosynthesis rate of P. yezoensis under different light intensities at 15℃
当温度为25℃、光强为40 μmol/(m2·s)时,孢子囊枝Pg随培养时间延长而线性上升(P<0.05),营养藻丝各组间无显著差异。3 个组变化情况与15℃大体一致,且孢子囊枝Pg显著高于营养藻丝(P<0.05)。
当温度为35℃、光强为40 μmol/(m2·s)时,培养1 d 孢子囊枝的Pg上升显著(P<0.05);第2 天,营养藻丝Pg降为负值(P<0.05),孢子囊枝Pg几乎为0(P<0.05);第3 天,营养藻丝恢复到6 h 水平。光强为100 μmol/(m2·s)时,2 种藻丝的总体变化趋势和40 μmol/(m2·s)时大体一致。300 μmol/(m2·s)胁迫下,6 h~2 d 中,营养藻丝和孢子囊枝的Pg在0 上下波动,变化趋势不显著;第3 天时,营养藻丝的Pg上升显著(P<0.05)。

图5 25℃不同光强对条斑紫菜丝状体总光合速率(Pg)的影响Fig.5 Gross photosynthesis rate of P. yezoensis under different light intensities at 25℃

图6 35℃不同光强对条斑紫菜丝状体总光合速率(Pg)的影响Fig.6 Gross photosynthesis rate of P. yezoensis under different light intensities at 35℃
2.3 温度和光强对丝状体呼吸耗氧速率的影响
温度为15℃时,光强为40 μmol/(m2·s)时,1 d时孢子囊枝与营养藻丝的Rd均显著上升(P<0.05)随后维持较为稳定的水平。当光强达到100 μmol/(m2·s)时,6 h 时营养藻丝的Rd显著高于孢子囊枝(P<0.05),随胁迫时间延长,孢子囊枝Rd逐渐升高,且显著高于营养藻丝(P<0.05)。光强300 μmol/(m2·s)时,实验前期,营养藻丝和孢子囊枝Rd呈上升趋势,且二者之间无显著差异(P>0.05)。
温度达到25℃时,光照为40 μmol/(m2·s)时,营养藻丝的Rd均显著高于孢子囊枝(P<0.05),且二者均呈上升趋势,此现象在孢子囊枝中尤为显著。光照为100 μmol/(m2·s)时,营养藻丝和孢子囊枝的Rd均呈先上升后下降趋势(P<0.05)。300 μmol/(m2·s)光强胁迫下,6 h 时,营养藻丝Rd显著高于孢子囊枝(P<0.05),6 h~2 d 时,孢子囊枝Rd上升(P<0.05),其余时间二者均无显著差异。
在35℃高温、40 μmol/(m2·s)光照胁迫下,营养藻丝和孢子囊枝的Rd波动较大,呈先上升后下降再上升的趋势,第1 天时,孢子囊枝Rd显著高于营养藻丝(P<0.05)。100 μmol/(m2·s)光强中,6 h~1 d 中,二者Rd呈显著上升(P<0.05),孢子囊枝的Rd显著高于营养藻丝。光强达到300 μmol/(m2·s)时,二者Rd与前2 组光照实验相比均有明显下降(P<0.05),实验过程中,营养藻丝与孢子囊枝Rd均呈上升趋势,且实验后期营养藻丝Rd显著高于孢子囊枝(P<0.05)。

图7 15℃不同光强对条斑紫菜丝状体呼吸耗氧速率(Rd)的影响Fig.7 Respiratory oxygen consumption rate of P. yezoensis under different light intensities at 15℃
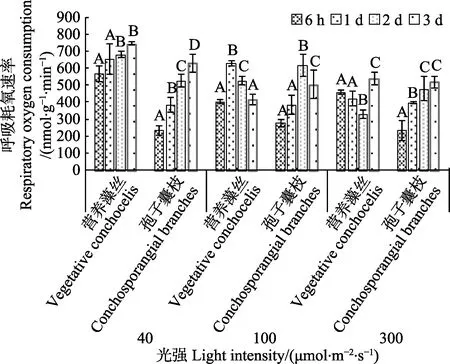
图8 25℃不同光强对条斑紫菜丝状体呼吸耗氧速率(Rd)的影响Fig.8 Respiratory oxygen consumption rate of P. yezoensis under different light intensities at 25℃
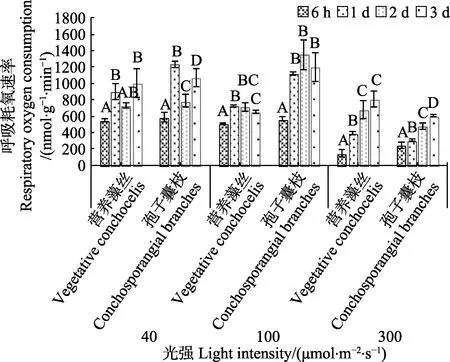
图9 35℃不同光强对条斑紫菜丝状体呼吸耗氧速率(Rd)的影响Fig.9 Respiratory oxygen consumption rate of P. yezoensis under different light intensities at 35℃
2.4 温度和光强对丝状体净光合速率的影响
营养藻丝的净光合速率(Pn)均为负值。孢子囊枝在15℃和25℃时Pn为正值,且随光强增加而下降;当温度达到35℃时,Pn急剧降低为负(表1)。
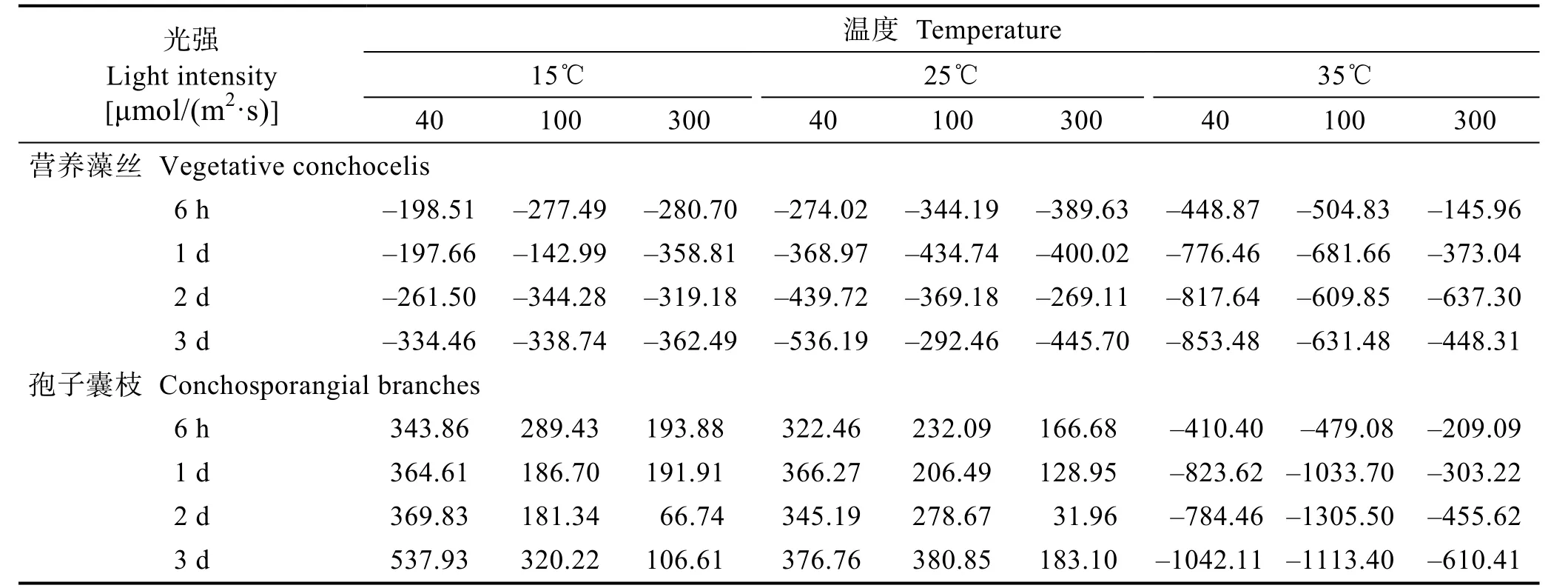
表1 温度、光照对条斑紫菜营养藻丝和孢子囊枝净光合速率(Pn)的影响Tab.1 The net photosynthetic rate of vegetative conchocelis and conchosporangial branches under different light intensities and temperatures
3 讨论
藻类进行光化学反应和电子传递的成分位于叶绿体类囊体膜上(Chow et al, 2005),叶绿素荧光参数可以反映PSⅡ的活性以及对光能吸收、传递、分配、耗散等内在性的联系(Maxwell et al, 2000)。当藻类在逆境条件下产生光抑制时,叶绿素各荧光参数均呈下降趋势(Zhou et al, 2004)。PSⅡ原初光能转化效率Fv/Fm反映植物在充足的暗适应后,所有PSⅡ反映中心均处于开放状态时的量子产量,是衡量植物光合系统完整性的指标之一(张守仁, 1999)。当植物遭遇外界胁迫时,Fv/Fm值存在不同程度的下降,则表明PSⅡ反应中心受到不同程度的损伤。总光合速率(Pg)表示在光照条件下,单位时间内(叶绿体)所合成的有机物的量或同化CO2的量,表观光合速率(Pn)表示单位时间内植物所积累的有机物含量或者从外界环境所吸收CO2量(陈卫东, 2018),由总光合速率(Pg)减去呼吸耗氧速率(Rd)所得。温度变化会影响藻细胞新陈代谢、种群增长和叶绿素含量(梁英等, 2009)。温度过高或过低均会使藻体生长缓慢,甚至死亡(张美如等,2009)。低温可以使植物类囊体膜上PSⅡ反应中心功能受到抑制(Terashima et al, 1989),降低PSⅡ的修复能力(Ottander et al, 1993);同时,PSⅡ也是对高温伤害最敏感的结构(Berry et al, 1980)。光是植物进行光合作用的能量来源,适宜的光强是保证植物正常生长发育的前提,但光强过高,则会对植物光合作用产生光抑制(郑宝福等, 1980; 陈国宜等, 1984; Zhang et al,1997)。植物适应或抵抗外界胁迫过程需要消耗大量能量,呼吸作用为植物生命活动提供大部分能量,通过增强呼吸作用来抵抗胁迫是植物抗逆途径之一。
温度和光照是影响紫菜丝状体生长发育的2 个最基本环境要素。通常在一定的光照和温度范围内,较高的温度、中短日长和较强的光照促进丝状体由营养藻丝发育为孢子囊枝(任国忠等, 1979; 骆其君等,1999; 陈翠芬等, 2008; 卢晓等, 2014; 夏云捷等, 2016;Zhong et al, 2016),但有关调控机制尚不清楚。实践工作中,营养藻丝在适宜的情况下,只要培养条件不变,可以长久维持营养生长,而不会向孢子囊枝转化。那么,光温的升高是否因不利于营养藻丝的生长,从而启动相关内源信号调控藻丝转向另一个发育阶段?为了验证这一假设,本研究设置温度和光照强度较大,包括适宜范围与极端胁迫。结果显示,在适宜温度范围内(15℃~25℃),除了温度为15℃、光强为40 μmol/(m2·s)时,二者在多数时间点的Fv/Fm相当,其他条件下,孢子囊枝的Fv/Fm、Pg与Pn均高于营养藻丝(P<0.05)。营养藻丝在25℃时,Fv/Fm、Pg与Pn值均低于15℃时,而孢子囊枝在25℃时,以上3 个指标均高于或相当于15℃时,说明孢子囊枝比营养藻丝适于更高的温度。这从Rd的变化也可以看出,2 种藻丝在15℃时,Rd差距不显著,在温度为25℃、光强为40 μmol/(m2·s)时,6 h~1 d 时,营养藻丝显著高于孢子囊枝。光合作用降低、呼吸作用增强是植物遭遇胁迫时的常见反应。因此,推测营养藻丝在温度为25℃、光照强度为40 μmol/(m2·s)时已受到胁迫。研究表明,35℃对紫菜丝状体已是致死高温胁迫,2 种藻丝的Fv/Fm趋于0,出现藻体死亡现象。2 种藻丝的以上光合生理指标基本上都随实验光强的增加而下降。说明过高的光强可以显著降低丝状体PSⅡ的光能利用效率。温度为15℃、光强为40 μmol/(m2·s)时,营养藻丝和孢子囊枝在多数时间点具有相近的Fv/Fm值,相同温度下,光强增加至100 μmol/(m2·s),孢子囊枝Fv/Fm值则显著高出营养藻丝;在温度为15℃~25℃、光照为300 μmol/(m2·s)时,虽然孢子囊枝的光合生理指标数值相比温度为15℃~25℃、光强为40~100 μmol/(m2·s)时有所下降,但还维持在较高水平,而营养藻丝的数值已降至0 附近,可见孢子囊枝对高光的耐受性显著高于营养藻丝。
孢子囊枝相对营养藻丝具备适应更高温度和光照的能力,可能与其在结构和成分上的改变有关。超微结构显示,首先,孢子囊枝中央液泡形成并增大(沈颂东等, 2000),液泡在多种逆境条件下起积极的渗透调节作用;第二,线粒体体积增大,数量变多(沈颂东等, 2000)。线粒体是进行呼吸代谢和能量转换的主要场所,细胞生命活动所需的能量大约有95%来自线粒体(Chance et al, 1979),条斑紫菜孢子囊枝中线粒体增多可能为机体抵抗外界胁迫提供了能量保障;第三,色素体被挤压分散排列在细胞边缘,并由大量的线粒体和淀粉粒分割开来(周文君等, 2006),极大提高了光能的吸收利用效率和有机物合成(光合碳固定);第四,细胞壁增厚疏松,外胶层变薄,内纤维质层极度加厚,细胞外壁凹凸不平,形成脊突,并随孢子囊枝日益成熟数量明显增加(朱家彦等,1984)。植物细胞壁增厚是抵御逆境胁迫的有效途径之一(庄惠如等, 2001)。另外,前期研究表明,在紫菜丝状体发育过程中,磷脂酰胆碱(PC)和溶血磷脂酰胆碱(LPC)含量增加(Wang et al, 2015)。PC 对PSⅡ膜复合物有保护作用(李冬海等, 2003),可以增强其放氧速率(Gounaris et al, 1983),同时,还可诱导Triton处理的光系统膜复合物的再活化(Ruan et al, 2002)。PC 脂酰基侧链的不饱和程度对调节细胞的高温/低温适应能力具有重要影响。LPC 是植物抗逆过程中重要的信号传递物质(卢静婵等, 2012)。同时,孢子囊枝中藻胆蛋白含量及其与叶绿素a 含量的比值等均有所上升(陈烨等, 2007)。藻胆蛋白为红藻主要捕光色素(姚南瑜, 1987),直接参与光合作用能量吸收和传递(Glazer, 1984),同时,还具有抗逆功能(Zhong et al,2016)。孢子囊枝藻胆蛋白含量上升,可增加光能捕获效率,促进光反应和碳水化合物合成。研究发现,在坛紫菜丝状体由营养藻丝发育到孢子囊枝过程,碳水化合物和蛋白质比值明显上升(陈烨等, 2007)。碳水化合物是细胞结构的主要成分和主要供能物质,碳水化合物的含量及其与蛋白质的比值是植物响应胁迫的重要指标,如,鹰嘴豆(Cicer arietinum)在干旱胁迫下,水溶性碳水化合物的含量显著升高(Mafakheri et al, 2011);细基江蓠(Gracilaria tenuistipitata var.
Liui)碳水化合物和蛋白质比值随温度的降低和光照的减弱下降(刘静雯等, 2001)。紫菜孢子囊枝细胞内这些结构和组成的改变,为孢子囊枝应对高温和高光胁迫提供了支撑。
综上所述,在条斑紫菜营养藻丝的光合作用被严重抑制的光强、温度条件下,孢子囊枝仍然具备相对高的光合活力,说明温度和光强升高到不利于营养藻丝生长的情况下,刺激藻丝转向孢子囊枝发育阶段,后者同时具备适应更高温度和光照的能力。育苗实践中,可利用二者对高温/高光耐受阈值的差异来调控丝状体的生长和发育。
