叉角厉蝽成虫对粘虫幼虫的捕食功能反应
2020-11-02廖贤斌高平赵航谢永辉陈雅琼王志江陈斌吴国星高熹
廖贤斌 高平 赵航 谢永辉 陈雅琼 王志江 陈斌 吴国星 高熹


摘要:【目的】探明室內叉角厉蝽对粘虫幼虫的捕食潜力,为叉角厉蝽应用于田间防治粘虫提供理论依据。【方法】以透明塑料盒为测试容器,在室内设不同密度(3、6、9、12和15头/盒)的3、4和5龄粘虫幼虫供叉角厉蝽成虫(1头/盒)取食,统计叉角厉蝽成虫的捕食量并进行相关分析。固定粘虫幼虫密度(50头/盒),设不同密度(1、2、3、4和5头/盒)的叉角厉蝽成虫,研究捕食者自身密度对捕食量的影响。等比例(天敌∶猎物=1∶10)增加叉角厉蝽成虫和粘虫幼虫,研究干扰反应对捕食量的影响。【结果】叉角厉蝽成虫对3、4和5龄粘虫幼虫的捕食功能反应均符合Holling Ⅱ模型。在室内条件下,叉角厉蝽成虫对4和5龄粘虫幼虫的理论最大取食量分别为10.09和3.55头,明显低于对3龄粘虫幼虫的理论最大取食量(78.74头)。叉角厉蝽成虫对3、4和5龄粘虫幼虫的搜寻效应方程分别为S=0.9009/(1+0.0114N)、S=0.7291/(1+0.0723N)和S=0.9322/(1+0.2626N),在相同虫龄内,搜寻效应随着猎物密度的增加而减小。捕食者密度对搜寻效应的影响及干扰反应的拟合结果分别为E=0.4254P-0.551和A=8.3413P-0.127,随着捕食者密度的增加,叉角厉蝽的平均捕食量都会逐渐减少,叉角厉蝽成虫与粘虫幼虫等比例增加时,叉角厉蝽也会产生种间干扰。【结论】叉角厉蝽成虫对3龄粘虫幼虫具有较好的捕食潜力,因此选择粘虫幼虫3龄时释放叉角厉蝽成虫为宜,但在田间实际释放时需考虑叉角厉蝽自身密度及种间干扰对其捕食能力的影响。
关键词: 叉角厉蝽;粘虫;捕食功能;干扰反应
中图分类号: S476.2;S433.4 文献标志码: A 文章编号:2095-1191(2020)08-1992-06
Predatory function response of adult of Eocanthecona furcellata(Wolff) to the larvae of Mythimna seperata(Walker)
LIAO Xian-bin1, GAO Ping1, ZHAO Hang1, XIE Yong-hui2, CHEN Ya-qiong2,
WANG Zhi-jiang2, CHEN Bin1, WU Guo-xing1, GAO Xi1*
(1College of Plant Protection, Yunnan Agricultural University, Kunming 650201, China; 2Kunming Branch of Yunnan Provincial Tobacco Company, Kunming 650021, China)
Abstract:【Objective】The predatory potential of Eocanthecona furcellata(Wolff) to the Mythimna seperata(Walker)larvae was explored, which provided a theoretical basis for control of E. furcellata to the M. perata larvae in field. 【Me-thod】With transparent plastic box as container, different densities of 3rd, 4thand 5th instar M. seperata larvae(3, 6, 6, 9, 12 and 15 ind/box) were set up indoors to feed on adults of E. furcellata(1 ind/box), and the predations of E. furcellata were counted and analyzed. The density of the M. perata larvae(50 ind/box) was fixed, and the adults of M. perata were set with different densities(1, 2, 3, 4 and 5 ind/box) to study the effect of the M. seperata density on the predation.Increasing the E. furcellata adult and M. perata larvae in equal proportions(natural enemy: prey = 1:10), and the effect of interference response to predation was studied. 【Result】The results showed that the predation function of the 3rd, 4th and 5th instar larvae of the M. seperata was consistent with the HollingII model.Under indoor conditions, the maximum theore-tical predation of E. furcellata adult on 4th and 5th larvae of the M. perata were 10.09 and 3.55, which were lower than those on the 3rd instar larvae(78.74). The search effect equations of E. furcellata adult on the 3rd, 4th, and 5th instar larvae of M. perata were S=0.9009/(1+0.0114N), S=0.7291/(1+0.0723N), S=0.9322/(1+0.2626N). During the same age period, the search effect decreased as the prey density increased. The effect of predator density on the search effect and the fitting results of the interference response were E=0.4254P-0.551 and A=8.3413P-0.127. With the increase of predator density, the amount, average predation of E. furcellata on the M. perata larvae gradually decreased. When E. furcellata adult and M. seperata larvae proportional increased, E. furcellata adult also produced interspecific interference. 【Conclusion】Studies have shown that E. furcellata adult has good predation potential for 3rd larvae M. seperata. When M. seperata larvae is 3rd instar, it has better control effect when releasing E. furcellata adult.But when E. furcellata is actually released in field,its own density and the impact of interspecific interference should be considered.
Key words: Eocanthecona furcellata (Wolff);Mythimna seperata(Walker); predatory function;interference response
Foundation item:National Key Research and Development Program(2018YFD0200703,2018YFD0200308); Science and Development Program of Yunnan Provincial Tobacco Company(2018530000241010)
0 引言
【研究意义】粘虫[Mythimna seperata (Walker)]隶属于鳞翅目(Lepidoptera)夜蛾科(Noctuidae),又称行军虫、剃枝虫,是一种世界性害虫。粘虫无滞育现象,条件适宜即可连续繁衍,因此随着发生区和气候的改变,粘虫每年发生的世代数和发生期也存在差异(韦秋凤,2012)。粘虫分布广泛,在我国除新疆外,各省份均有报道(江幸福等,2014);且食性广,除禾本科植物外,还取食为害林木、果树幼苗及豆、麻等作物,将作物的叶片吃光,或咬断麦穗、谷穗,造成减产甚至绝产,严重威胁我国农作物的生产安全(李光博,1996)。我国长期以来主要依赖化学农药防治粘虫,但农药的长期施用不仅破坏生态环境,影响天敌生存,还致使粘虫的抗药性明显增强,防治效果下降。生物防治措施因对生态环境和人体安全,且害虫不易产生抗药性等特点,已逐渐成为建设绿色无公害农业的重要研究内容。因此,研究粘虫生物防治措施对其长期绿色防控具有重要意义。【前人研究进展】捕食性天敌昆虫是害虫生物防治的重要组成部分,是最早被人类认识和利用进行害虫防治的自然天敌,也是目前生物防治研究的主要方向(林明江等,2012;池成林等,2018),在林业、设施园艺等害虫防控研究上已取得一定成效(杨怀文,2015;张帆等,2015;党英侨等,2018)。王玉杰等(2015)研究发现8和10龄广斧螳若虫对荔枝蝽若虫的日捕食量最高,具有作为有效天敌防治荔枝蝽的潜力;唐天成等(2018)发现中华通草蛉和大草蛉对黑刺粉虱均具有较好的捕食作用。叉角厉蝽[Eocanthecona furcellata(Wolff)]是一种捕食性天敌,其捕食范围广,能取食大多数鳞翅目昆虫(林长春等,1998)。目前,国内外已有较多关于叉角厉蝽的研究(Gupta et al.,2013;Tuan et al.,2016),针对其捕食能力的研究上也有相关报道。陈然等(2015)研究发现叉角厉蝽成虫对3龄斜纹夜蛾幼虫具有较好的捕食能力,不仅叉角厉蝽自生密度会对其捕食效应产生影响,叉角厉蝽种间干扰反应也会影响其捕食能力;李文华等(2015)研究叉角厉蝽对黄野螟的捕食功能反应时发现,高龄叉角厉蝽若虫对3龄黄野螟的捕食能力最强,在一定空间下叉角厉蝽自身密度会影响其捕食效应;叉角厉蝽对豆荚野螟、草地贪夜蛾和绿额翠尺蛾等害虫也具有一定的捕食潜力(谢钦铭等,2001;Pillai and Agnihotri,2013;唐敏等,2019)。叉角厉蝽还会影响害虫成虫的生殖行為。陈美云等(2014)研究叉角厉蝽成虫对斜纹夜蛾生殖行为的胁迫时发现,叉角厉蝽成虫能干扰斜纹夜蛾正常的产卵活动,改变斜纹夜蛾的卵块结构、大小和空间分布,减少斜纹夜蛾产卵量。【本研究切入点】目前,国内针对粘虫的生物防治研究较少,多集中于对粘虫天敌的调查,天敌捕食能力的研究仅唐艺婷等(2018)测定了益蝽对粘虫幼虫的捕食功能反应,而叉角厉蝽对粘虫幼虫的捕食能力研究未见报道。【拟解决的关键问题】通过研究叉角厉蝽对粘虫幼虫的捕食能力、寻找效应和干扰反应,确定叉角厉蝽成虫对3、4和5龄粘虫幼虫的捕食能力、叉角厉蝽成虫自身密度及种间干扰反应对其捕食能力的影响,进而明确室内条件下叉角厉蝽成虫对粘虫幼虫的捕食潜力,以期为叉角厉蝽应用于田间防治粘虫提供理论依据。
1 材料与方法
1. 1 试验材料
叉角厉蝽:采集自云南省元江县,在室内温度(28±1)℃、湿度(65±10)%、光照周期L∶D=14∶10环境下长期饲养的稳定种群,试验前叉角厉蝽成虫均饥饿24 h。粘虫:采集自云南省宣威市,在室内温度(25±1)℃、湿度(60±10)%、光照周期L∶D=14∶10环境下饲养的稳定种群,以3、4和5龄幼虫供试。
1. 2 叉角厉蝽对粘虫的捕食功能反应
以透明盒作为测试容器,测试容器为长14.0 cm、宽8.3 cm、高7.0 cm的塑料盒。粘虫幼虫密度分别设为3、6、9、12和15头/盒,共5个梯度水平。将相应虫龄及数量的粘虫幼虫移入测试容器中,加入适量玉米叶,在测试容器上挖数个洞用于通风,然后移入1头饥饿24 h的叉角厉蝽成虫,24 h后统计叉角厉蝽的捕食量。每处理5次重复。
1. 3 捕食者自身密度对功能反应的影响
每个测试容器中放入50头3龄粘虫幼虫及不同密度的叉角厉蝽成虫,叉角厉蝽的密度为1、2、3、4和5头/盒。24 h后观察叉角厉蝽的捕食量,每处理3次重复。
1. 4 干扰反应
每个测试容器按天敌∶猎物=1∶10,移入1头叉角厉蝽成虫、10头粘虫;2头叉角厉蝽成虫、20头粘虫;3头叉角厉蝽成虫、30头粘虫;4头叉角厉蝽成虫、40头粘虫;5头叉角厉蝽成虫、50头粘虫。24 h后观察叉角厉蝽捕食量,每处理3次重复。
1. 5 统计分析
叉角厉蝽对粘虫的功能反应方程:
式中,Ne为供试猎物密度,N为被捕食的猎物数量,T为试验持续时间,a为瞬时攻击率,Th为处置1头猎物的时间。因为T =1 d(24 h),以上公式简化为:
叉角厉蝽对粘虫搜寻效应方程:
式中,a为瞬时攻击率,Th为处置1头猎物的时间,N为被捕食的猎物数量,S为搜寻效应值。
Hassell模型:
式中,Q为搜寻常数,P为捕食者的密度,m为干扰常数,E为捕食作用率,Ne为捕食的猎物总数,N为猎物密度(Hassell,1969)。
Watt反应模型方程:A=aP-b。
式中,A为竞争条件下的捕食量;a为常数(无竞争条件下每头天敌的捕食量估计),P为天敌密度,b为种内竞争参数(Watt,1959)。
2 结果与分析
2. 1 叉角厉蝽成虫对粘虫的捕食功能反应
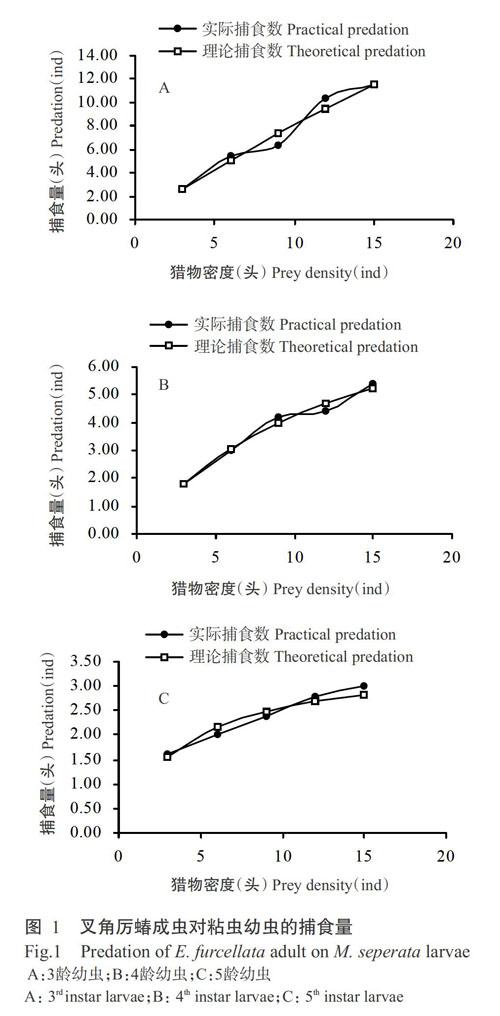
叉角厉蝽成虫对3、4和5龄粘虫幼虫的捕食量见图1。叉角厉蝽成虫对粘虫幼虫的捕食量随着粘虫幼虫密度的增大而增加,但其增加幅度随着粘虫密度增加而缓慢减小。叉角厉蝽成虫对3、4和5龄粘虫幼虫捕食功能反应的拟合结果见表1,其相关系数(r)为0.9219~0.9975,卡方检验(χ2)均小于0.0500,说明叉角厉蝽成虫对3、4和5龄粘虫的捕食功能反应均符合Holling Ⅱ模型。由表1可知,叉角厉蝽成虫对5龄粘虫幼蟲的瞬时攻击率(a)最高,为0.9322,对4龄粘虫幼虫的瞬时攻击率最低,为0.7291;随着粘虫虫龄的增大,叉角厉蝽成虫对粘虫的处理时间(Th)也越长,叉角厉蝽成虫对5龄粘虫的处理时间最长,为0.2817 d;叉角厉蝽成虫对3、4和5龄粘虫幼虫的理论最大捕食量分别为78.74、10.09和3.55头。
叉角厉蝽成虫在捕食粘虫幼虫的过程中,对于3龄粘虫幼虫,叉角厉蝽成虫一般会直接将口针插入其体内,此时粘虫虽有反抗,但反抗力度较小,叉角厉蝽的口针会一直插在粘虫体中,直至其死亡。对于虫体较大的4和5龄幼虫,当叉角厉蝽将口针插入虫体后,粘虫会出现剧烈抵抗,甚至会用口器攻击叉角厉蝽,部分叉角厉蝽会将口针收回,停顿在虫体周围片刻后,再寻找其他猎物;而有些叉角厉蝽不会将口针拔出,30~40 s后粘虫幼虫逐渐趋于平静,2~3 min后叉角厉蝽开始取食。
2. 2 叉角厉蝽成虫对粘虫幼虫的搜寻效应
叉角厉蝽成虫对粘虫幼虫的搜寻效果如图2所示,叉角厉蝽成虫对3、4和5龄粘虫幼虫的搜寻效应方程分别为S=0.9009/(1+0.0114N)、S=0.7291/(1+0.0723N)和S=0.9322/(1+0.2626N)。叉角厉蝽成虫对3龄粘虫幼虫的搜寻效应始终大于对4和5龄粘虫幼虫的搜寻效应。即当粘虫幼虫处于一定虫龄内,叉角厉蝽成虫对粘虫幼虫的搜寻效应随着猎物密度的增加而减小;粘虫幼虫密度一定时,叉角厉蝽成虫的搜寻效应随着粘虫幼虫虫龄的增大而减小。
2. 3 叉角厉蝽自身密度对粘虫搜寻效应的影响
根据方程E=QP-m(E=Ne/NP),叉角厉蝽成虫对3龄粘虫幼虫的拟合结果为E=0.4254P-0.551(r=0.944,χ2=0.0047)。由表2可知,当设3龄粘虫幼虫的密度为50头时,叉角厉蝽成虫的总捕食量与其自身密度呈正相关,而叉角厉蝽对粘虫的平均捕食量随着其自身密度的增加而减少、搜寻效应随着其自身密度的增加而减小。
2. 4 干扰反应
用Watt反应模型进行拟合,得到a=8.3413,b=0.127,拟合结果为A=8.3413P-0.127(r=0.9984,χ2=0.0003)。随着叉角厉蝽成虫数量的增加,叉角厉蝽的总捕食量增加,平均捕食量下降(表3)。
3 讨论
本研究结果表明,叉角厉蝽成虫对粘虫幼虫的捕食能力随着猎物密度的增加而上升,但上升幅度逐渐减小。叉角厉蝽对3、4和5龄期粘虫幼虫的捕食功能反应均符合Holling Ⅱ模型,且叉角厉蝽对3龄粘虫幼虫的理论最大捕食量为78.74头,说明叉角厉蝽对3龄粘虫具有较好的捕食能力。叉角厉蝽成虫对粘虫幼虫的最大捕食量排序为3龄>4龄>5龄,且对3龄粘虫幼虫的捕食量及搜寻效应均大于4和5龄粘虫幼虫。一方面,可能由于粘虫幼虫在3龄后进入暴食期,粘虫体型随着食量的增加而快速增长,高龄粘虫幼虫的体型明显大于中低龄的粘虫幼虫,使得在满足同等的营养需求下叉角厉蝽对高龄粘虫幼虫的捕食量远低于中低龄的粘虫幼虫;另一方面,高龄粘虫幼虫有较强的反抗能力,会干扰叉角厉蝽的取食,致使叉角厉蝽的取食效率降低。唐敏等(2019)在研究叉角厉蝽对草地贪夜蛾的捕食能力时也发现,草地贪夜蛾5龄幼虫在受到攻击时会表现出较强的防御能力,削弱了叉角厉蝽对草地贪夜蛾的处置效率。因此,在田间实际释放叉角厉蝽时,为取得较好的防治效果,应尽量选择在粘虫虫龄较低时进行。
巫鹏翔等(2016,2017)研究表明,在一定的空间下,异色瓢虫和七星瓢虫在捕食枸杞木虱时其自身密度和种间干扰均会影响捕食作用,本研究也有类似的结果。在一定的生存环境内,当猎物的密度固定时,虽然叉角厉蝽的总捕食量会随着自身密度的增加而上升,但平均捕食量及搜寻效应会随之逐步下降,说明叉角厉蝽自身密度的增加会产生干扰作用(m=0.556);在空间一定的条件下,叉角厉蝽与粘虫密度等比例增加时,叉角厉蝽成虫间表现出种内干扰(b=0.127)。因而在实际应用中需考虑叉角厉蝽自身密度和种间干扰对捕食能力的影响,实际释放前需尽量调查田间的粘虫虫态和数量,尽量选择在粘虫低龄期且大量繁衍时期释放,减少自身密度和种间干扰的影响。在玉米田间释放叉角厉蝽时,因叉角厉蝽不仅能取食粘虫,对草地贪夜蛾和斜纹夜蛾也具有捕食能力,在前期调查时还需掌握草地贪夜蛾和斜纹夜蛾的数量,以便在实际释放中能取得较好的防治效果。
本研究是在实验室封闭的条件下进行,猎物密度及环境因素均被控制在一定条件下,是在较理想状态下研究天敌昆虫的捕食能力。而在野外,除了空间及猎物资源外,光照、湿度、温度、其他天敌、害虫等天气及生物因素均会影响天敌昆虫的实际控害效果,在实际应用时,需根据建立的模型进行估算,以达到最佳的控害作用(Heinz and Nelson,1996;王媛等,2015)。因而在田间释放叉角厉蝽防治粘虫时,需考虑田间环境因素及田间实际的害虫数量来确定叉角厉蝽释放的时间和数量。
4 结论
叉角厉蝽成虫对3龄粘虫幼虫具有较好的捕食能力,因此选择在粘虫幼虫3龄时释放叉角厉蝽为宜,但在田间实际释放时需考虑自身密度及种间干扰对其捕食能力的影响。
参考文献:
陈美云,陈科伟,王兰兰,贾彩娟,冼继东. 2014. 叉角厉蝽对斜纹夜蛾生殖行为的干扰胁迫[J]. 环境昆虫学报,36(6): 884-889. [Chen M Y,Chen K W,Wang L L,Jia C J,Xian J D. 2014. Studies on the effect of nonconsumption stress of Eocanthecona furcellata(Wolff) on reproductive fitness of Spodoptera litura(Fabricius)[J]. Journal of Environmental Entomology,36(6): 884-889.]
陈然,梁广文,张拯研,曾嵘,冼继东. 2015. 叉角厉蝽对斜纹夜蛾的捕食功能反应[J]. 环境昆虫学报,37(2): 401-406. [Chen R,Liang G W,Zhang Z Y,Zeng R,Xian J D. 2015. The functional response of Cahtheconidea furcellata(Hemiptera: Asopinae) to Spodoptera litura(Lepidoptera:Noctuidae)[J]. Journal of Environmental Entomo-logy,37(2): 401-406.]
池成林,李強,马丽. 2018. 胡蜂亚科昆虫生物学特性及控害效果研究进展[J]. 南方农业学报,49(6):1139-1146. [Chi C L,Li Q,Ma L. 2018. Biological characteristics and controlling pest effects of Vespinae insect: A review[J]. Journal of Southern Agriculture,49(6):1139-1146.]
党英侨,王小艺,杨忠岐. 2018. 天敌昆虫在我国林业害虫生物防治上的研究进展[J]. 环境昆虫学报,40(2): 242-255. [Dang Y Q,Wang X Y,Yang Z Q. 2018. Advances in biological control of forest insect pests by using natural enemies in China[J]. Journal of Environmental Entomology,40(2): 242-255.]
江幸福,张蕾,程云霞,罗礼智. 2014. 我国粘虫研究现状及发展趋势[J]. 应用昆虫学报,51(4): 881-889.[Jiang X F,Zhang L,Cheng Y X,Luo L Z. 2014. Current status and trends in research on the oriental armyworm,Mythimna separata(Walker) in China[J]. Chinese Journal of Applied Entomology,51(4): 881-889.]
李光博. 1996. 中国农作物主要病虫害[M]. 北京: 中国农业出版社:657-723. [Li G B. 1996. Major crop pests and diseases in China[M]. Beijing: China Agriculture Press:657-723.]
李文华,贾彩娟,陈惠平,付浪,温健,陈科伟. 2015. 叉角厉蝽对黄野螟幼虫的捕食功能反应[J]. 环境昆虫学报,37(4): 843-848. [Li W H,Jia C J,Chen H P,Fu L,Wen J,Chen K W. 2015. Functional response of Eocanthecona furcellate(Wolff) to the larvae of Heortia vitessoides (Moore)[J]. Journal of Environmental Entomology,37(4): 843-848.]
林长春,王浩杰,任华东,洪长富. 1998. 叉角厉蝽生物学特征研究[J]. 林业科学研究,11(1): 89-93.[Lin C C,Wang H J,Ren H D,Hong C F. 1998. Studies on the Biological Characteristics of Cantheconidea furcellata(Wolff)[J]. Forest Research,11(1): 89-93.]
林明江,安玉兴,管楚雄,许汉亮,李继虎,梁瑞波. 2012. 甘蔗螟虫捕食性天敌昆虫利用前景[J]. 甘蔗糖业,(3): 29-33. [Lin M J,An Y X,Guan C X,Xu H L,Li J H,Liang R B. 2012. Application prospects of predator of sugarcane borers[J]. Sugarcane and Canesugar,(3): 29-33.]
唐敏,邝昭琅,李子园,陆永悦,陈科伟,刘光华. 2019. 叉角厉蝽对草地贪夜蛾幼虫的捕食功能反应[J]. 环境昆虫学报,41(5): 979-985. [Tang M,Kuang Z L,Li Z Y,Lu Y Y,Chen K W,Liu G H. 2019. Functional response of Eocanthecona furcellate(Wolff) to the larvae of Spodoptera frugiperda(J. E. Smith)[J]. Journal of Environmental Entomology,41(5): 979-985.]
唐天成,张艳,李程锦,曹欣然,陈珍珍,许永玉. 2018. 中华通草蛉和大草蛉幼虫对黑刺粉虱若虫的捕食功能反应[J]. 应用昆虫学报,55(2): 217-222. [Tang T C,Zhang Y,Li C J,Cao X R,Chen Z Z,Xu Y Y. 2018. Predatory responses of Chrysoperla sinica(Tjeder) and Chrysopa pallens larvae to Aleurocan spinfetus(Quaintance) nymphs. Chinese Journal of Applied Entomology,55(2):217-222.]
唐艺婷,郭义,何国玮,刘晨曦,陈红印,张礼生,王孟卿. 2018. 不同龄期的益蝽对粘虫的捕食功能反应[J]. 中国生物防治学报,34(6): 825-830. [Tang Y T,Guo Y,He G W,Liu C X,Chen H Y,Zhang L S,Wang M Q. 2018. Functional responses of Picromerus lewisi Scott(Hemiptera: Pentatomidae) attacking Mythimna separata(Wal-ker)(Lepidoptera: Noctuidae)[J]. Chinese Journal of Bio-logical Control,34(6):825-830.]
王媛,陈越,张拯研,曾嵘,冼继东. 2015. 不同龄期异色瓢虫对烟蚜的捕食潜能研究[J]. 环境昆虫学报,37(5): 1075-1080. [Wang Y,Chen Y,Zhang Z Y,Zeng R,Xian J D. 2015. Study on preying potential of different stages of nymphs and adults of Harmonia axyridis Pallas on Myzus persicae(Sulzer)[J]. Journal of Environmental Entomology,37(5): 1075-1080.]
王玉杰,吳娇,赵怡楠,黎荣欣,赵冬香. 2015. 广斧螳若虫对荔枝蝽若虫的捕食功能反应与搜寻效应[J]. 植物保护学报,42(3): 310-315. [Wang Y J,Wu J,Zhao Y N,Li R X,Zhao D X. 2015. Functional response and searching rate of giant Asian mantis Hierodula patellifera(Serville) nymphs to litchi stink bug Tessaratoma papillosa(Drury) nymphs[J]. Journal of Plant Protection,42(3):310-315.]
韦秋凤. 2012. 黏虫的生物学特性与防治技术初探[J]. 广西农学报,27(6): 32-34. [Wei Q F. 2012. Discussion on biological features and control technology of armyworm[J]. Journal of Guangxi Agriculture,27(6): 32-34.]
巫鹏翔,马宝旭,徐婧,何嘉,张蓉,张润志. 2017. 异色瓢虫成虫对枸杞木虱的捕食作用[J]. 植物保护学报,44(4): 582-588. [Wu P X,Ma B X,Xu J,He J,Zhang R,Zhang R Z. 2017. Predation of Poratrioza sinica Yang & Li by Harmonia axyridis adults[J]. Journal of Plant Prection,44(4): 582-588.]
巫鹏翔,欧阳浩永,徐婧,何嘉,张蓉,张润志. 2016. 七星瓢虫成虫对枸杞木虱的捕食作用[J]. 应用生态学报,27(11): 3712-3718. [Wu P X,Ouyang H Y,Xu J,He J,Zhang R,Zhang R Z. 2016. Predation of Poratrioza sinica Yang & Li by the adults of Coccinella septempunctata[J]. Chinese Journal of Applied Ecology,27(11): 3712-3718.]
谢钦铭,梁广文,罗诗,朱君林. 2001. 叉角厉蝽对绿额翠尺蛾幼虫的捕食作用的初步研究[J]. 江西科学,19(1): 21-23. [Xie Q M,Liang G W,Luo S,Zhu J L. 2001. Observation of the predacious function of a bug Cantheconidae furcellata on litchi looper Thalassodes proquadria[J]. Jiangxi Science,19(1): 21-23.]
杨怀文. 2015. 我国农业害虫天敌昆虫利用三十年回顾(上篇)[J]. 中国生物防治学报,31(5): 603-612. [Yang H W. 2015. Review in utilization of insect natural enemies during the period from 1985 to 2015 in China(Part 1)[J]. Chinese Journal of Biological Control,31(5): 603-612.]
張帆,李姝,肖达,赵静,王然,郭晓军,王甦. 2015. 中国设施蔬菜害虫天敌昆虫应用研究进展[J]. 中国农业科学,48(17): 182-195. [Zhang F,Li S,Xiao D,Zhao J,Wang R,Guo X J,Wang S. 2015. Progress in pest management by natural enemies in greenhouse vegetables in China[J]. Scientia Agricultura Sinica,48(17): 182-195.]
Gupta R K,Gani M,Arora R K. 2013. Prey infected with nucleopolyhedrovirus is preferred by the predatory stink bug Eocanthecona furcellata without altering its field fitness[J]. Biocontrol Science and Technology,23(12): 1442-1457.
Hassell M P. 1969. New induction population model for insect parasities and its bearing on biological control[J]. Nature,223: 1113-1137.
Heinz K M,Nelson J M. 1996. Interspecific interactions among natural enemies of Bemisia in an inundative biological control program[J]. Biological Control,6(3): 384-393.
Pillai A K,Agnihotri M. 2013. Biology and predatory potential of Eocanthecona furcellata(Wolff) on Maruca vitrata Geyer[J]. Madras Agricultural Journal,21(1):95-101.
Tuan S J,Yeh C C,Atlihan R,Chi H. 2016. Linking life table and predation rate for biological control:A comparative study of Eocanthecona furcellata(Hemiptera:Pentatomidae)fed on Spodoptera litura(Lepidoptera:Noctuidae) and Plutella xylostella(Lepidoptera: Plutellidae)[J]. Journal of Economic Entomology,109(1):13-24.
Watt K E F. 1959. A mathematical model for the effect of densities of attacked and attacking species on the number attack[J]. The Canadian Entomologist,91(3): 129-144.
(责任编辑 麻小燕)
收稿日期:2019-12-16
基金项目:国家重点研发计划项目(2018YFD0200703,2018YFD0200308);云南省烟草公司科技计划项目(2018530000241010)
作者简介:*为通讯作者,高熹(1980-),博士,副教授,主要从事昆虫生理与害虫防治研究工作,E-mail: chonchon@163.com。廖贤斌(1995-),研究方向为害虫生物防治,E-mail: 565936610@qq.com
