脂多糖对河南华溪蟹免疫能力的调节作用
2020-10-30周妍英王兰
周妍英 王兰

摘要:以河南华溪蟹(Sinopotamon henanense)血淋巴细胞为材料,不同濃度脂多糖(1.0、2.0、4.0和8.0μg/mL)分别刺激溪蟹12h、24h后,收集各处理组溪蟹血淋巴细胞进行试验。利用多功能酶标仪测定酸性磷酸酶(ac-id phosphatase , ACP )、 碱性鳞酸酶(alkaline phosphatase , AKP) 和 溶菌酶(lysozyme , LSZ) 等溶酶体酶的活性及酚氧化酶(Phenoloxidase,P0)的活性;采用中性红染色法测定溶酶体膜的稳定性;采用实时焚光定量PCR法检测纷氧化酶原(prophenoloxidase,proPO)、溶菌酶2个基因mRNA表达水平的变化。结果表明:与空白对照组相比,注射脂多糖后,溪蟹血淋巴细胞酸性磷酸酶、碱性嶙酸酶、溶菌酶活性和酚氧化酶活性均有所升高;溶酶体酶膜稳定性下降;在脂多糖注射24h后,酚氧化酶原和溶菌酶mRNA表达水平均有显著上调。可见,脂多糖可以诱导河南华溪蟹血淋巴细胞溶酶体稳定性下降并且释放其中的溶酶体酶,免疫相关基因表达量上调,在一定程度上对溪蟹免疫能力有增强效果。
关键词:免疫能力;脂多糖;河南华溪蟹
中图分类号 S948 文献标识码 A 文章编号 1007-7731(2020)16-0128-05
随着水产养殖业的发展和养殖规模的不断扩大,营养、环境和新陈代谢等急剧变化的应激条件容易对各类水生动物造成疾病,甚至死亡,给水产养殖业带来了巨大的经济损失。以往使用各种抗生素来防治水生动物养殖病害,其负面影响较为严重,我国相关部门已逐步限制其在水产养殖业中的使用。近年来,对水生动物尤其是蟹类免疫机理的深入探索和认识,加上对免疫增强剂的研究,为水产养殖中病害防治开辟了一条新的思路[1]。为了防控水产养殖中出现的病害,国内外的一些学者开始将关注点投向了免疫增强剂。目前发现,多种化合物已经作为免疫增强剂投入到水产养殖业中[2-3]。
目前发现,脂多糖(lipopolysaccharide,LPS)是革兰氏阴性细菌细胞壁的主要成分,不仅可以引起机体体液免疫应答,也可以提高机体的非特异性免疫功能,有效增强虾蟹类、鱼类免疫能力[4-7],效果明显,已被应用于水产养殖的相关研究。研究表明,脂多糖可以分别激活凡纳滨对虾(Litopenaeus vannamei)[5]、中华绒螯蟹(Eriocheir si-nensis)[6]酚氧化酶原系统释放酚氧化酶(Phenoloxidase,PO),进而将酚氧化成醌,最终形成黑色素,黑色素能够使外来病菌和宿主隔离,从而达到免疫效果。还有文献表明,脂多糖刺激锯缘青蟹(Scylla paramamosain)血淋巴细胞免疫相关酶活性升高[7],血细胞吞噬功能也增强。脂多糖作为饲料添加剂可以增加黑虎虾(Alpheus bellulus)的存活率,诱导免疫相关基因的表达量上调[8]。王玉芬等[9]研究指出,竹笋多糖能增强中华绒螯蟹(Eriocheir si-nensis)的非特异性免疫反应,可添加在中华绒螯蟹词料里来提高其免疫功能。赵紫越等[10]报道,黄芪多糖可以诱导中华绒螯蟹血细胞中的抗脂多糖因子、抗菌肽、酚氧化酶和Toll样受体基因表达水平普遍上调,提高了蟹体免疫能力。此外,大黄多糖刺激拟穴青蟹(Scyllparamamo-sain)后,血清中非特异性免疫指标活性均显著提高[11]。
甲壳动物缺乏免疫球蛋白,其体液免疫主要是一些非特异性的酶或其他免疫因子来共同完成的。据报道,甲壳动物血淋巴细胞主要分为颗粒细胞和无颗粒细胞两大类,体液免疫因子酚氧化酶一般以酶原的形式即存在于颗粒细胞中[12,13]。当血细胞脱颗粒时,将酚氧化酶原释放到胞外,被激活转变成有活性的酚氧化酶。还有研究报道,溶酶体是甲壳动物血淋巴细胞中重要的亚细胞器。在一些外来物的刺激作用下,溶酶体酶的膜很容易破裂而释放出其中的水解酶(酸性磷酸酶(acid phospha-tase,ACP )、碱性磷酸酶(alkaline phosphatase ,AKP) 和溶菌酶(lysozyme,LSZ)等),进一步发挥免疫作用[14]。本研究以河南华溪蟹(Sinopotamon henaneme)血淋巴细胞为试验材料,用脂多糖作为免疫刺激剂,研究其对溪蟹免疫相关酶(酸性磷酸酶、碱性磷酸酶、溶菌酶和酚氧化酶)活性、溶酶体膜的稳定性、溶菌酶和酚氧化酶原基因mRN-NA表达水平的调节作用,探索脂多糖对溪蟹免疫功能的影响,以期为水产养殖病害防治提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 试验动物 河南华溪蟹购自太原市五龙口水产批发市场。采用曝气自来水,于循环系统的水族缸中暂养两周以上,温度为15~18℃,连续充气,养殖密度为4~5只/L,每2d喂食1次。
1.1.2 主要试剂 脂多糖(E. Coli,055: B5,L2880,Sig-ma);中性红(Sigma);乙醇、氯仿和异丙醇均为分析纯;四种酶的活性及蛋白质含量测定试剂盒均购于南京建成生物工程研究所。总RNA提取试剂盒(RNA isoPlus)、反转录试剂盒(Prime ScriptTM RT reagent Kit with gDNAEraser)、实时荧光定量PCR试剂盒PremkTaqTM(Ex TaqTMVersion 2.0 plus dye)均为Takara试剂盒。ProO、LSZ和内参基因β-actin的引物均由上海生工有限公司合成。
1.1.3 主要仪器 超低温冰箱(Thermo Forma,美国);冷冻离心机(Eppendorf 5804R,德国);多功能酶标仪(Spec-tra max,M5,美国)。普通PCR仪(Eppendorf Mastercyclergradient,德国)、凝胶成像系统(Alpha FluorChem HD2,美国)、微量分光光度计(Eppendorf,Hamburg,德国)和实时荧光定量PCR仪(ABI 7500,美国)。
1.2 试验方法
1.2.1 试验设计 暂养结束后,随机选取健康活泼和個体大小一致(平均湿重为20.0±0.5g)的,附肢完整的溪蟹个体,分成5组,每组10只个体。设置1个空白对照组(无菌生理盐水)和4个脂多糖作用组,浓度分别是1.0、2.0、4.0和8.0μg/mL,刺激12h、24h,取血淋巴进行后续实验。
1.2.2 样品制备 将每组溪蟹个体分别置于冰上,用无菌注射器从溪蟹第4步足基部抽取血淋巴,将所取得的血液与等体积的抗凝剂同时装入离心管混匀,离心弃上清,制成106的细胞悬液,0.5mL的样品经液氮速冻,以备提取RNA用;一部分用于溶酶体膜稳定性的测定;剩余的样品-80℃保存,待用。
1.2.3 溶酶体膜的稳定性将20μL的血细胞悬浮液放入离心管中,每管加入40μL的中性红溶液(用TBS缓冲液配制成0.33%的液体),在10℃孵育2h,200g离心5min,用TBS缓冲液洗涤;加入200μL的混合液(1%冰醋酸:50%乙醇=1:1)混匀后,孵育。在多功能酶标仪上,测定其吸光值,结果用O.D./mg/mL蛋白来表示。
1.2.4 酶活性测定 ACP、AKP、LSZ和P0活性均依据南京建成试剂盒中说明书中的方法来测定。
1.2.5 proPO,LSZ mRNA表达水平的检测以所有处理组合成的cDNA为模板,在实时荧光定量PCR仪上进行定量试验。每个样品重复3次进行定量。数据结果由公式2-△△Ct计算出基因的表达量。试验所用引物序列见表1。
1.3 数据处理 采用统计软件SPSS20.0中的AN0VA法对各组间数据进行差异分析,其中P<0.05表示有显著性差异,P<0.01表示有极显著性差异。用软件Origin 10.0作图。
2 结果与分析
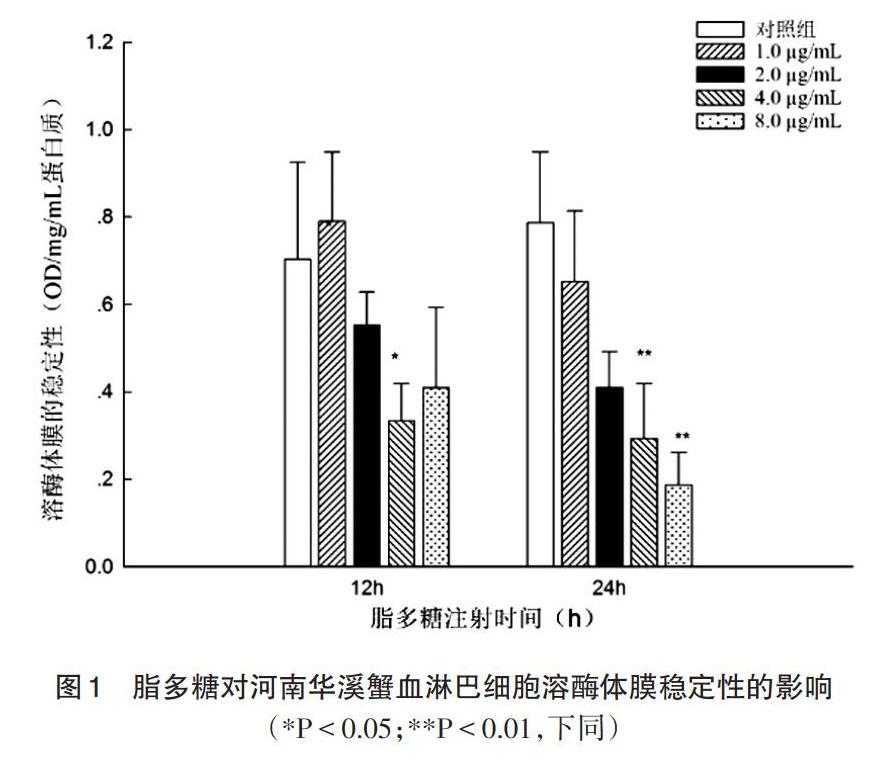
2.1 脂多糖对河南华溪蟹血淋巴细胞溶酶体膜稳定性的影响 由图1可知,脂多糖刺激溪蟹12h,血淋巴细胞溶酶体膜的稳定性有所下降,其中,当脂多糖浓度是4.0μg/mL时,溶酶体膜的稳定性较对照组相比显著下降。脂多糖刺激溪蟹24h,脂多糖浓度分别是4.0和8.0μg/mL时,与对照组相比,血淋巴细胞溶酶体膜的稳定性极显著下降(P<0.01)。
2.3 脂多糖对河南华溪蟹血淋巴细胞酚氧化酶原基因表达水平的影响 由图3可知,脂多糖刺激溪蟹后,血淋巴细胞中酚氧化酶原基因表达水平呈现上升趋势。在脂多糖(2.0、4.0和8.0μg/mL)作用12h时,酚氧化酶原
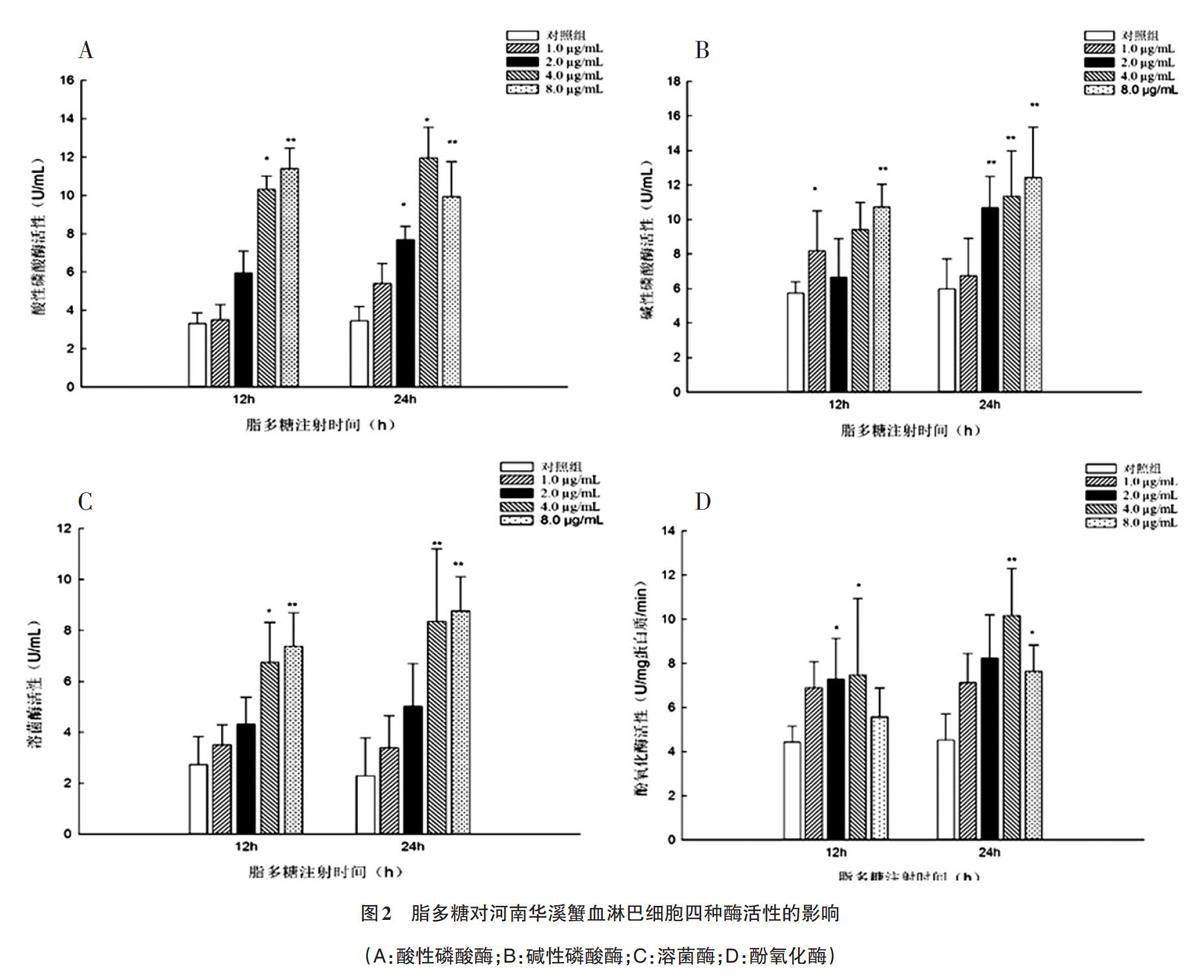
2.2 脂多糖对河南华溪蟹血淋巴细胞ACP、AKP、LZM及PO活性的影响 由图2可知,溪蟹血淋巴细胞中酸性磷酸酶活性随着脂多糖注射浓度增加而逐渐升高,在脂多糖为4.0和8.0μg/mL时,酸性磷酸酶活性显著升高。碱性磷酸酶活性和溶菌酶活性变化与酸性磷酸酶活性变化趋势大体上一致(图2B、2C)。当脂多糖注射时间延长至24h,除低浓度组(1.Oμg/mL)外,其他脂多糖浓度组作用下,碱性磷酸酶活性出现了极显著升高(P<0.01)。溶菌酶活性在较高浓度的脂多糖(4.0和8.0μg/mL)作用时,也出现了显著升高现象(P<0.05或者P<0.01)。从图2D看出,随着脂多糖作用时间延长,酚氧化酶活性一直升高。当脂多糖浓度是4.0μg/mL时,其活性达到峰值,在较高浓度8.0μg/mL时,酚氧化酶活性有所下降。基因表达水平分别是对照组的3.6、5.5、5.6倍。随着作用时间延长至24h,酚氧化酶原基因表达水平一直升高,在较高浓度时,其表达水平达到对照组的8.6倍。
2.4 脂多糖对河南华溪蟹血淋巴细胞溶菌酶基因表达水平的影响 由图4可知,在脂多糖(2.0和8.0μg/mL)作用12h时,溶菌酶基因表达水平分别是对照组的3.3和5.5倍(P<0.01)。随着作用时间延长至24h,溶菌酶基因表达水平一直升高,在较高浓度(8.0μg/mL)时,其表达水平达到对照组的6.7倍。注射脂多糖后,溪蟹血淋巴细胞中溶菌酶基因表达水平整体上呈现上升趋势。
3 讨论
甲壳动物的免疫机能主要依赖非特异性免疫,由血淋巴细胞或者从血淋巴细胞释放出来的免疫因子来共同完成。其中,血淋巴细胞中的亚细胞器溶酶体在免疫识别和防御中起着重要作用。溶菌酶是甲壳动物免疫系统中的重要成分,能够水解革兰氏阳性菌细胞壁的多糖,并使之水解释放出来,形成水解酶体系,破坏和消除入侵外来物。酸性磷酸酶是动物体内巨噬细胞溶酶体的标志酶[15],也是甲壳动物溶酶体酶的重要组成部分,它可以通过水解表面带有磷酸酯的异物而破坏异物。碱性磷酸酶活性的提高,可以加速相应组织的物质代谢,为机体提供所需的无机磷,积累更多的能量。杨季芳等人报道,从南极海洋寡营养细菌提取的脂多糖能显著提高三疣梭子蟹(Portunus trituberculatus)血清中酸性磷酸酶和碱性磷酸酶活性[16],免疫增强效果较好。同样,本研究结果显示,微量的脂多糖可以引起河南华溪蟹血淋巴细胞溶酶体膜稳定性下降,溶酶体酶(酸性磷酸酶和碱性磷酸酶)活性升高,这与谢鹏关于脂多糖对凡纳滨对虾血细胞的研究结果也类似[17]。文英等研究发现,大黄多糖刺激拟穴青蟹后,血清中酸性磷酸酶、碱性磷酸酶、酚氧化酶和溶菌酶等非特异性免疫指标活性均显著提高[11],与本研究结果一致。
据报道,脂多糖可以刺激甲壳动物血淋巴细胞脱颗粒释放免疫因子如酚氧化酶来增强机体免疫能力[18]。甲壳动物体内酚氧化酶激活系统在免疫识别和防御中发挥着关键作用,该系统以非活化状态存在于颗粒细胞中,脂多糖可引起细胞胞吐作用,激活酚氧化酶原系统,伴随着大量免疫活性因子的产生,通过多条途径参与免疫作用。Soderhall等曾报道了脂多糖可以促使机体产生一种激活蛋白酶,从而使无活性的酚氧化酶原转化为有活性的酚氧化酶[19]。研究发现,大黄多糖刺激拟穴青蟹4d后,血细胞中的酚氧化酶原基因表达水平上调至对照组的2.1倍,酚氧化酶活性也显著升高[11]。潘清清[20]。证实大黄多糖可显著提高锯缘青蟹血淋巴酚氧化酶活力,这与本论文中脂多糖刺激河南华溪蟹后,血淋巴细胞中酚氧化酶原表达水平上调,酚氧化酶活性也增加的结果相似。脂多糖可以通过改变梭子蟹血清中的溶菌酶活力及血细胞吞噬活性来提高梭子蟹的非特异性免疫功能[21]。还有研究报道,脂多糖能显著提高三疣梭子蟹(Portunustrituberculatus)血清中溶菌酶活力[16]。尼罗罗非鱼(Ore-chromis niloticus)在脂多糖刺激后,参与其非特异性免疫的结构域接合子蛋白(TIRAP)基因表达上调[22]。本研究结果表明,注射脂多糖后,河南华溪蟹血淋巴细胞中的溶菌酶基因表达水平上调,伴随着溶菌酶活力也升高,这与上述报道结果是相同的。
综上所述,脂多糖可以通过改变河南华溪蟹血清中的酸性磷酸酶、碱性磷酸酶、溶菌酶和酚氧化酶活力及免疫相关酶基因表达来提高溪蟹的非特异性免疫功能,但是否影响溪蟹血淋巴细胞的亚显微结构,是否对非特异性免疫其他相关因子有影响,以及通过什么途径来提高溪蟹免疫能力,还有待进一步研究。
参考文献
[1]ChenY Y,Chen J C,Kuo Y H,et al. Lipopolysaccharide and β-1,3-glucan-binding protein (LGBP) hind to seaweed polysaccharidesand activate the prophenoloxidase system in white shrimp Litope-naeus vannamei[J]. Dev Comp Immunol,2016,55 : 144-151.
[2] Wang Y j,Hu M H,Chiang M W L,et al. Characterization of subpopulations and immune-related parameters of hemocytes in thegreen-lipped mussel Perna viridis [J]_. Fish Shellfish Immunol,2012,32:381-390.
[3]周妍英,井维鑫,董峰,等.脂多糖对镉致河南华溪蟹免疫因子毒性的调节作用[J].生物技术通讯,2016,27(6):816-819.
[4]Burge E J,Madigan D J,Burnett L E,et al. Lysozyme gene expres-sion by haemocytes of Pacific white shrimp, Litopenaeus vanna-mei, after injection with Vibrio[J]. Fish Shellfish Immunol, 2007,22(4):327-339.
[5]0kumura T. Effects of lipopolysaccharide on gene expression of an-timicrobial peptides (penaeidins and crustin) , serine proteinaseand prophenoloxidase in haemocytes of the Pacific white shrimp,Litopenaeus vannamei [J]. Fish Shellfish Immunol,2007,22 (1-2):68-76.
[6]张行.脂多糖和β-1,3-葡聚糖结合蛋白(LGBP)激活中华绒螯蟹(Eriocheir sinensis)酸氧化酶原系统的研究[D].上海:华东师范大学,2016.
[7]Gopalakrishnan S,Chen F Y, Harikrishnan T,et al.Modulation andinteraction of immune-associated parameters with antioxidants inthe immunocytes of crab Scylla paramamosain challenged with li-popolysaccharides[J]. Evid Based Complement Ahern Med,2011:824-962.
[8] Rungrassamee W, Maibunkaew S, Karoonuthaisiri N, et al. Applica-tion of bacterial lipopolysaccharide to improve survival of theblack tiger shrimp after Vibrio harveyi exposure [J] Dev Comp Im-munol, 2013,41 (2) : 257-262.
[9] 王玉芬,李義,陈亚军,等.竹笋多糖对免疫抑制中华绒螯蟹非特异性免疫的影响[J].伺料研究,2014,7:1 -4.
[10] 赵紫越,高彤,袁春营,等.黄芪多糖和低浓度嗜水气单胞菌对中华绒螯蟹免疫相关因子基因表达的影响[J].伺料研究,2020,43(03):41-44.
[11]文英,曹劲松,张泽蕙,等.大黄多糖对拟穴青蟹免疫相关基因表达的影响[J].中国水产科学,2015,22(03) :387-392.
[12]周妍英.镉对河南华溪蟹血淋巴细胞的毒性效应及脂多糖的调节作用[D].太原:山西大学,2017.
[13]Zhou Y Y,Luo Z M,Wang L. Modulation of oxidative damage bylipopolysaccharides and antioxidant responses in the freshwatercrab Sinopotamon henanense Bott 1967 (Decapoda: Brachyura:Potamonidae) during cadmium exposure [J]. Journal of Crusta-cean Biology, 2019,39(4): 477-484,
[14]Zhou Y Y,Jing W X,Dahms H U,et al. Oxidative damage, ultra-structural alterations and gene expressions of hemocytes in thefreshwater crab Sinopotamon henanense exposed to cadmium [J].Ecotoxicol Environ Saf,2017,138: 130-138.
[15] 周进,宋晓玲,王秀华,等.A3肽聚糖对牙鲆不同组织中超氧化物歧化酶及磷酸酶活力的影响[J].中国水产科学,2004, 11(4):296-302.
[16] 杨季芳,郭卢云,陈福生,等.2株南极海洋寡营养细菌(A1-teromonas stellipolaris)脂多糖对三疣梭子蟹(Portunus tritubercu-latus)非特异性免疫活性的影响[J].海洋与湖沼,2011,42(02):294-299.
[17]谢鹏.脂多糖、多巴胺对凡纳滨对虾血细胞作用机制的研究[D].青岛:中国海洋大学,2010..
[18]Xu H S,Lyu S J,Xu J H et al. Effect of lipopolysaccharide onthe hemocyte apoptosis of Eriocheir sinensis [J]. J Zhejiang UnivSci B,2015,16(l2):971-979.
[19]Soderhall K,Lee S Y. Early events in crustacean innate immunity[J]. Fish and Shellfish Immunology,2002,12:421-437.
[20]潘清清.锯缘青蟹免疫增强剂的筛选及在病害防控中的应用[D].武汉:华中农业大学,2008.
[21]王海丽,黄幸雷,杨季芳.南極寡营养细菌ANT52的脂多糖对梭子蟹非特异性免疫功能的影响[J].浙江万里学院学报,2011,24(6):78-83.
[22]刘洁,高风英,卢迈新,等.尼罗罗非鱼TIRAP基因克隆、组织表达及其在无乳链球菌、脂多糖和聚肌胞苷酸刺激下的免疫应答[J].大连海洋大学学报,2020,doi.org/10.16535/j.cnki.dlhyxb.
(责编:张宏民)
基金项目:山西省高等学校科技创新项目(2019L0844);忻州师范学院科研基金项目(2018KY20)。
作者简介:周妍英(1984-,女,山西人,博士,讲师,研究方向:重金属污染对水生动物毒性效应。 *通讯作者 收稿日期:2020-06-30
