麦类作物籽粒硬度的研究进展
2020-10-28安江红张文静赵瑛琳南金生
安江红,张文静,赵瑛琳,韩 冰,南金生
(1.内蒙古农业大学生命科学学院,内蒙古呼和浩特 010011;2.内蒙古科技信息网络工程技术研究中心,内蒙古呼和浩特 010020)
普通小麦(Triticum aestivum L.)、大麦(Hordeum vulgare L.)、燕麦(Avena sativa L.)和黑麦(Secale cereale L.),是世界上重要的粮食作物。小黑麦是第1种人工杂交产生的新型作物,结合了小麦和黑麦种质与籽粒产量的优良特性,具有抗病和抗逆的优势。
籽粒硬度是指破坏籽粒时受到的阻力,即破坏籽粒需要的力[1]。籽粒硬度是国际较通用的商品小麦分类的重要指标,是市场分级和定价的重要性状之一,影响润麦加水量、出粉率、破损淀粉数量和面粉颗粒度,决定磨粉品质和食品加工品质[2]。大麦是世界上重要的饲料作物和啤酒工业的原料,籽粒硬度影响大麦麦芽品质,且籽粒颗粒大小还影响动物的消化和吸收[3]。燕麦是粮饲兼用作物,是世界性的栽培作物,分为裸粒型(裸燕麦)和带稃型(皮燕麦)两大类。籽粒硬度影响燕麦米的成米率、燕麦粉颗粒度和燕麦片的爽滑度等[4]。笔者就麦类作物籽粒硬度研究进行了综述,旨在更好地了解麦类作物籽粒硬度的研究现状,从而为其遗传改良和育种提供理论基础。
1 籽粒硬度检测方法
麦类作物中,小麦籽粒硬度检测方法研究最为全面和深入。测定小麦籽粒硬度的传统方法[5-8]、国标方法[9]以及近年来发展的快速测定方法比较多[10-12](表1),以力学方法最为经典[13]。其中,近红外光谱法(near infrared reflect,NIR)、单籽粒谷物特性测定仪法(single kernel characterization system,SKCS)与现行国标方法硬度指数法(GB/T 21304—2007)之间有很好的一致性[14-16]。也有研究用其他方法检测谷物籽粒硬度,例如针尖压入法[17]、基于可见-近红外光谱的径向基函数(RBF)神经网络模型[18]、质构仪法等[4,19-20]。麦类作物的籽粒具有类似的物理结构特点,但在检测籽粒硬度时应选择适合的方法,否则无法表征其真实的硬度。例如小麦籽粒短圆,而燕麦籽粒普遍偏细长,单籽粒较小麦轻,在使用SKCS 法进行检测时由于无法均匀地使单一籽粒落入称量斗,导致测量值不准确,SKCS 法不适于燕麦籽粒硬度检测。另外,燕麦具有高脂肪的特点,其脂肪质量分数高达6%~8%,这一特点导致燕麦粉极易黏结成块,无法顺利通过筛网,影响硬度测量的准确性,因此,基于碾磨的方法也不适用于燕麦籽粒硬度检测[20]。质构仪法可以实现单籽粒的压缩,不需要磨粉,适合没有标准检测方法的作物籽粒硬度检测。
2 籽粒硬度与籽粒成分的关系
麦类作物籽粒组分主要包括淀粉、蛋白质、脂质和膳食纤维等。籽粒胚乳质地是籽粒化学成分综合作用的结果[21],麦类作物籽粒的成分与籽粒硬度直接相关。
利用扫描电镜观察硬质与软质小麦籽粒的断面,发现软质小麦的淀粉近似球形,而硬质小麦籽粒断面破损淀粉颗粒较多,胚乳结构相对致密[22]。籽粒中淀粉颗粒和蛋白质基质间的黏结力以及包被淀粉颗粒的蛋白质基质连续性共同决定了小麦籽粒硬度[23]。研究发现,这种蛋白质主要存在于水洗淀粉表面,大小约15 kDa,命名为Friabilin 蛋白,在软质小麦中的表达量高[24],约为硬质小麦的10倍。燕麦籽粒比小麦籽粒软,在使用相近的力碾压籽粒后,燕麦成片状,而小麦成粉状(图1)。与小麦类似,软质燕麦的淀粉粒之间空隙较大,与蛋白呈分离状态,而硬质燕麦淀粉粒紧凑,与蛋白紧密相连,细胞壁空隙小(图2)[4]。另外,燕麦中脂质含量丰富,通常与直链淀粉形成淀粉-脂质复合物[25-26],而影响淀粉的生理特性,未脱脂燕麦淀粉的破损颗粒较多,脱脂后破损颗粒减少,颗粒也变得均匀[27]。燕麦脂质可能参与淀粉和蛋白的结合,共同影响籽粒硬度。
小麦籽粒硬度是在籽粒脱水过程中形成的[28-29]。小麦的硬度指数与籽粒中水分的变化在一定范围内正相关[30],12%~13%的水分含量是小麦抗粉碎硬度指数最佳灵敏范围[31]。燕麦与小麦类似,籽粒水分含量在10.62%~11.84%时,硬度上升最显著[4]。β-葡聚糖、阿拉伯基木聚糖是细胞壁的结构多糖,在胚乳细胞壁相互连接,增大了细胞壁的支撑性,大麦籽粒中的非淀粉多糖含量与籽粒硬度存在正相关关系[32],这与臧容宇[4]在燕麦上的研究类似,硬质燕麦糊粉层细胞壁厚度显著高于软质燕麦,但李梦黎等[20]通过检测β-葡聚糖含量,发现其与燕麦籽粒硬度的相关性不显著。β-葡聚糖是燕麦重要的功能成分,其与籽粒硬度的相关性有待进一步研究。
3 麦类作物籽粒硬度分子机制研究进展
3.1 小麦籽粒硬度的分子机制
麦类作物中,对小麦籽粒硬度的研究是最早开始的(图3)。人们对小麦籽粒硬度的认识始于19世纪末[33],到20世纪后半叶,小麦分级制度的建立促进了小麦籽粒硬度的遗传研究。早期研究表明,籽粒硬度的遗传力高,受一个或两个主效基因和一些修饰基因控制[34]。到1986年,GREENWELL和SCHOFIELD[35]首次用1% SDS 从小麦水洗淀粉中发现了Frabilin 蛋白,使得小麦籽粒硬度的分子生物学研究有了突破性进展。根据高效液体层析法和氨基酸序列分析,证实了Frabilin 蛋白主要由两种蛋白构成,因其富含色氨酸区域,故将其命名为puroindoline a(Pina)和puroindoline b(Pinb)[36-37]。Pinb 编码的蛋白简称为PINA,Pinb 编码的蛋白简称为PINB,二者氨基酸序列同源性达到60%。Pina和Pinb 基因启动子上游序列的同源性高达78.4%,而转录起始位点后400 bp 范围内相似性为54.8%,且两个蛋白质氨基酸的差异主要发生在末端序列第10~20个氨基酸残基之间[38]。PINA蛋白的缺失或编码PINB蛋白的基因突变均造成小麦胚乳质地变硬[2]。KRISHNAMURTHY 等[39]将野生型Pina和Pinb 基因转入水稻,转基因水稻胚乳质地明显变软。BEECHER 等[40]将我国春野生型Pinb 序列转入硬粒小麦,使其在胚乳中表达,SKCS 硬度值由70 降为25。也就是说,小麦puroindoline 基因可以用于谷物籽粒硬度性状的修饰。小麦在开花后10 d内PIN 蛋白含量极低,从第15 天后开始快速增加,到32 d 到达最大值,软质麦的PIN 蛋白含量高于硬质麦[41]。
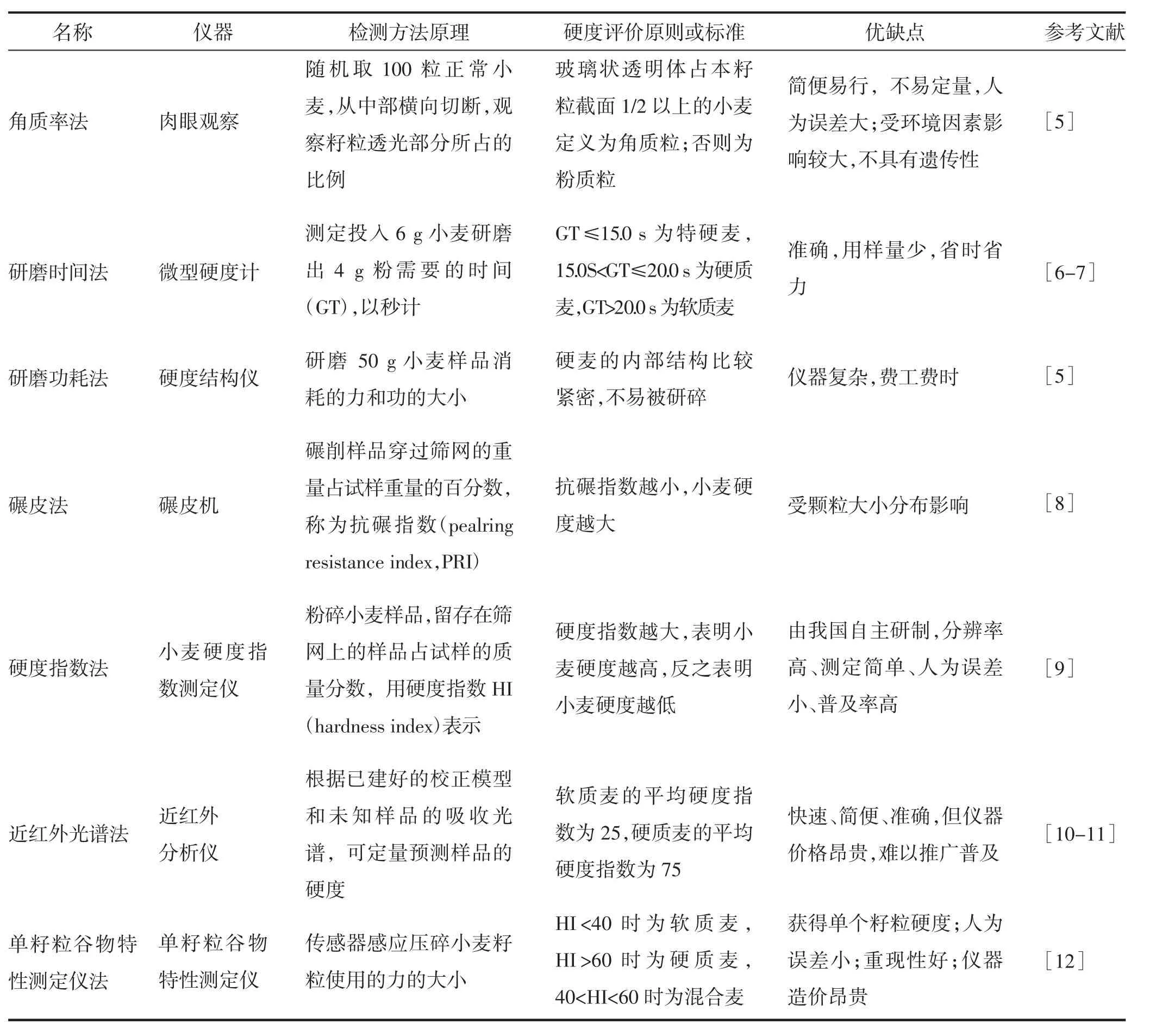
表1 小麦籽粒硬度检测方法
3.2 燕麦籽粒硬度的分子机制
燕麦籽粒硬度的研究比小麦晚。1998年,TANCHAK 等[42]发现了具有与小麦puroindoline 基因相似特点的两对燕麦cDNA 克隆,分别是3B3-5、3B3-7和3B3T-3、3B3T-5。其中3B3和3B3T分别编码147和142个氨基酸,均含有典型的10个半胱氨酸骨架和1个色氨酸丰富区,故命名为tryptophanins。该基因在燕麦、小麦、大麦和黑麦籽粒中特异性表达,在水稻籽粒中不存在。2000年,GAUTIER 等[43]利用小麦籽粒硬度基因Pina-D1和Pinb-D1的特异性引物在燕麦中PCR 扩增得到了puroindoline-like的基因,并命名为avenoindolines。通过序列比对发现avenoindolines 与puroindolines的相似性要大于与tryptophanins的相似性。2015年,GAZZA等[44]在两个普通栽培燕麦品种Donata和Primula中利用RT-PCR和Western blotting的方法并没有检测到Ainb的转录本和AINB蛋白,同时Aina的转录本和AINA蛋白的含量很低,表明AINA和AINB在燕麦籽粒硬度中的作用可能是微效的。但在水洗淀粉表面发现另外3个14 kDa的蛋白,都包含一个含4个色氨酸的疏水区,分别命名为Vromindoline 1(Vin-1)、Vromindoline 2(Vin-2)和Vromindoline 3(Vin-3),分别由Vin-1、Vin-2和Vin-3 基因编码,且在燕麦A、C和D 基因组上都存在。通过向硬粒小麦(无Pina和Pinb的同源基因)Svevo 中转化Vin-D2a和Vin-A3a 基因,均在转基因后代中表达并在淀粉粒表面积累了其编码的蛋白VIN-2.1和VIN-3.1,使得转基因硬粒小麦Svevo的籽粒硬度降低约50%,表明vromindolines 基因确实可以引起籽粒硬度的改变,可以用于其他谷物籽粒硬度性状的修饰。皮燕麦中,Vin-2 基因在开花后7 d开始表达,14 d 开始在淀粉颗粒表面积累,在21 d表达量达到峰值,28 d在淀粉颗粒表面积累到最大值,42 d后开始逐渐下降。
3.3 其他麦类籽粒硬度的分子机制
大麦、黑麦和小黑麦的籽粒硬度研究开展较晚。2001年,BEECHER 等[45]首次从8种常见的北美栽培大麦中克隆出Pina和Pinb的同源序列hordoindoline a(Hina)和hordoindoline b(Hinb),通过序列比对发现,大麦的序列含有非保守氨基酸替换。Hordoindolines 基因定位在大麦基因组的5H 上,该区域可以解释22%的SKCS 硬度差异[46]。大麦中,HINA在开花后5 d 开始表达,8 d 是HINA表达的最高峰,20 d后表达量逐渐降低,而HINB在开花后8 d 才被发现,20 d 才逐渐减少[47]。
六倍体小黑麦是将小麦和黑麦经过人工杂交产生的一种新型作物,主要包括初级小黑麦和林保小黑麦两种类型:初级小黑麦是由硬粒小麦和黑麦杂交产生,不存在普通小麦具有的硬度位点而具有黑麦中的硬度位点,该位点编码的Secaloindoline(SIN)蛋白能使胚乳质地变软;林保小黑麦是初级小黑麦与普通小麦杂交产生的后代,来自黑麦或者小麦的硬度位点都能够遗传给后代,在子代中硬度的变化范围很大,从极软到极硬都有分布[48]。小黑麦硬度位点位于5R 染色体上,包含两个基因为Sina和Sinb,分别编码SINA和SINB。研究发现,软硬质小黑麦secaloindoline 基因的编码区完全一致,但二者表达的淀粉结合型Friabilin 差异很大,这可能是因为它们在启动子区存在差异[49](图3)。
4 籽粒硬度与品质性状的关系
品质性状主要包括外观品质、加工品质和营养品质等。加工品质包括磨粉品质和食品加工品质两个方面。衡量磨粉品质的指标主要包括出粉率、灰分含量和面粉色泽;食品加工品质包括面粉品质、面团品质、烘焙品质和蒸煮品质等。
小麦籽粒硬度对磨粉品质和食品加工品质均具有重要影响。硬质小麦出粉率较高、破损淀粉颗粒多、吸水率高,发酵时易膨胀,适宜制作面包;软质小麦淀粉粒较细,制作出的面条口感爽滑[50]。研究表明,小麦籽粒硬度与灰分含量呈显著或极显著正相关[51];燕麦籽粒硬度与灰分含量、面粉白度显著相关,与脂肪含量、淀粉糊化温度等相关性不显著[12];大麦籽粒硬度与籽粒的吸水性显著负相关[52]。
麦类作物籽粒硬度基因及其不同的等位变异类型对其加工品质也有重要影响。小麦籽粒硬度基因野生型Pina-D1a和Pinb-D1b的等位变异基因类型有很多,例如大片段缺失的变异类型,Pina-D1b(缺失15 380 bp)、Pina-D1r(缺失10 415 bp)、Pina-D1s(缺失4 420 bp)、Pina-D1u(缺失6 460 bp),还有一些由少数位点碱基突变和插入缺失产生的变异类型Pina-D1m、Pina-D1n、Pina-D1l,其中Pina-D1m 基因编码的蛋白质第35位的脯氨酸变为色氨酸,导致胚乳质地变硬[53-56]。Pinb 也有多种等位变异基因,例如Pinb-D1h、Pinb-D1i、Pinb-D1j、Pinb-D1k、Pinb-D1m、Pinb-D1n、Pinb-D1o,这些均为多位点突变,表现为软质[57]。这些puroindoline 基因位点发生变异会导致小麦籽粒产生不同程度的硬度变化,进而影响加工品质。小麦面粉中增加puroindoline 蛋白利于改善面包结构,使气孔大小分布均匀,但会造成面包体积下降[58]。Pinb-D1b的小麦具有较好的磨粉品质和优良的面包体积[59],比Pina-D1b的出粉率高、面粉颗粒大、黏度仪崩解值小[60]。Pinb-D1d 基因型小麦的出粉率、面粉亮度较高,馒头色泽和张弛性评分较高,有较好的馒头制作品质;而Pinb-D1g和Pinb-D1e基因型的中国湿面条的评分较高[61]。
5 展望
目前,籽粒硬度的研究与前沿技术相结合,例如,已有研究利用CRISPR/Cas9 介导的基因编辑技术对Pinb 基因启动子区进行编辑,为在小麦中实现Pinb 基因的定点突变奠定了基础[62];也有研究进行了小麦籽粒硬度的蛋白质组学分析,为进一步探索小麦籽粒硬度的遗传机制提供了参考[63]。
小麦籽粒硬度的研究较为系统且贴近生产生活,在研究深度和广度上都走在其他麦类作物前列,但燕麦、大麦和黑麦等作为优良的杂粮作物有广阔的发展前景和更大的经济效益,可以为麦类作物育种提供更多优异基因。麦类作物籽粒硬度遗传机理的深入解析,对籽粒硬度优异等位基因挖掘、分子标记的开发、麦类作物品质育种与改良以及指导生产生活等方面都有重要意义。
