缺磷培养下黑木相思苗木的生理生化响应
2020-10-27裘珍飞范春节曾炳山
裘珍飞 范春节 曾炳山
( 中国林业科学研究院热带林业研究所,广东 广州 510520)
黑木相思(Acacia melanoxylon)原产于澳大利亚南部,是相思类最高大乔木之一,具有固氮、速生、材质优良,且有一定的耐干旱、耐瘠薄、耐寒冷等特点。20 世纪90年代引进我国后,通过系统选育,获得生长迅速、树干通直、木纹美丽和耐寒性强的优良无性系,并在我国华南地区推广应用,种植规模日益扩大,现已成为速生丰产,珍贵用材和生态公益等多用途树种[1]。由于我国南方土壤普遍呈酸性,有效磷(P)含量低,经营人工林时因缺P 影响生长量[2]。生产中通过加施P 肥提高植物可利用P 水平,但过度的P 施用会导致成本增加、水土污染、P 矿资源枯竭等社会和环境问题[3]。因此,研究P 胁迫对苗木生长和养分含量及相关生理指标的影响,对了解黑木相思适应P 胁迫的机制和栽培中P 养分的调控都具有重要的科学意义。
植物长期处在P 胁迫下,其形态、营养吸收和分配、生理生化及光合性能等均会发生适应性变化[4]。研究表明低P 胁迫下,植物的地上部干物质量降低,根系体积和表面积增加,从而引起根冠比、比根长等指标的增加[5-6];也引起植物体内超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等保护酶系统的抗氧化胁迫反应[7-8];P 胁迫影响叶片中光合色素含量及光能利用能力和效率[9-10];长时间和重度P 胁迫造成植物新陈代谢紊乱,引起体内养分吸收失去平衡,改变植物体内化学组分[11-12]。P 胁迫下作物如何响应和适应一直是国内外的研究热点之一。目前,黑木相思的研究主要集中在选育和繁育上,有关P 胁迫下,黑木相思地上部及根系形态的变化;内部产生的生理生化响应机制及根、茎、叶各器官对养分吸收和利用的影响等相关内容尚未见研究。本试验旨在揭示黑木相思适应P 胁迫的机制,为我国华南地区的黑木相思高效栽培和P 肥的科学合理施用提供参考依据。
1 材料和方法
1.1 试验材料
以黑木相思组培苗为材料,选取生长统一的生根瓶苗(苗高(2.71±0.08) cm,全株鲜物质量(0.033±0.01) g),培养在海绵支撑的育苗盘中,每盘种植20 株,盘内浇灌营养液。
1.2 试验方法
1.2.1 试验设计
设置全P 处理浇灌Hoagland’s 全营养液[13],缺P 处理浇灌Hoagland’s 营养液中不添加KH2PO4(136 mg/L),但加入KCl(74.5 mg/L)用于平衡钾离子,其他营养液元素组成不变。整个试验在热带林业研究所温室中进行,每处理3 盘,共6 盘,培养3个月后测定各项指标。
1.2.2 测定方法
生物量、根系形态及养分测定。每盘随机抽取苗木10 株,每处理共30 株,小心清除根部海绵支撑物,测量苗高和苗杆直径,并记录每苗叶片数,称重全株鲜物质量。把植株分成根、茎、叶3 部分,根系采用ScanMaker i800plus 彩色平台扫描仪(上海中晶科技,中国)扫描图片,经相应软件分析后获得根系形态参数。收集上述根、茎、叶,在 105 ℃恒温烘干箱中杀青30 min,后在70 ℃下烘干至质量恒定,分别称量根、茎、叶的干物质量。利用烘干的样品,采用凯氏滴定法测定全氮(N)含量,采用钼锑抗比色法测定全P 含量,采用火焰原子吸收分光光度法[14]测定全钾(K)含量。按公式(1)~(3)计算根冠比、养分利用效率、养分积累量。

叶绿素含量测定。称取叶片0.2 g,碾磨后用80%丙酮萃取、过滤、洗涤至叶子碎片无色,定容50 mL,采用UV-120 分光光度计(津岛,日本)测定[15]。
氧化酶活性测定。称取叶片0.2 g,加0.1 mol/LPH 为7.4 的磷酸缓冲液于冰水浴条件下制备成10%组织匀浆,用Allegra X-30 R 高速冷冻离心机(Beckman Coulter,美国)离心20 min,设置温度为4 ℃,转速10 000 r/min,静置后取上清液测定氧化酶活性,采用氮蓝四唑法测定SOD 活性,采用紫外吸收法测定CAT 活性,采用愈创木酚法[16]测定POD 活性。
光合参数测定。采用LI-6800 型便携式光合测定仪(LI-COR,美国)测定叶片净光合速率(A)等光合参数。全P 和缺P 处理各选取4 株苗木,每苗木固定1 片中部的成年叶作为测定对象,测定时间为9:00—11:00。采用光合测定仪自带的光源控制光合有效辐射(PAR)分别为1 800、1 500、1 200、1 000、800、600、400、200、100、50、25、0 μmol/(m2·s),共12个水平。由于黑木相思成年叶为狭窄叶,按照实际被夹在叶室中的叶面积对测定的A进行换算,依据光响应曲线的经典Farquhar 模型[17]进行拟合,获得光补偿点(LCP)、光饱和点(LSP)、最大净光合速率(Amax)、表观量子效率(AQY)及暗呼吸速率(Rd)等光合参数。
1.3 数据分析
试验数据用SPSS 16.0 进行方差分析及差异显著性检验,用Excel 2007 处理图表。
2 结果与分析
2.1 缺P 对黑木相思形态和生长的影响
由图1 可知,与全P 处理相比较,从外观上看,缺P 处理下,苗木地上部分生长明显细弱,叶片稀疏,而根系部分相对发达,特别是细根增多。由表1 可知,缺P 处理下,叶片数和苗杆直径极显著下降(P<0.01),苗高和全株鲜物质量显著下降(P<0.05),而根鲜物质量和干物质量无显著变化,根冠比以缺P 处理极显著高于全P 处理(P<0.01)。

图1 全P 和缺P 处理下黑木相思苗木生长形态Fig. 1 Growth morphology of A. melanoxylon seedlings under normal P and P deficiency

表1 全P 和缺P 处理下黑木相思苗木生长状况Table 1 Growth status of A. melanoxylon seedlings under normal P and P deficiency
2.2 缺P 对黑木相思根系形态参数的影响
由表2 可知,缺P 处理下黑木相思苗木总根长极显著高于全P 处理(P<0.01),增加了22.6%,而根的平均直径以全P 处理极显著大于缺P 处理(P<0.01),总根表面积和体积以缺P 处理略大于全P 处理,但处理间无显著差异,表明黑木相思苗木通过增加须根量,保持根的表面积和体积来适应缺P 环境。

表2 全P 和缺P 条件下黑木相思苗木根系形态参数Table 2 Root parameters of A. melanoxylon seedlings under normal P and P deficiency
2.3 缺P 对黑木相思氧化保护酶活性的影响
由表3 可知,对比全P 处理,缺P 处理在叶片和根系中SOD、POD、CAT 活性均显著(P<0.05)或极显著(P<0.01)升高。3 种酶活性升高幅度以叶片中的POD、CAT 较大,增幅达113.7%和473.3%,叶中SOD 及根系中的3 种酶活性增幅为26.7%~70.4%。

表3 全P 和缺P 培养下黑木相思根和叶中SOD、POD 和CAT 活性Table 3 SOD, POD and CAT activity of leaves and roots of A. melanoxylon seedlings under normal P and P deficiency U/mg
2.4 缺P 对黑木相思叶片光合参数的影响
由图2 可知,在PAR 强度范围内,缺P 处理下的A低于全P 处理,随着PAR 增强,2 种处理A的差值逐渐加大。通过拟合光响应曲线,获得的光响应特征参数比较(表4),缺P 处理的LSP 比 全P 降 低652.8 μmol/(m2·s),LCP 提 高14.4 μmol/(m2·s),AQY 降 低33.33%,Amax降低20.21 %,而Rd两者间差异不显著。

图2 全P 和缺P 条件下黑木相思叶片光响应曲线Fig. 2 Light response curves of A. melanoxylon seedlings leaves under normal P and P deficiency
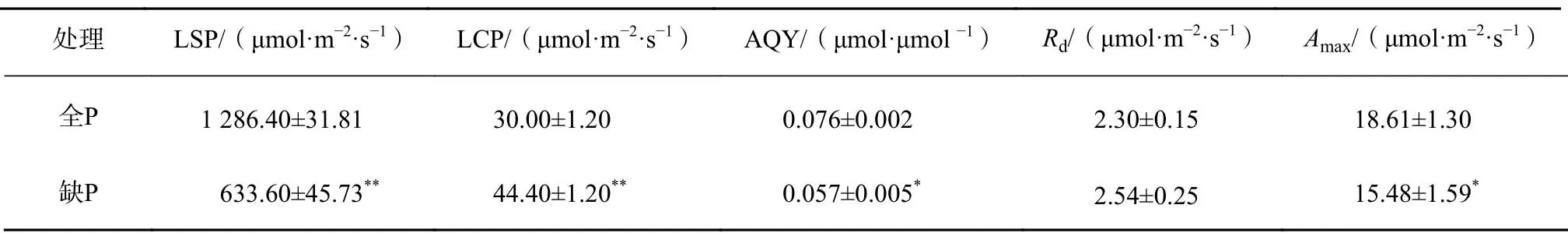
表4 全P 和缺P 条件下黑木相思叶片光合作用参数Table 4 Photosynthetic parameters of A. melanoxylon seedlings leaves under normal P and P deficiency
2.5 缺P 对黑木相思叶绿素含量的影响
由图3 可知,缺P 处理下,叶绿素a、b 含量及叶绿素总量显著低于全P 处理(P<0.05),其中叶绿素b 下降幅度达21.82%,叶绿素a 降幅为17.07%,叶绿素总量降幅为18.38%。

图3 全P 和缺P 条件下黑木相思苗木叶绿素含量Fig. 3 Chlorophyll content of A. melanoxylon seedlings leaves under normal P and P deficiency
2.6 缺P 对黑木相思各器官全N、全P、全K 含量的影响
由表5 可知,缺P 处理下,叶片中全N、全P 和全K 的含量极显著低于全P 处理(P<0.01),茎中全P 和全K 的含量极显著低于全P 处理(P<0.01),而全N 含量缺P 处理显著低于全P 处理(P<0.05),根中全N 含量在2个处理间差异不显著,而全P 和全K 存在极显著(P<0.01)和显著差异(P<0.05)。说明缺P 条件下除了根对N 的吸收不受影响外,根对P 和K 及茎、叶对N、P 和K 的吸收和储存都受到显著影响。其中影响最大的是根和茎中的全P 含量,缺P 处理比全P 分别降低了82.9%和86.8%,叶片中的全P 含量降低了59.7%,根、茎、叶中N 含量缺P 处理比全P 降低了9.3%~19.7%,而K 含量降低了14.7%~29.5%。

表5 全P 和缺P 条件下黑木相思苗木各器官养分含量Table 5 Nutrient content in different organs of A.melanoxylon seedlings under normal P and P deficiency mg/g
2.7 缺P 对黑木相思养分利用效率的影响
由表6 可知,缺P 条件下黑木相思各器官对N、P、K 养分的利用效率有显著差异,特别是P 的利用效率,根、茎、叶各器官在缺P 培养时都极显著高于全P 培养(P<0.01),其中根和茎的P 利用率分别提高了6.3 和6.7 倍,叶的P 利用率提高了2.5 倍。在N 和K 的利用率中,除根对N 的利用率在缺P 和全P 培养下无显著差异外,其他以缺P 培养显著(P<0.05)和极显著(P<0.01)高于全P 培养。说明缺P 条件下大大提高了黑木相思的N、P、K 的利用效率。
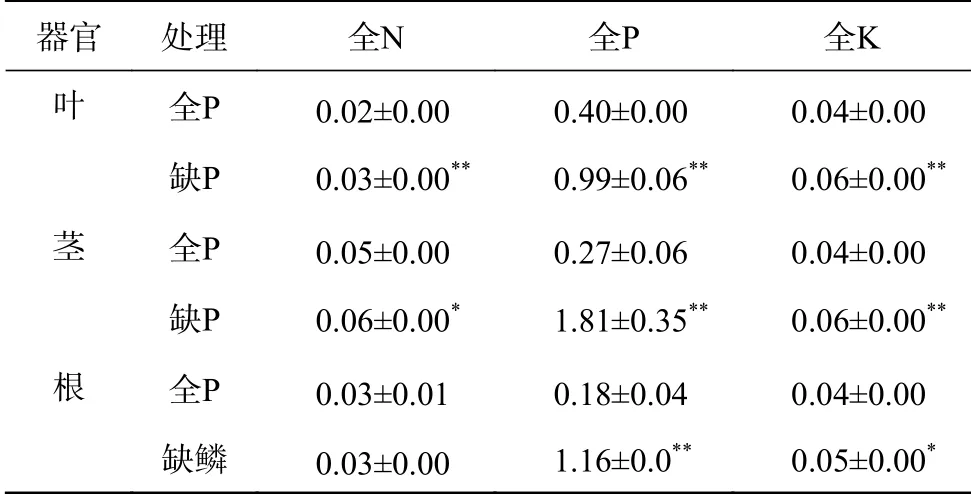
表6 全P 和缺P 条件下黑木相思苗木养分利用效率Table 6 Nutrient use efficiency of A. melanoxylon seedlings under normal P and P deficiency mg/g
3 结论和讨论
缺P 后,黑木相思苗木地上部生长受到显著限制,主要表现在株高降低,茎杆变细,叶量减少,而地下部总根长增加,根系表面积和体积保持稳定,表明黑木相思苗木通过减小地上部分生物量,保持地下部生物量,增加根系长度来适应缺磷环境,这些植株生长和根系形态的变化与大多 数 林 木 受 到 磷 胁 迫 的 形 态 变 化 相 似[5-6,11-12,18]。地上部生长量降低一方面可能是营养元素缺乏,体内代谢减缓所致,另一方面也是植株应对磷缺乏的形态适应机制,通过降低地上部生物量的分配及光合作用产物向根部的转移,促进地下部特别是细根生长,以扩大根系对磷的吸收。黑木相思苗木在缺P 下培养3个月,虽然表现出植株细弱,叶量减少的症状,但尚未出现叶脉紫红色,老叶有坏死斑块易脱落等缺素病症[19],说明黑木相思对磷缺乏具有一定的耐受力。
在顺境条件下,植物呼吸、光合作用及一些生化合成反应产生的自由基与清除自由基需要的氧化保护酶处于一种动态平衡状态,但在逆境条件下,自由基会加速产生,积累起来的活性氧就会对细胞产生伤害[20]。缺P 处理后,黑木相思苗木叶片和根系中3 种氧化保护酶(SOD、POD 和CAT)的活性同步升高,表明黑木相思通过提高各种氧化保护酶活性,来清除缺P 胁迫后苗木体内积累的活性氧,进而保护细胞免受其氧化损伤。另外,氧化保护酶活性受不同植株类型及磷胁迫程度影响,如巴西橡胶(Hevea brasiliensis)各种氧化保护酶活性随着磷胁迫时间的增加呈现持续增加或先升后降的趋势[7]。农作物中普遍认为保护酶系统活性变化幅度可衡量品种耐缺磷胁迫能力的强弱[21],例如,张玉霞等[22]对大豆(Glycine max)低磷胁迫研究结果表明氧化保护酶活性变化受基因型控制,磷高效基因型受胁迫后,氧化保护酶活性变化较大,磷低效基因型则变化不明显。本研究中黑木相思叶片中的POD 和CAT活性,缺P 处理比全P 增幅达113.7%和473.3%,反应了黑木相思耐低磷的生理适应机制。
缺P 处理下,黑木相思叶绿素含量,A等光合作用指标显著下降,这与马尾松(Pinus massoniana)的研究结果相同[23]。吴楚等[24]认为磷胁迫下水曲柳(Fraxinus mandshurica)叶片中可溶性蛋白下降导致1,5-二磷酸核酮糖羧化酶/加氧酶含量减少,从而引起光合作用速率的降低,而对叶绿素生物合成和含量影响不显著。Amax、Rd和AQY 也是反映植物光合能力的3个基础指标[25]。Amax反映了植物在强光下吸收和转换光能的能力,Rd代表了植物在黑暗下呼吸作用消耗的光合产物,AQY 反映了植物在弱光下吸收和转换光能的能力。黑木相思缺P 处理下Amax和AQY 显著下降,表明叶片捕获强光和弱光的能力都降低,从而限制光合速率,而Rd升高不显著,说明暂时没有对光合产物产生过量的消耗。
缺P 处理下,黑木相思苗木对N、P 和K 的吸收和存储显著降低,在根、茎、叶器官中,影响最大的为P 含量,降低了59.7%~87.0%,其次为K 含量,降低了14.7%~29.5%,N 含量降低了9.25%~19.7%。研究表明,马尾松在重度缺P 条件下,苗木对N 和K 的吸收量也显著减少,并且对N 的吸收影响要大于K[26]。缺P 处理下黑木相思根、茎、叶对P 元素分配表现为根和茎中P 含量大幅度减少,而叶中仍保含有一定量的P 含量,这可能是为了满足叶片进行一定的光合作用,维持正常的生理生化活动所需,也是黑木相思响应磷胁迫适应机制。在澳大利亚多种年代序列土壤中生长的植物研究表明,相思树(A.rostellifera)在极低磷土壤中仍保持叶总P 含量大于0.5 mg/L,这是相思树种适应低磷环境形成的一种叶P 组分分配模式[27]。缺P 条件下大大提高黑木相思根、茎、叶器官的养分利用效率,这与桉树(Eucalyptus robusta)在重度磷胁迫下,P 利用效率大幅增加相同[28]。研究表明磷胁迫下P 利用效率的提高与根际和叶片的酸性磷酸酶活性增强有关[29],叶P 功能组分研究表明,光合磷利用效率与代谢磷和结构性磷之比正相关[30]。
本文在表型和生理生化上探讨了黑木相思磷胁迫响应机制,事实上植物应答低磷环境引起体内转录因子、磷转运蛋白等大量基因响应,构成复杂的磷代谢调控网络[31]。如拟南芥(Arabidopsis thaliana)在磷胁迫下,占总基因29%的基因表达发生变化[32],1 000 多个功能基因被诱导和抑制[33],因此以后从分子生物学水平解析黑木相思磷胁迫的基因响应,将更清晰认识磷胁迫适应机制,为提高养分利用效率,实现高效栽培提供有效途径。
