吉林省典型生态区玉米种植土壤细菌多样性研究
2020-10-26王媛媛王继岩
王媛媛,王继岩,焉 莉,高 强
(吉林省商品粮基地土壤资源可持续利用重点实验室/ 吉林农业大学资源与环境学院,吉林 长春 130118)
吉林省地处松辽平原,属温带大陆性季风气候区,地势呈明显的东南高、西北低特征,四季分明,自然生态环境良好,是我国重要的黑土保护区。2018年吉林省玉米播种面积为423万hm2,占全省农作物总播种面积的69.6%,占全国玉米总播种面积的10.3%,是吉林省第一大粮食作物。根据气候条件、环境因子和地形地貌等方面的差异,将吉林省分为3个生态区,即东部湿润山区、中部半湿润平原区和西部半干旱平原区[1-2]。3个生态区的积温和总日照时数自东向西逐渐增加,而总降水量则逐渐降低。有研究证明,吉林省玉米产量为中部>东部>西部[3];氮肥利用率则呈现自东向西逐渐降低的趋势[4];钾肥利用率表现为西部>东部>中部[5];速效磷含量则自东向西逐渐下降[6]。可见吉林省玉米产量和肥料利用率存在显著的区域差异。前人的研究多集中于小尺度的田块作物产量、肥料利用率与土壤微生物的关系,但从生物地理学的角度来发掘和探索基于区域尺度的气候因素、土壤肥力特征与土壤微生物结构关系的研究相对较少。
土壤微生物对外界环境的细微变化非常敏感,细菌、真菌和放线菌是其重要组成部分,在降解有机质和促进营养循环等方面起积极作用[7],且共同维系着生态结构和功能稳定[8]。其中,细菌是土壤微生物中含量最多、丰富度最高的类群[9],环境因子的变化诸如地理因素、积温、气候变化、降雨量、理化性质、酶活性的差异性将引起土壤细菌群落种类和多样性的改变[10],从而直接影响土壤中养分物质的循环[11]。有研究证明,土壤pH值是影响细菌群落结构的主要驱动因子[12]。LIU等[13]对细菌群落结构的研究表明,细菌的群落结构组成和丰富度除了受pH值影响外,还与土壤有机质密切相关。王楠楠等[14-15]研究证实了降水量的变化能够明显驱动土壤微生物的群落结构和组成变化,不同降水条件下细菌的优势种群和丰度差异显著。CONG等[16]研究证明了土壤温度是影响微生物群落分布的主要因素,温度升高增强了土壤微生物的代谢活性,从而影响土壤微生物量及微生物群落结构和组成[17]。不同地区的气候条件差异可能导致环境因子对土壤微生物的影响程度不同,因此有必要分析何种环境因子对当地的土壤微生物群落特征有重要影响。
该研究以吉林省玉米种植土壤为研究对象,通过测定不同生态区玉米种植土壤的化学性质、酶活性及细菌微生物群落结构特征,分析环境因子对当地土壤细菌微生物群落结构的影响,系统研究气候、土壤化学性质、生物性质对土壤细菌的影响机制,分析土壤环境因子与土壤微生物之间的联系,寻找影响土壤微生物结构多样性的主要驱动因子,对改进吉林省玉米种植土壤的管理策略具有重要意义。
1 材料与方法
1.1 研究区介绍
土壤采样点分布于吉林省3个不同生态区,其气候特点见表1。东部湿润山区的年均温为4.20 ℃,年均降雨量为590~910 mm,有效积温为2 569.3 ℃,无霜期为115~170 d。采样位置主要包括延边的敦化和汪清、白山的江源和靖宇、吉林的桦甸和蛟河以及通化的辉南。暗棕壤和白浆土是东部生态区的主要土壤类型。

表1 吉林省不同生态区气候条件及土壤理化性质
中部半湿润平原区的年均温为5.4 ℃,年均降雨量为450~710 mm,有效积温为2 831.4 ℃,无霜期为125~160 d。采样位置主要包括吉林的磐石、舒兰和永吉,四平的公主岭、梨树、双辽、伊通,辽源的东辽和东丰,通化的梅河,长春的德惠、榆树和双阳。黑钙土、白浆土和黑土是中部生态区的主要土壤类型。
西部半干旱平原区的年均温为4.8 ℃,年均降雨量为378~494 mm,有效积温为2 940.2 ℃,无霜期为135~157 d。采样位置主要包括白城的大安、洮南、通榆、镇赉和松原的前郭、扶余、长岭。黑钙土、草甸土和风沙土是西部生态区的主要土壤类型。
1.2 土壤样品采集
于2018年6—7月对吉林省东部湿润山区、中部半湿润平原区和西部半干旱平原区3个生态区的玉米种植土壤样品进行采集,采样深度为0~20 cm,采用五点采样法共采集了72份土壤样品,每种土壤样品重复采集3次,结果取其3次重复的平均值,以确保实验的精确度。所有样品采集后立即过2 mm孔径筛,然后分成3份,1份于-80 ℃条件下保存用于高通量测序,1份存放在4 ℃冰箱用以测定土壤酶活性,1份土壤风干保存用以测定土壤理化性质。
1.3 理化性质、酶活性的测定与土壤测序
1.3.1理化性质测定方法
采用风干土壤样品进行理化性质分析,测定方法参照文献[18]。pH值采用电位法测定,水土质量比为2.5∶1;土壤有机质含量采用重铬酸钾容量法(外加热法)测定;碱解氮含量采用碱解扩散法测定;速效磷含量采用钼锑抗比色法 ( 0.5 mol·L-1碳酸氢钠浸提)测定;速效钾含量采用火焰光度计法(醋酸铵浸提)测定。
1.3.2酶活性测定方法
脲酶活性采用苯酚钠-次氯酸钠比色法测定[19],以24 h后1 g土壤中NH3-N含量表示。蔗糖酶活性采用3,5-二硝基水杨酸比色法测定[20]274-276,以24 h后1 g干土生成的葡萄糖量表示。磷酸酶活性采用磷酸苯二钠比色法(酸性磷酸酶用醋酸盐缓冲液,中性磷酸酶用柠檬酸盐缓冲液,碱性磷酸酶用硼酸盐缓冲液)测定[20]312-313,以24 h后1 g土壤中释放出的酚含量表示。每个样品均做无基质对照和无土对照,所有样品均做3次重复,以保证数据的准确度。
1.3.3土壤细菌16sDNA Amplicon高通量测序分析
使用土壤试剂盒(美国Omega),从0.5 g土壤中提取土壤总DNA,选用细菌16S rRNA基因的高度可变的V3-V4区用来测序,PCR扩增选用细菌16S rDNA V3-V4 区特异性引物,338F(5′-barcode+ACTCCTACGGGAGGCAGCA-3′),806R(5′-GGACTACHVGGGTWTCTAAT-3′)。PCR条件:98 ℃ 30 s;98 ℃ 15 s,50 ℃ 30 s,72 ℃ 30 s,25~27个循环;72 ℃ 5 min。扩增结果进行2%琼脂糖凝胶电泳检测,选取目标片段,用Axygen回收试剂盒回收PCR产物并纯化。最后,对合格的文库用Illumina技术进行测序。
1.4 数据分析
使用QIIME软件中的OTU表计算操作分类单元中的Chao1指数。使用R软件绘制维恩图和典范对应分析(CCA)图,分析样品中共有和独有OTU的数量关系以及环境因子对细菌菌群结构多样性的影响。利用SPSS 19.0和Excel 2010软件进行吉林省不同生态区土壤细菌丰度数据的单因素分析(ANOVA),用Duncan法分析不同生态区玉米种植土壤的细菌差异性。
2 结果与分析
2.1 吉林省不同生态区玉米种植土壤的理化性质
由表1可知,目前吉林省玉米种植土壤的pH平均值为6.58,有机质含量平均值为13.11 g·kg-1,碱解氮含量平均值为111.93 mg·kg-1,速效磷含量平均值为35.42 mg·kg-1,速效钾含量平均值为128.08 mg·kg-1。不同生态区的土壤理化性质差异明显。其中西部半干旱平原区的pH值较高,土壤偏碱性;中部半湿润平原区土壤呈中性;东部湿润山区pH值偏低,土壤呈酸性。东部湿润山区的土壤有机质含量最高,中部半湿润区次之,西部半干旱平原区土壤有机质含量最低。东部湿润山区土壤碱解氮和速效钾含量较高,中部半湿润平原区和西部半干旱平原区则较低,但2个区域无显著差异。综上所述,东部湿润山区的土壤肥力较高,中部半湿润平原区次之,西部半干旱平原区的土壤肥力较低。参考已有研究可知,中部地区的玉米产量较高,东部地区次之,西部产量最少[4-5]。这是土壤肥力和气候、地形地势等多因素综合作用的结果。
2.2 吉林省不同生态区玉米种植土壤的酶活性特征
由表2可知,玉米种植土壤的脲酶活性均值为36.58 mg·g-1·d-1,蔗糖酶活性均值为120.69 mg·g-1·d-1,碱性磷酸酶活性均值为0.88 mg·g-1·d-1,中性磷酸酶活性均值为2.68 mg·g-1·d-1,酸性磷酸酶活性均值为1.76 mg·g-1·d-1。脲酶和蔗糖酶均为土壤中重要的水解酶,可直接或间接反映土壤生物学活性强度。中部半湿润平原区的脲酶活性显著低于东部和西部地区(P<0.05),而东部湿润山区的蔗糖酶活性显著低于中部和西部地区(P<0.05)。东部湿润山区的酸性磷酸酶活性在3个生态区中最高,西部半干旱平原区的碱性磷酸酶活性显著高于其他2个生态区(P<0.05)。通过以上分析可知,偏酸性土壤的酸性磷酸酶活性较高,偏碱性土壤的碱性磷酸酶活性较高。
2.3 吉林省不同生态区玉米种植土壤细菌群落的Alpha多样性指数
由表1可知,中部半湿润平原区和西部半干旱平原区的Chao1指数平均值显著高于东部湿润山区,说明其具有较高的细菌菌群丰度。西部半干旱平原区的Chao1指数变异系数最小,说明西部半干旱平原区的样本相对中部和东部生态区来说更为均匀。
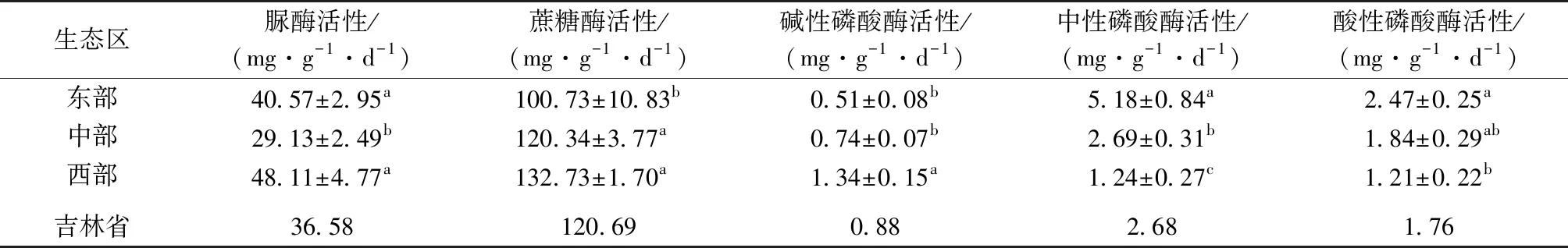
表2 吉林省不同生态区玉米土壤的酶活性
2.4 吉林省不同生态区玉米种植土壤的细菌群落结构组成
对所有样品进行16S rRNA测序,得到的细菌分类OTU共12 412个。其中,中部半湿润平原区细菌OTU数最多,共10 096个;其次是西部半干旱平原区,细菌OTU数为7 854个;而东部湿润山区细菌OTU数为6 585个。3个生态区共同包含的细菌OTU数有3 370个,占各个生态区的比例为33.38%~51.18%。各个生态区独有的细菌OTU数目及占比分别为中部1 514(18.05%)、东部1 418(11.04%)和西部727(15.00%)。这表明不同生态区间的细菌群落结构相似性较高,其中中部和西部群落共有OTU数较高,细菌群落结构较为相似;西部和东部群落共有OTU数相对较少,细菌群落结构存在部分差异。
土壤细菌在门、属分类水平上的相对丰度如图1所示。将3个生态区所有样品进行整体分析发现,在门水平上放线菌门(Actinobacteria,占比19.14%~45.64%)为相对丰度最高的优势菌门,其次是变形菌门(Proteobacteria,21.65%~35.22%)、绿弯菌门(Chloroflexi,11.56%~17.67%)、酸杆菌门(Acidobacteria,5.91%~14.66%)、芽单胞菌门(Gemmatimonadetes,5.43%~10.49%)。从3个生态区分布看,放线菌门呈现自西向东逐渐减少的趋势,而变形菌门、酸杆菌门、拟杆菌门的相对丰度与之相反,呈现自东向西逐渐减少的趋势,西部半干旱平原区的绿弯菌门含量最高,中部半湿润平原区的芽单胞菌门含量最高。
属水平上真菌丰度较高的有芽单胞菌属(Gemmatimonas,1.35%~3.93%)、红色杆菌属(Rubrobacter,0.55%~4.86%)、鞘脂单胞菌属(Sphingomonas,1.32%~2.38%)、慢生根瘤菌属(Bradyrhizobium,1.28%~2.49%)和苍白杆菌属(Ochrobactrum,0.05%~6.98%)。吉林省3个生态区玉米种植土壤细菌群落中,芽单胞菌属在中部半湿润平原区占比最高,其次是东部半湿润山区和西部半干旱平原区,苍白杆菌属、慢生根瘤菌属呈现自东向西逐渐降低的趋势。排名前20的细菌菌属一致,但含量有细微差别。

图1 吉林省各生态区细菌门及属水平的相对丰度Fig.1 Relative abundance of bacterial at phylum and genus levels in various ecological regions of Jilin Province
2.5 土壤细菌群落与环境因子的CCA分析
该研究采用CCA分析法探究影响吉林省各生态区细菌群落组成的环境因子,更加直观、清晰地体现出土壤理化特性及气候因素对土壤细菌群落多样性及优势类群的影响。如图2所示,环境因子中pH值的射线最长,其次为射线碱解氮和有机质含量,因此,pH值、碱解氮含量、有机质含量对细菌微生物群落结构影响程度最高,是土壤细菌优势类群变化的主要驱动因子;其次是降雨量和纬度,可见气候因子中降雨量和纬度对细菌群落结构影响也较大。酸性磷酸酶活性、年均降雨量、中性磷酸酶夹角为锐角;pH值、积温、脲酶活性、碱性磷酸酶活性、蔗糖酶活性之间夹角均为锐角,表明这几种环境因子之间存在正相关关系,且可能具有一定的协同效应。从吉林省不同生态区细菌分布来看,大部分西部半干旱平原区的土壤样品分布在CCA横坐标轴的负方向,东部湿润山区的土壤样品分布在CCA横坐标轴的正方向;而中部半湿润平原区的土壤样品分布主要集中在坐标轴附近。西部和东部生态区样本分布距离相对较远,说明2个生态区中的土壤微生物细菌群落结构有各自独特的特点。

样品编号首字母W代表西部地区;首字母C代表中部地区; 首字母E代表东部地区样品。图2 细菌群落与环境因子的典范对应分析Fig.2 CCA analysis of bacterial community and environmental factors
通过R软件利用偏最小二乘回归分析,对物种丰度矩阵和样本分组数据构建PLS-DA判别模型(图3),也能发现吉林省3个生态区的土壤微生物细菌群落特征存在很多的共性,差异较小。
3 讨论
3.1 降雨量和纬度对土壤微生物的影响
气候条件和地理因素等自然条件是导致不同生态区土壤性质存在差异的主要原因之一。研究区位于温带大陆性季风气候区,因此土壤细菌微生物群落存在很多共性,但3个生态区存在一定的地域差异,任何环境变化都会对土壤微生物群落结构多样性有一定影响[21]。笔者研究表明气候条件中降雨量和纬度对土壤细菌微生物构成有显著影响。已有研究表明降雨可改善土壤水分状况,提高土壤微生物活性,甚至改变土壤微生物群落结构[22]。不同降雨条件下,真菌和细菌的优势种群和丰度差异显著[14-15]。也有研究表明,随着降雨量升高,土壤含水量升高,土壤微生物呼吸率变大[23]。土壤水分过多或缺乏,均会使土壤微生物量及其活性降低,微生物量周转变慢,而适宜的土壤水分含量有利于微生物生长繁殖,进而导致土壤微生物量升高[24]。全球变暖必然导致气候变化,其中降雨变化尤为突出,可能对土壤微生物多样性和群落结构产生深刻影响。纬度变化对土壤微生物的影响目前还没有明确。但有研究发现土壤有机碳、总氮含量均随纬度的升高而指数型减小[25];李喜霞等[26]的结论却相反,认为土壤有机碳含量、C/N比和C/P比随纬度升高而显著增加,这可能是因为2个研究点的环境差异明显。该研究同时发现纬度与有机质含量有协同效应。

样品编号首字母W代表西部地区;首字母C代表中部地区; 首字母E代表东部地区样品。图3 不同生态区细菌群落的PLS-DA分析Fig.3 PLS-DA analysis of bacterial communities in different ecological regions
3.2 pH值、碱解氮和有机质含量对细菌微生物的影响
土壤环境、动植物和微生物的生存条件由土壤理化性质决定。吉林省3个不同生态区玉米种植土壤的pH值存在显著差异,主要呈现自西向东递减的趋势;土壤肥力则从东向西逐渐递减。研究结果发现pH值对细菌微生物群落结构有重要影响。有研究表明土壤pH值能提高土壤中细菌数量[27]。这是因为细菌喜碱性环境生长,土壤中的微碱性环境更有利于细菌的活动和繁殖。笔者研究也发现中、西部地区土壤细菌的群落丰度和群落多样性高于东部地区。
除了自然因素外,人为因素对土壤微生物群落结构产生了更严重的影响。GUO等[28]的研究结果发现近20 a间我国耕地土壤酸化严重,pH值降低明显。大量文献证实土壤pH值会影响局部[29]、区域[30]和全球范围[31]的细菌群落组成和多样性,且过量、不合理使用氮肥等会导致土壤酶活性和土壤pH值降低,从而抑制土壤细菌生长[32]。该研究发现吉林省玉米种植区土壤中放线菌门占比随pH值降低而明显降低,放线菌主要能促使土壤中的动物和植物遗骸腐烂,可见未来土壤微生物的降解和分解能力也许会受影响。同时研究还发现碱解氮和有机质含量对该区细菌微生物群落结构也有显著影响,这与SESSITSCH等[33]和LIU等[13]的研究结果一致。可能土壤全氮和全碳含量增加为细菌微生物的生长提供了充足的能源和碳源,促进了细菌数量的增加。
4 结论
吉林省不同生态区玉米种植土壤肥力呈现自东向西逐渐降低的趋势,中部半湿润平原区的脲酶活性最低,蔗糖酶活性和碱性磷酸酶活性呈现自西向东逐渐降低的趋势,中性磷酸酶和酸性磷酸酶活性呈现自西向东逐渐增加的趋势。中部半湿润平原区和西部半干旱平原区的细菌多样性显著高于东部湿润山区,细菌菌群丰度呈现自西向东逐渐降低的趋势。3个生态区的优势细菌门均为变形菌和放线菌,而芽单胞菌和苍白杆菌则是优势菌属。放线菌门呈现自东向西逐渐增加的趋势,变形菌门含量变化与之相反。芽单胞菌属呈现中部半湿润平原区>东部半湿润山区>西部半干旱平原区的趋势,苍白杆菌属呈现自东向西逐渐降低的趋势。典范分析结果表明,气候影响因素中降雨量和纬度对细菌微生物群落组成影响较大,pH值、碱解氮和有机质含量是土壤细菌优势类群变化的主要驱动因子。
