甘蓝型油菜小孢子培养中基因型及处理条件对成苗的影响
2020-10-23皮明雪郭少敏杨雯竹张敏琴吴江发杜才富
皮明雪,郭少敏,张 涛,杨雯竹,张敏琴,吴江发,杜才富
(1.贵州省农业科学院油菜研究所,贵阳 550008; 2.安顺学院,贵州 安顺 561000)
小孢子培养是通过培养游离花粉小孢子,使得花粉小孢子进行胚胎发育生成新个体的过程。可以快速纯化遗传背景复杂的材料,获得纯合且稳定的双单倍体(Double haploid,DH)再生植株,可缩短育种周期,提高育种效率,在QTL定位、遗传育种研究和诱变育种等方面具有广阔的应用前景[1-6]。由于条件限制,国内小孢子培养的供体材料多生长于大田,小孢子发育在花期容易受外界环境扰动的影响;吴士红等研究认为,在高海拔地区的出胚率比在低海拔的出胚率高[7]。高海拔地区的天气在油菜开花期相对稳定,促进了小孢子同步发育。赵小萍等认为,供体植株在人工气候室生长条件下的小孢子培养出胚效果比大田和温室的好[8],人工气候室的光照、温度和湿度均可自动化控制,可免供体植株受外界环境冲击,利于得到发育同步性高的小孢子。小孢子再生苗诱导培养时通常15 d左右需要换一次培养基进行继代培养,长出再生植株需继代2~3次,为减少时间和经济成本,应提高胚状体一次成苗率。目前,在实际应用中存在再生苗加倍率较低的问题,并且由于常用的秋水仙素毒性较大,单纯的加大秋水仙素的用量和延长处理时间不能很好的解决问题,这大大降低了小孢子培养技术的应用价值。因此,提高出胚率、小孢子胚一次成苗率和加倍率一直是小孢子培养技术的重要研究点。本试验研究了不同基因型对小孢子培养胚产量和再生植株自然加倍率的影响,以及小孢子振荡培养过程中光照条件与后期低温处理对胚状体一次成苗率和再生植株自然加倍率的影响,以期提高油菜小孢子培养效率,优化油菜小孢子培养、再生和加倍技术体系;并获得了一个胚产量和自然加倍率都很高的甘蓝型油菜DH系。
1 材料和方法
1.1 供体材料
试验材料为7个甘蓝型油菜F1杂交种和1个DH系(表1),F1亲本均为DH系。18 G 309和D 256为经过检验的出胚较高的DH单株系,Q 219B系列,5365系列来自2018年育种亲本纯化得到的DH株系,育种亲本出胚较好。F1杂交都是在5365系列和Q 219 B系列的DH单株开花后进行的,F1杂种和D 256于2019年6月上旬种植在贵州省油菜研究所的人工气候室(25~16 ℃),每个材料选用8株供体植株,于2019年8月中旬至9月中旬取材,供体生长期间控制好水肥。
1.2 方 法
1.2.1小孢子的提取与培养
选择在08:00—10:00时取材,选取健康植株的主花序和生长状态较好的分支花序,用冰盒带回实验室备用。小孢子的提取与培养方法参照余凤群等[9]的方法。
1.2.2振荡培养光照条件和低温处理
25 ℃恒温培养至肉眼可见胚出现时,将培养皿移至50 r·min-1、25 ℃恒温摇床上振荡培养,并进行光照培养(16 h光/8 h暗,光强2 000 lx)和暗培养2个处理,7 d后统计胚产量(胚·蕾-1),并将出胚较好的2个基因型材料的子叶胚置于4 ℃条件下,分别进行0、5、10、15、20、25 d的低温处理。
1.2.3一次成苗率和自然加倍率的统计
将低温处理过的子叶型胚接种在琼脂浓度为8 g·L-1的B 5固体培养基上,于25 ℃,16 h光/8 h暗,15 000 lx光照强度条件下培养,15 d后统计一次成苗率(长出3~5片叶子并形成根系计为成苗)。将成苗植株取出,洗去培养基后移栽到湿润的土壤中,盖上薄膜保湿,置于人工气候室中练苗,7 d后揭开薄膜,14 d后移栽至地里(期间每天适当浇水),在盛花期和结果期,参照程利华[10]的方法统计自然加倍率,根据花的大小、花药发育情况、成熟花粉粒量以及角果形态,将植株分为自然加倍型和未加倍型。
2 结 果
2.1 基因型和母本效应对甘蓝型油菜小孢子培养胚产量的影响
在本试验中,对8份甘蓝型油菜进行小孢子培养,统计胚产量。5365-19新9和5365-21新6源于同一个育种材料背景5365的DH株系后代,胞质来源相同,基因型高度相似;Q 219 B-1和Q 219 B-3与之情况相同。结果如表1所示,材料间存在显著差异,2个F1代杂交种D 393和D 524的胚产量很高,分别为119.87胚·蕾-1和98.48胚·蕾-1;DH系D 256次之,胚产量为68.51胚·蕾-1;其它几个材料的胚状体产量则较少,尤其D 485没有获得胚状体。D 284和D 485的亲本基因型相似,胚产量结果相差不大,D 381和D 407也如此;而D 393和D 407的父本相同,母本不同,胚产量结果却存在显著差异,分别为119.87胚·蕾-1和23.10胚·蕾-1;D 393和D 372共用了DH亲本18 G 410,其余亲本5365-19新9和5365-21新6基因型高度相似,胚产量分别为119.87胚·蕾-1和13.72胚·蕾-1,差异显著。结果表明,基因型是影响甘蓝型油菜小孢子培养胚产量的重要因素。
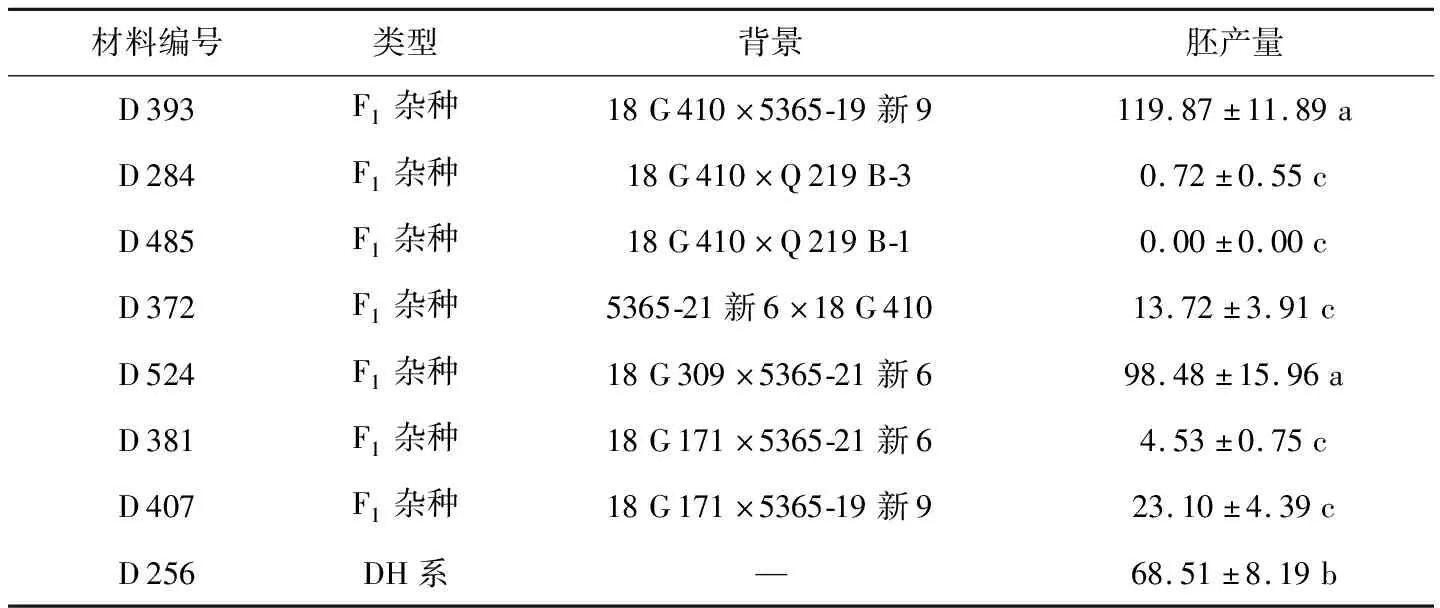
表1 不同基因型甘蓝型油菜的小孢子胚产量结果
2.2 振荡培养光照条件和低温处理对甘蓝型油菜小孢子培养一次成苗率的影响
在天然状态下,从完成受精作用形成种子胚到种子发育成熟的过程中,油菜种胚也可以进行光合作用为自身提供营养。小孢子胚打破了花粉的发育进程成为胚状体,在培养过程中可以进行光合作用,为探究光照是否可以促进胚状体成熟,故而设计了培养过程中的光照/黑暗培养对比。
因D 393和D 524在振荡光照培养与黑暗培养条件下出胚效果较好,故选为试验材料,分别于4 ℃低温条件下处理0、5、10、15、20、25 d后,将子叶型胚培养于B 5固体培养基上,半个月后统计一次成苗率。结果如表2所示,D 393的一次成苗率在振荡光照培养与黑暗培养条件下没有显著差异,但随着低温处理时间的延长呈先升高后降低的趋势;D 524同样如此。在光照培养条件下,D 393在低温处理15 d后的一次成苗率最高,为54.95%,暗培养条件下同样低温处理15 d的一次成苗率最高,为61.48%,均显著高于对照;在光照和暗培养条件下,D 524的一次成苗率均是在低温处理15 d后最高,分别为44.07%和50.28%,显著高于对照。因此,甘蓝型油菜小孢子培养的一次成苗率与振荡培养的光照条件关系不大,但受到低温处理时间的显著影响,尤其4 ℃低温处理15 d能提高小孢子一次成苗率0.8~3倍。
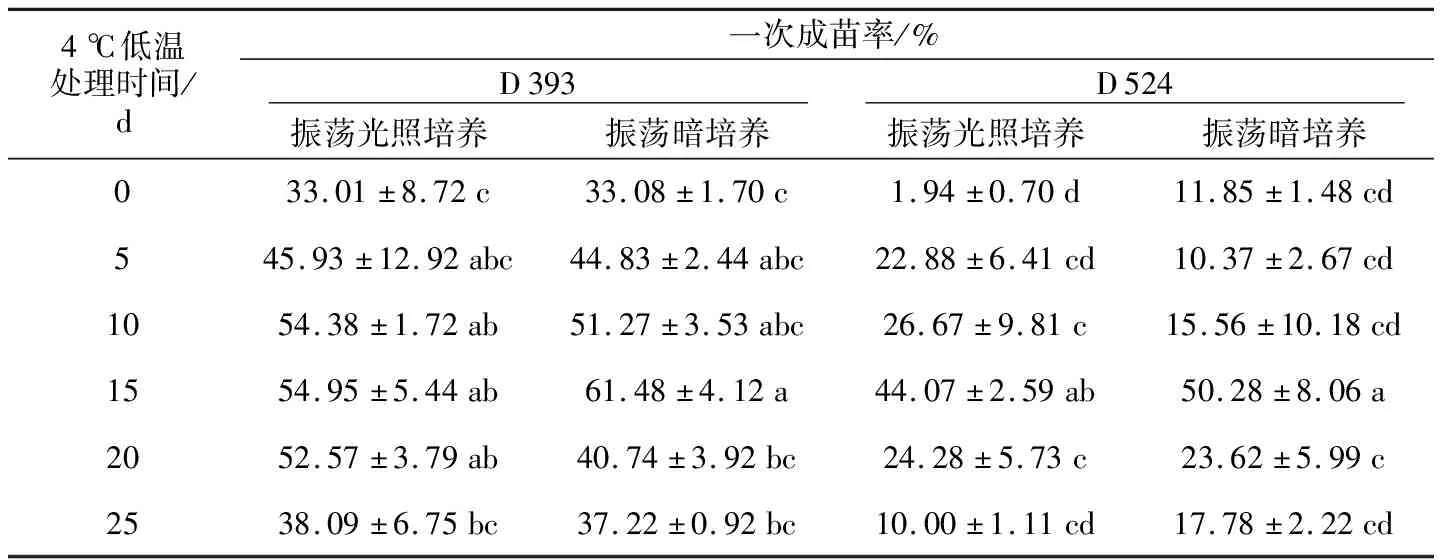
表2 振荡培养光照条件和低温处理对甘蓝型油菜小孢子培养一次成苗率的影响
2.3 基因型、光照条件和低温处理对甘蓝型油菜小孢子培养自然加倍率的影响
在盛花期和结果期,分别观察经过振荡光照培养与黑暗培养及各低温处理的D 393和D 524小孢子再生苗的田间表型,将花序轴较粗壮、花蕾大能正常开花、花大小同正常二倍体、花药发育完全、成熟花粉量大、能正常散粉、角果长而饱满的植株计为加倍型;花序轴细、花蕾较小、大部分花未开便脱落、部分能开花、花也较小、多数花药强烈退化、有些花药小且空瘪不能散粉、结成短小萝卜状角果的植株计为未加倍型。如表3所示,在光照培养下,D 393的自然加倍率与暗培养的没有明显差异,各个低温处理时间下的加倍率差异也不显著;D 524也是如此。D 393的自然加倍率在73.52%~85.05%之间,均值为79.87%;D 524的自然加倍率在21.49%~33.41%之间,平均为28.00%;D 393的平均加倍率是D 524的2.85倍。结果表明,自然加倍率没有受到振荡培养过程中的光照以及低温的影响,而基因型对小孢子再生植株的自然加倍率影响较大。
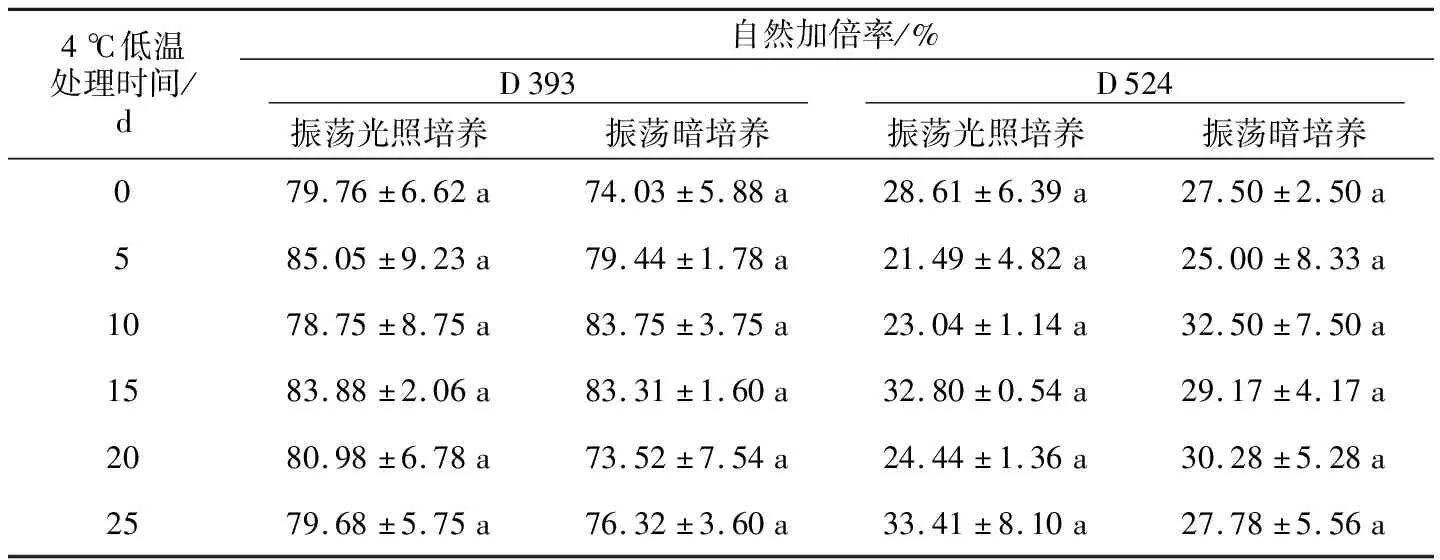
表3 基因型、光照条件和低温处理对甘蓝型油菜小孢子培养自然加倍率的影响
3 讨 论
3.1 基因型影响胚产量
供体材料的基因型是影响甘蓝型油菜小孢子培养胚产量的重要因素[11-13]。统计了8个不同基因型甘蓝型油菜材料的胚产量,其中只有D 393和D 524这2个F1代杂交种和DH系D 256的胚产量较高,每蕾分别高达119.87、98.48胚和68.51胚,其它几个材料则胚状体产量较少,甚至有的为0,差异极显著,表明基因型对甘蓝型油菜小孢子培养胚产量的影响很大。王汉中等[12]对42个不同基因型材料进行小孢子培养,结果不同材料的平均胚产量变幅为每蕾0~11.67胚;在李书宇[13]的研究中,64份不同基因型材料的胚产量也存在较大差异,每蕾变幅为0~31.23胚。本研究中最高胚产量比以上研究高出很多,除了基因型的原因,可能还与供体植株种植在人工气候室,小孢子发育同步性高有关[8]。
本研究发现,基因型的影响中,母本对于小孢子培养的出胚效果影响更大。在本研究中,具有相同母本的材料出胚率不同,D 393、D 284和D 485;D 381和D 407之间差异显著;说明相同的母本组合中,母本对父本的响应有所不同。相同的父本组合材料之间,D 393和D 407的差异为5.2倍;D 524和D 381之间的差异达20.86倍;因此相同父本情况下,母本材料选择的不同,对出胚率可能有巨大的影响,但还需进一步的实验进行证实。刘泽等[14]研究表明,出胚率高的亲本杂交,其F1代一般具有较高的出胚率;2个出胚率低的亲本杂交,所得F1杂种的出胚率并非最低,但是高产胚率亲本与低产胚率亲本杂交后,改良F1杂种胚产量的效果并不明显。本研究的结果与之类似,但是母本的选择有望使得油菜小孢子培养选择时,获得更高的出胚率。
3.2 一次成苗率提高的可能性
低温处理对甘蓝型油菜小孢子胚状体的一次成苗率有显著影响。本研究探究了甘蓝型油菜小孢子培养中振荡培养的光照条件和低温处理时间对胚状体一次成苗率的影响,结果发现,一次成苗率没有受到振荡培养光照条件的影响,但随着4 ℃低温处理时间的延长呈先升高后降低的趋势,在低温处理15 d时最高。研究表明,2 ℃低温处理的效果比4 ℃好,10 ℃对成苗率影响不大[15,16]。低温处理时期(胚状体转接到B 5固体培养基之前或之后)对胚状体成苗率的影响也不明显,但是后者所占空间较大,不便操作[17]。低温处理对小孢子胚状体再生率的影响主要在于低温处理时间,研究认为,低温处理4 d最佳,也有人认为最佳低温处理时间为10 d和14 d[16-18]。不同研究者得到的最佳低温处理时间有所不同,可能与供体材料的基因型及选取时间段不同有关。但总的来说,4 ℃低温处理10~15 d能显著提高甘蓝型油菜小孢子胚状体的一次成苗率。Dias研究发现,川贝母胚状体内的过氧化物酶活性和表达量受到了低温处理的诱导上调,因此他认为过氧化物酶是川贝母小孢子胚状体成苗的重要因素[19];万丽丽等认为,低温处理显著提高小孢子胚状体再生能力的原因可能是低温处理引起了小孢子再生过程中超氧化物酶活性的下降,极大加快了呼吸速率与热量的产生,使次生代谢物质产生减少,从而避免了细胞脱分化[18]。在甘蓝型油菜中,低温处理能提高小孢子胚状体一次成苗率的原因是否与过氧化物酶有关有待进一步探究。
3.3 自然加倍率的提高
理论上,小孢子只携带了供体植株的一半染色体,由小孢子得到的再生植株应为单倍体,但是实际上不同植物小孢子再生植株都出现了不同比例的二倍体化。甘蓝型油菜的小孢子植株自然加倍率普遍较低。本研究中,D 393的自然加倍率在73.52%~85.05%之间,D 524的为21.49%~33.41%,D 393的加倍率是D 524的2~3倍。在程利华[10]的研究中,7份甘蓝型油菜中有3个材料的植株中发生了自然加倍,最高的加倍率达到94.33%,其余2个仅有7.69%和4.55%,而其它4个材料的则为0。结果表明,基因型对小孢子再生植株的自然加倍率影响较大。研究发现,小麦的小孢子再生植株自然加倍率受到低温预处理的影响[20,21]。在本研究中甘蓝型油菜D 393和D 524植株的自然加倍率没有受到振荡培养过程中光照条件以及后期低温处理的影响,至于会不会受到低温预处理的影响有待进一步研究。大白菜小孢子再生植株的自然加倍率普遍较高,比甘蓝型油菜和甘蓝都要高,但是受基因型影响[22]。多数白菜材料自然加倍率可达50%以上,还有不少材料高达70%以上[23,24]。李菲等[25]探讨了大白菜小孢子培养自然加倍的机理,认为小孢子培养的自然加倍发生在胚诱导初期,大白菜小孢子再生植株较高的自然加倍率可能与小孢子培养中热激诱导激发了单倍体小孢子核的加倍有关。激素处理或其他诱导方法也一样能导致单核小孢子加倍[26]。目前还没有关于甘蓝型油菜小孢子培养自然加倍机理的相关报道,D 393的胚产量和自然加倍率很高,具有研究意义。
