中国东部暖温带刺槐物候模型比较*
2020-10-23于裴洋同小娟张静茹刘沛荣
于裴洋,同小娟**,李 俊,张静茹,刘沛荣
中国东部暖温带刺槐物候模型比较*
于裴洋1,同小娟1**,李 俊2,张静茹1,刘沛荣1
(1.北京林业大学生态与自然保护学院,北京 100083;2.中国科学院地理科学与资源研究所陆地水循环及地表过程重点实验室,北京 100101)
植被物候是陆地生态系统对气候变化响应的一个有力指标,其对水、碳交换和能量平衡发挥着重要作用。在全球气候变暖背景下,植被物候变化规律及预测植物物候期成为研究热点。本研究基于中国东部暖温带10个观测点的气象数据和刺槐地面物候观测资料,利用模拟退火算法对SW、Unichill和DNGDD三种模型的各项参数进行优化,对刺槐春秋季物候期(叶芽开放期、展叶始期、开花始期和叶变色期)进行模拟,通过内部检验对比分析SW、Unichill和DNGDD模型对春、秋季物候期的模拟效果,以选择出最适合预测刺槐物候期的模型。结果表明,刺槐4个主要物候期与同期各项气温平均值间均呈极显著负相关关系。利用模拟退火法对SW、Unichill和DNGDD模型进行参数估计得到的数值符合刺槐的生长发育规律。与DNGDD模型和Unichill模型相比,SW模型对春季物候期模拟效果较好,其模拟的叶芽开放期、展叶始期和开花始期对应的交叉检验方差解释量R2分别为0.807、0.876和0.907,均方根误差RMSE为6.0、4.6和4.4d。DNGDD模型则对秋季物候(叶变色期)模拟效果较好,其模拟的叶变色期的交叉检验方差解释量R2为0.580,RMSE为13.4d。因此可以得出,SW模型适合对刺槐春季物候(叶芽开放期、展叶始期、开花始期)的模拟,DNGDD模型则较适用于模拟秋季物候(叶变色期)。
刺槐;物候模型;春季物候;秋季物候;积温;退火算法
植物物候对气候变化十分敏感。气候变化从多方面影响植物物候,如物种分布[1-2]、种群的动态[3]和生命史进化[4]等。IPCC第五次报告指出,1901–2012年全球平均气温上升了0.89℃[5]。气候变暖造成全球多数地区植物叶芽开放、展叶、开花等春季物候期提前[6-8]。因此,开展温带阔叶林区树木物候期的研究,对了解气候变化与物候关系具有重要意义。
温带树木物候主要受气温影响[9]。朱媛君等[10]对呼和浩特近年物候研究发现,气温是影响该地区物候的主要因素,刺槐盛花期与其前3个月平均气温呈显著负相关关系。王明等[11]研究了暖温带乔木和灌木对气候变化的响应,结果表明乔灌木的各物候期与气温表现为负相关。刘璐等[12]研究中国北方地区的苹果始花期变化,明确了影响苹果始花期的气候要素为平均气温和平均地温。
为准确预测植物物候如开花期、变色期,国内外许多学者发展了不同物候模型。物候模型可以分为静态模型和动态模型两大类[13]。静态模型是基于数学关系建立的模型,如统计模型。动态模型包括SW模型、Unichill模型等,其利用模型对生物机制进行再现和模拟,这种模型相对于静态模型更为准确[14]。王焕炯等[15]利用Unichill模型对中国白蜡展叶始期进行模拟表明,近60a中国大部分地区的白蜡树展叶始期提前,且北方地区提前趋势更为明显。柏秦凤等[16]研究发现,中国富士系苹果花期由南向北逐渐推迟。李荣平等[17]采用模拟退火算法积温模型对开花期进行模拟,得到紫丁香开花始期和开花盛期>5℃·d积温具有较好的稳定性,能够利用积温模型进行模拟。温带地区植物的叶变色期有推迟的趋势,这可能是夏末秋初的温度升高导致的[18],Park等[19]使用GDD模型对韩国两种温带落叶树种叶变色期对气候的响应研究发现,两种树种叶变色期与该物候期前两个月的平均气温密切相关。汪箫悦[20]使用阈值模型、传统GDD和DNGDD模型对秋季物候进行预测,DNGDD模型的相关性明显优于其余两种模型,在中国北部和西南部地区,DNGDD模型预测的秋季物候期则晚于GDD模型模拟值。
目前,国内外对于物候模拟的研究已经取得了很多成果,但是由于植物生长发育机理的复杂性,仍然存在许多问题。多数物候模型对于春季物候的模拟效果较好。然而,对于秋季物候模拟则并不理想且研究较少。因此,不能仅简单地利用平均气温对秋季物候进行模拟,应考虑更多的气象因素[21],不断改进模型以使其能在大尺度范围内也同样适用。
刺槐为温带树种落叶乔木,豆科刺槐属,在年均温8~l4℃、年降水量500~900mm的地区生长良好,因其速生性强,且易繁殖,抗逆性优良,被世界各地广泛引种[22]。中国大部分地区的气候条件均能满足刺槐的生长需求,这使得其分布范围较广。刺槐是中国研究最全面最深入的树种之一,是温带落叶乔木的典型树种之一。本研究基于中国东部暖温带10个观测点的刺槐物候数据和气象数据,探究刺槐物候期与气温的相关性;采用模拟退火算法,比较了SW、Unchill和DNDGG三种动态模型模拟值和实测值,对各模型进行了内部检验和交叉检验,筛选和优化模型,以期选择出能准确模拟该区域刺槐4个主要物候期即叶芽开放期、展叶始期、开花始期和叶变色期的模型,为林业应对气候变化研究提供技术支撑,并为模型的优化提供研究思路。
1 资料与方法
1.1 物候数据
研究所选代表树种为刺槐(L.),主要对其叶芽开放期、展叶始期、开花始期和叶变色期进行模拟研究。物候数据来自中国物候观测网(http://www.cpon.ac.cn/),该网数据符合物候观测标准[23]。共选取10个站点的物候数据,由于自然气候和虫害等因素导致物候数据部分缺失。各站点的物候数据观测时段见表1。

表1 10个物候观测站资料信息
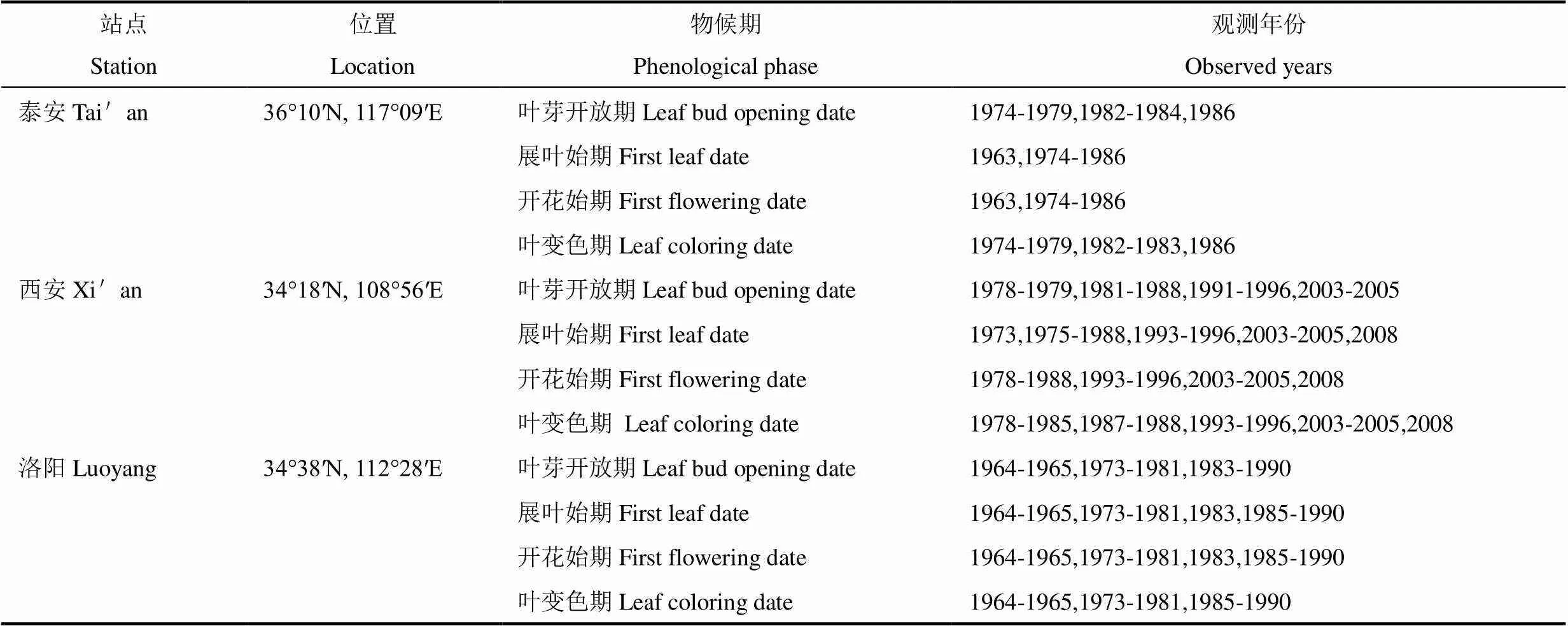
续表
1.2 气候数据
气候数据来源于中国气象数据网(http://data. cma.cn/site/index.html),包括10个物候站点对应的气象站点的日平均气温、日最高气温和日最低气温。在选取气象站点时依据物候观测站点选取与之对应的气象站点,其中除盖州站外其余物候站点均有与之地理位置相同的气象站点,则直接使用这些站点的气象数据。盖州无与之地理位置相同的气象站点,以地理位置相近的营口站替代。
1.3 物候模拟模型
(1)SW模型
SW模型(即热时模型)从积温理论发展而来,只考虑了温度的热积累[24-26],包括t0、Tb和F*三个参数。该模型假设能量与植被生长之间存在线性关系,当达到一定量的蓄热单位后能量就会爆发,此时即为植物生长的开始。SW模型在模拟春季物候期中被广泛应用。北方地区刺槐物候期的主要影响因素为春季积温,因此,使用SW模型能取得较好的模拟效果。SW模型表示为
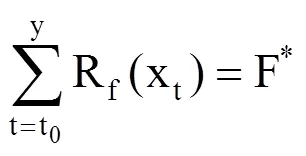
其中
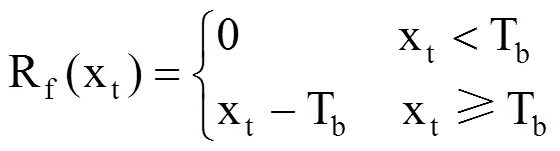
式中,Rf(xt)为强迫速率函数,表示逐日平均气温高于某一界限的值[27]。xt为第t天(DOY,以1月1日为1)的日平均气温(℃),Tb为界限温度(℃),需要根据不同植物各类确定,t0为累积起始时间(设置为每年的1月1日,即t0=1),以日序表示。F*指完成某物候发育所需的积温阈值(℃·d),利用日平均气温和实测物候资料,采用模拟退火法求解得到。利用式(1),当逐日高于某一界限值的气温累加值达到界限值F*时,对应的日期即为模拟预测的植物物候期日序(y)。
(2)Unichill模型
Unichill模型即统一冷激模型,不仅考虑了生长期强迫温度的影响,还考虑了休眠期低温。该模型把植物发芽阶段细化为休眠期和静止期两个阶段。休眠期是指植物在外界条件良好的情况下,由于某些内部因素阻止发育和细胞生长的时期[28];静止期是指休眠结束后生长发育开始受到外界因素驱动的一段时间,当经历外界低温持续刺激才能结束休眠进入静止期。该模型假定,每年9月1日后,虽然外界环境条件适合但由于内部因素阻止了树木的生长发育和细胞生长,树木生长基本停止,随后进入秋冬季节,气温降低,外界条件不适合树木生长,中国北方刺槐一直处于休眠期;并不断接受外界冷激刺激,当积累到冷激阈值后休眠期结束进入静止期,随着外界气温升高,刺槐会开始生长发育,随后生长发育速度则开始受外界气温主导。Unichill模型为


其中

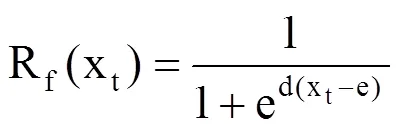
式中,参数a、b、c、C*与冷激函数有关;Rc(xt)为冷激速率函数(即冷激单元),表明冷激速度对日平均气温(℃)的响应;Sc是冷激单元之和,表示芽目前的冷激速率之和;t0设置为固定日期,为前一年的9月1日;t1为静止期,与SW模型不同,t1不是固定日期而是变化的,当逐日低温积累达到阈值C*时(℃·d),对应的日期为静止期日序t1。Rf(xt)为强迫速率函数(即强迫单元),参数d、e确定了静止期发育速度与日平均气温(℃)的函数关系,输入日平均气温与实测数据使用模拟退火法求解参数值。Sf是强迫单元之和,表示强迫速率的日总和,利用式(4),当强迫单元之和达到阈值F*时,tb为对应物候期的日序。
冷激单元和强迫单元均为日平均气温(xt)的函数,其中参数较多,有a、b、c、d、e、C*、F*七个参数,这些参数需要利用日平均气温和实测物候资料,采用模拟退火法求解得到。
(3)DNGDD模型
DNGDD模型即基于昼夜气温的积温模型。中国北方地区昼夜温差较大,白天和夜间气温对刺槐生长发育的速率影响不同,DNGDD模型结合中国北方地区的气候特点把平均气温用白天气温和夜间气温替代[29],并进行加权平均,权重系数分别为k和1−k,更符合刺槐生长规律。即
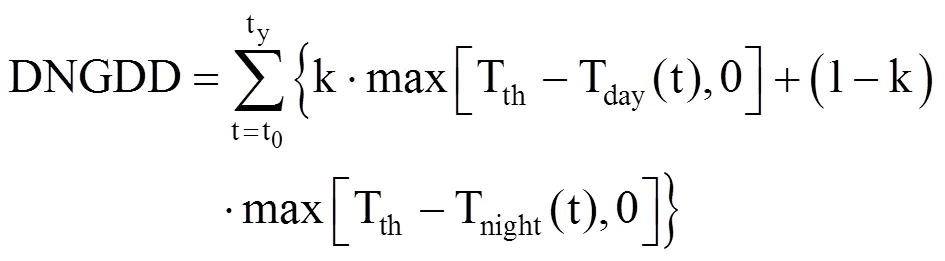
式中,t0为开始累积的日期,春季累积首日为当年1月1日,秋季为7月1日,Tday(t)为白天气温(℃),Tnight(t)为夜晚气温(℃),气象数据为20:00−次日20:00气温,全天测量4次,分别为2:00、8:00、14:00和20:00,出现的最大值为日平均最高气温,即Tday(t),出现的最小值为日平均最低气温,即Tnight(t),y为对应物候期,Tth为界限温度(℃),模拟春季物候期Tth=0℃,叶变色期Tth=20℃,k和1−k为对应权重,考虑了昼夜温差对物候期的影响,该模型较好地模拟了物候对昼夜气温的响应。中国东部暖温带落叶阔叶林区在全球气候区划分中属温带较湿润地区,该气候条件下物候期对昼夜气温的响应相反,因此,取k>1(k=10/9,1−k=−1/9)。DNGDD的阈值需要利用日最低气温、日最高气温和实测物候数据,采用模拟退火法求解得到,利用式(7),当昼夜气温的积温之和达到阈值时,对应的日期为模拟预测的植物物候期日序(ty)。
1.4 物候模型参数估计和模型检验
模型用最小二乘法拟合,使函数f(x)在参数空间x中最小。即
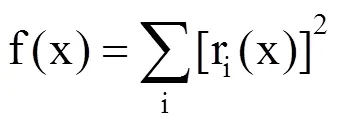

式中,di(x)是第i年的预测日期,di(obs)是第i年的观测日期,最小二乘法无法推导出最优解。因此,选用模拟退火法来求解非线性最小二乘问题,以寻求模型的最优参数[30]。模拟退火是指在金属冷却、退火或液体冻结和结晶过程中与热力学原理的类比,该方法的原理是玻尔兹曼概率分布。模拟退火法从较高气温并设定任意一组参数开始,所有参数的变化也是随机的,每次变化都会产生新的平方和的残差,最终要寻找到目标函数的全局最优解。该方法被广泛应用于模型参数估计中,能更好地找到全局最优解而非局部最优解。
利用Matlab读取10个站点奇数年的气温资料和物候资料,采用模拟退火算法进行参数拟合优化,并进行物候期的模拟,与实测物候数据进行对比,计算出方差解释量R2、均方根误差RMSE、平均绝对误差MAE,此为内部检验[8];将所有站点剩余未用的偶数年的气温数据代入各模型中,模拟预测物候期,与实测物候数据进行对比,计算相应的R2、RMSE和MAE,此为交叉检验。交叉检验能更好地反映模型预测的准确度。
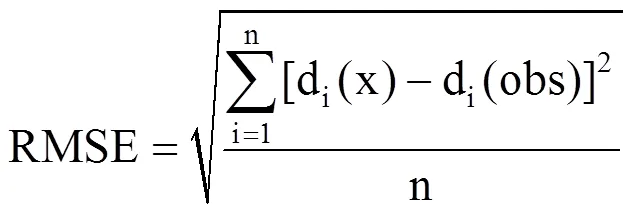
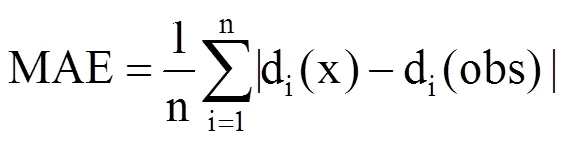
式中,n为某种植物的实际观测数量,di(x)为第i年的预测日期,di(obs)为第i年的观测日期。
2 结果与分析
2.1 物候与气温关系分析
利用10个观测站点的历年物候期观测资料,与同期平均气温、平均日最低气温、平均日最高气温进行相关分析。由表2可以看出,实测刺槐4个主要物候期与同期的各项气温平均值均呈极显著负相关关系(P<0.01),说明平均气温越高,各物候期的日序值越小、物候越提前。但从相关系数的具体数值看,春季物候和秋季物候与不同生育期气温平均值的相关程度略有区别。与展叶始期相比,春季叶芽开放期、开花始期与平均气温、日平均最低气温和日平均最高气温的相关性更高。秋季叶变色期与平均气温、日平均最低气温和日平均最高气温的相关系数均相对较低,且差异不大。因此可见,模拟刺槐物候期时,不但要考虑平均气温的变化,还需要考虑日最低气温和最高气温。

表2 10个站点历年记录刺槐各物候期日序(1月1日起)与同期气温的相关系数
注:*、**分别表示相关系数通过0.05、0.01水平的显著性检验。下同。
Note:*is P<0.05,**is P<0.01. DOY is the ordinal day from Jan.1.The same as below.
2.2 物候模型参数优化
利用所有站点奇数年的气温资料和物候资料,采用模拟退火法进行参数拟合。由表3可以看出,刺槐的生长发育基础气温为0℃(Tb=0℃),符合大多数植物生长发育的基础气温范围(0~35℃),说明估计值Tb=0℃符合刺槐生长的生物学规律。刺槐积温阈值的估计值在各物候期均不相同(表3−表5),其中SW模型和Unichill模型的F*值随物候期的推进先后逐渐增大。表明植物生长发育需逐步累积热量,达到生长发育的热量阈值才会进入下一发育阶段。DNGDD模型中春季叶芽开放期、展叶始期、开花始期的积温阈值也符合上述规律;在叶变色期,由于参数t0与界限温度的改变,积温累积开始日期不同,积温阈值在叶变色期与春季的三个物候期相比变化较大。由此可见,利用模拟退火法对模型进行参数估计得到的数值符合刺槐的生长发育规律。
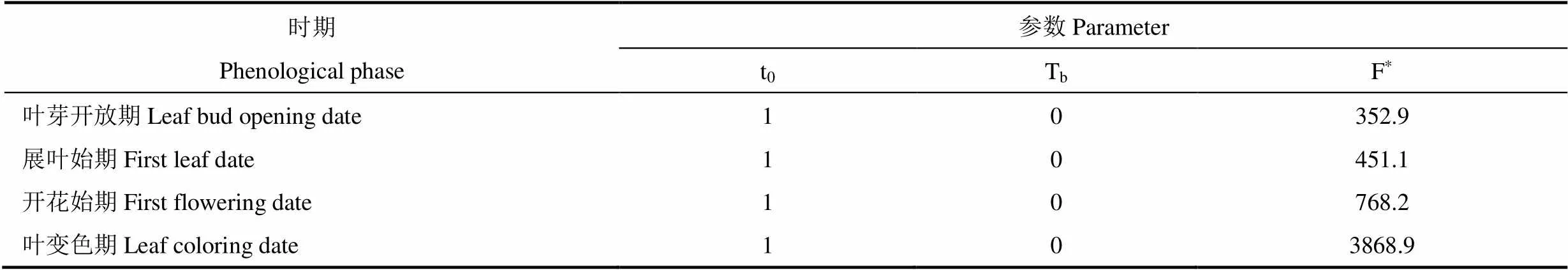
表3 SW模型的参数估计

表4 Unichill模型的参数估计

表5 DNGDD模型的参数估计
2.3 物候模型模拟春季物候期比较
采用三种物候模型结合模拟退火法,对刺槐各物候期进行模拟,并对模拟值与实测值进行对比分析。由图1−图3可见,SW模型得到的物候期模拟值与实测值拟合线最接近1:1线,说明该模型能较准确地模拟刺槐叶芽开放期、展叶始期和开花始期;Unichill模型对春季物候期模拟效果好于DNGDD模型。从方差解释量和均方根误差上看(表6),SW模型交叉检验的均方根误差值小于DNGDD模型,与Unichill模型的值十分接近。对比三种模型交叉检验的方差解释量发现,SW模型的值均大于DNGDD和Unichill模型,在模拟春季物候期中SW模型的方差解释量最优。综合方差解释量与均方根误差,在模拟刺槐春季物候期上,以SW模型为最优,而使用昼夜气温的DNGDD模型效果远不如使用平均气温的SW和Unichill物候模型。
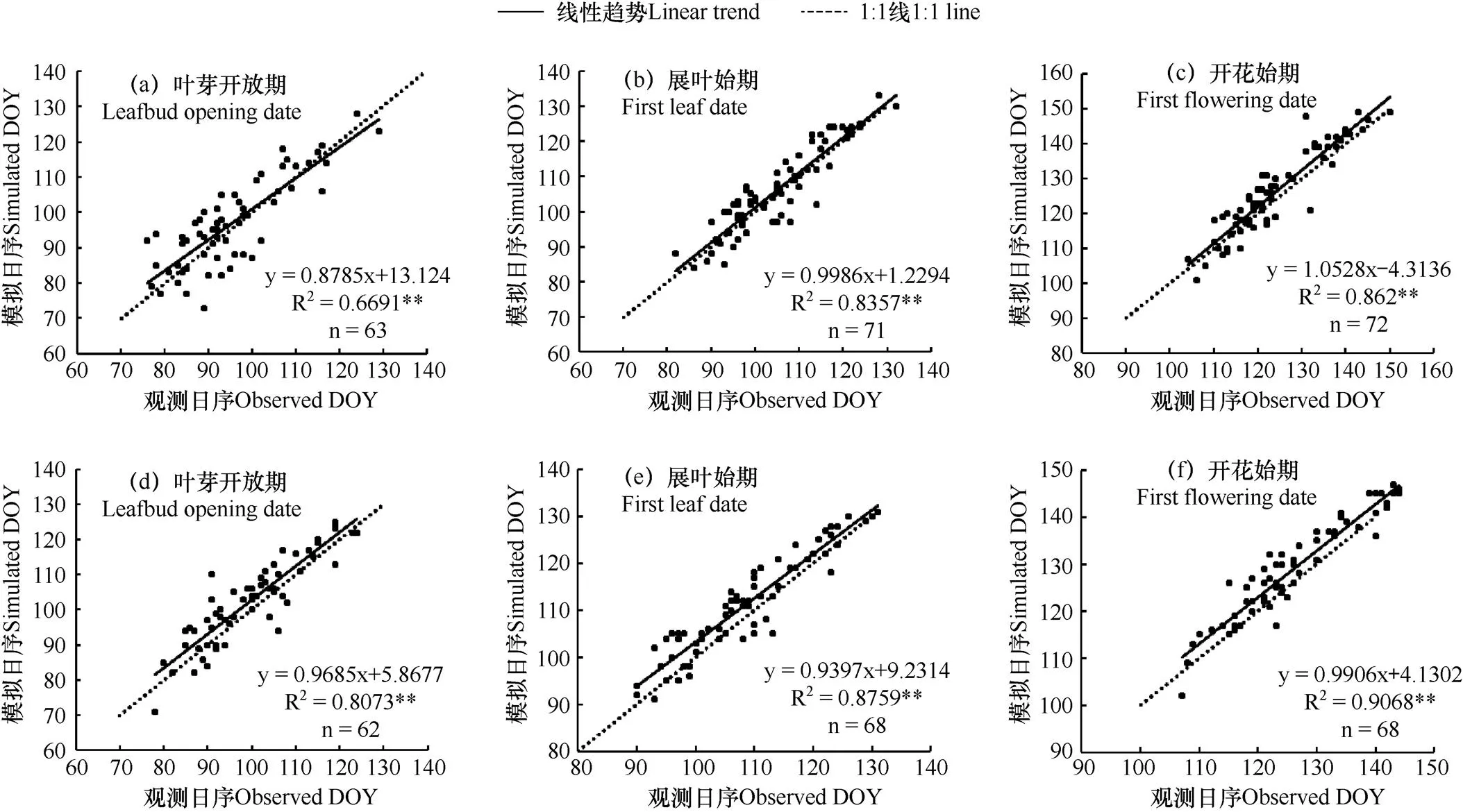
图1 SW模型模拟的春季物候期与实测值的对比
注:图a、b、c为内部检验,使用站点奇数年的数据,图d、e、f为交叉检验,使用站点偶数年的数据。图2、图3同。
Note: DOY is the ordinal day from Jan. l. Fig.a, b and c are internal checks, which the odd-year data from sites was used in, Fig.d, e and f are cross checks, which the even-year data from sites was used in. The same as Fig.2 and Fig.3.
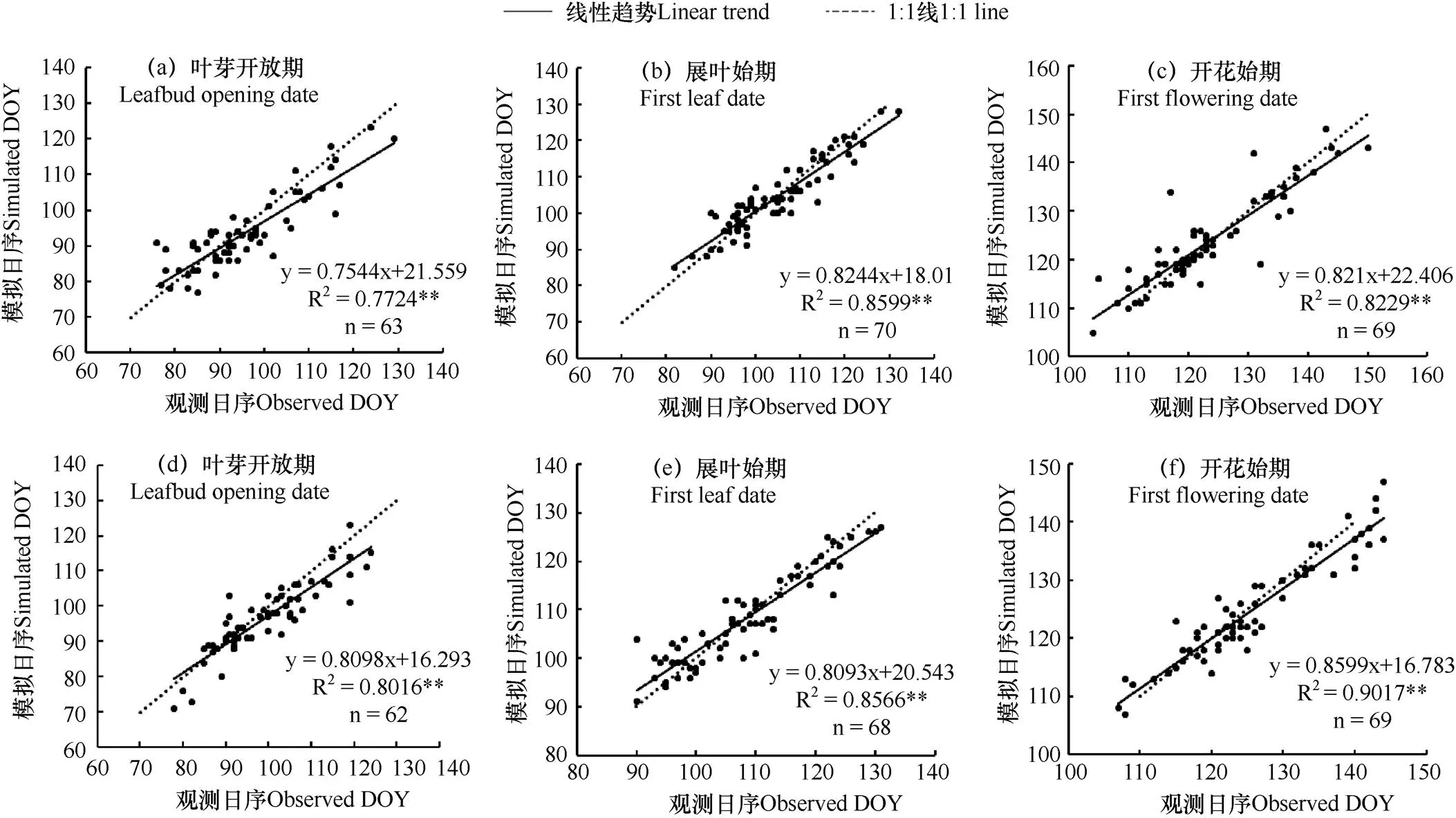
图2 Unichill模型模拟的春季物候期与实测值的对比

图3 DNGDD模型模拟的春季物候期与实测值的对比
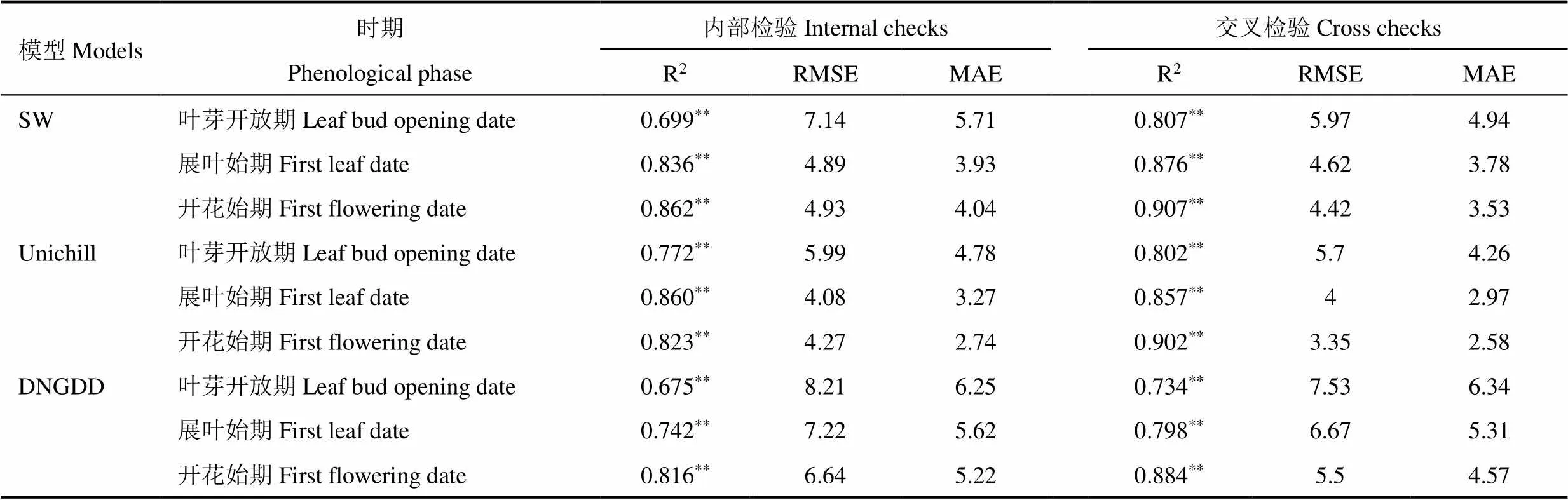
表6 三种模型的检验结果
2.4 物候模型模拟秋季叶变色期比较
使用SW模型和DNGDD模型对刺槐秋季叶变色期进行模拟,实测值与模拟值结果如图4。由图可见,基于DNGDD模型得到的模拟值与实测值的拟合线较SW模型的拟合线更接近1:1线,说明DNGDD模型更准确地模拟了秋季叶变色期。从两者交叉检验的数值上看,DNGDD模型的方差解释量远大于SW模型,均方根误差也小于SW模型 (表7)。因此,在模拟秋季叶变色期时,DNGDD模型表现更佳。表明使用改进后的DNGDD模型对叶变色期模拟的准确度有了极大提升,也是所选模型中最优的。

图4 SW模型和DNGDD模型对秋季叶变色期的模拟结果
注:图a、b为内部检验,使用站点奇数年的数据,图c、d为交叉检验,使用站点偶数年的数据。
Note: Fig.a and Fig.b are internal checks, which the odd-year data from sites was used in, Fig.c and Fig.d are cross checks, which the even-year data from sites was used in.
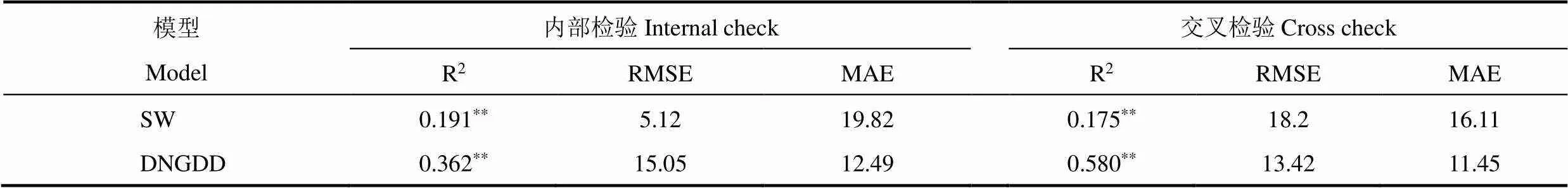
表7 SW和DNGDD模型对叶变色期模拟结果的检验
3 讨论与结论
3.1 讨论
本研究以中国东部暖温带刺槐为研究对象,结合气象数据与物候资料,对比分析SW模型、Unichill模型和DNGDD模型对春秋季物候期的模拟效果。在模拟春季物候期上,SW模型模拟效果最优,这是由于本研究区在暖温带地区,冷激需求的限制并不起决定作用,限制物候期的主要因素为春季积温,而SW模型考虑了春季积温,并未考虑冷激因素。王炳赟等[31]利用热时模型模拟了川渝地区两种刺槐和一种紫荆的展叶始期和开花始期,发现仁寿和北碚刺槐展叶期的观测值与模拟值的拟合度可分别达到87.5%和100%,在模拟春季物候时SW模型模拟效果最好。张爱英[32]在对北京地区观赏植物开花预测中发现,SW模型的交叉检验的方差解释量R2最高,RMSE最小,明显优于统计模型的结果,最为准确地模拟了北京地区的植物花期。对牡丹江地区的植物花期进行模拟发现,使用SW模型数据的方差解释量R2达到了0.71,明显高于回归模型的方差解释量[33]。本研究对SW模型模拟刺槐的春季物候期作出了补充,对其适用范围进行推广。但仅仅对刺槐一种典型树种进行研究不足以说明SW模型在春季物候模拟上的普适性,今后应扩大研究对象,选取多种典型乔木与灌木,对SW模型的模拟效果作进一步研究。SW、Unichill和DNGDD均基于积温理论,要求研究对象为同一种类型的植物或是已知该地区的经纬度依赖于当前的气候条件[34],若扩大研究范围,则应结合当地的经纬度与气候条件对模型进行改进。仲舒颖等[35]对全国范围内观赏植物的花期进行了模拟,将传统春季积温模型进行改进,与时空特征相联系得到始花期和末花期的方差解释量均大于0.8,RMSE在4~6d,也为模型的改进提供了研究思路,同时也证明影响植物开花的因素除了气温,当地的水热条件对物候期具有一定的影响[36]。
SW模型较准确地模拟了刺槐春季物候期,但其对叶芽开放期、展叶始期、开花始期交叉检验的方差解释量R2分别仅为0.807、0.876、0.907。由此可见,用仅考虑日平均气温的模型模拟植物物候是不够的,除气温外,降水、光周期等也影响植物的生长发育。Chen等[37]在对青藏高原地区的植被进行模拟时,除利用传统SW模型等外,还采用了TPP模型,即考虑降水和气温双重影响的模型,季前降水量与气温类似必须达到降水的临界值,结合降水等气象因素对传统SW模型进行了改进,也进一步证明了仅用日平均气温的物候模型所模拟的物候期偏差较大。本研究中,SW、Unichill和DNGDD模型仅考虑了气温的影响,未考虑到其它气象因素,于是造成模拟准确度不高,今后需考虑降水和光周期的影响,以进一步提高模型模拟的准确性。
目前,对于温带季风区刺槐叶变色期模拟的研究相对较少。本研究选用了SW模型和DNGDD模型对刺槐叶变色期模拟发现,DNGDD模型效果较优但其方差解释量也仅0.58,说明基于积温理论的物候模型对秋季物候期的模拟效果并不理想。因此,除传统物候模型外,目前已有许多学者使用遥感对物候期进行模拟和验证。曹沛雨等[38]基于已有研究分类评述了当前常用的植被物候观测技术,得出应合理评估多源数据和多种方法间植被物候的差异。Luo等[39]利用NDVI数据结合模型对生长季开始期(SOS)进行分析与预测,表明在整个区域内SOS每10a将提前0.65~1.79d。基于遥感的NDVI数据和区域模型模拟的结果相结合是目前研究物候的主要趋势,地面观测数据的缺失,使得数据不连续,遥感方法则从大尺度范围对物候进行观测,利用NDVI数据模拟物候期弥补了物候观测单站点的不足。
3.2 结论
SW模型对刺槐叶芽开放期、展叶始期和开花始期模拟,其交叉检验方差解释量R2最大(0.807~0.907),模拟效果最佳且误差最小。SW模型模拟春季物候期的效果优于Unichill模型,春季物候期与平均气温的相关性最强,而与平均日最低气温和平均日最高气温的相关性不显著,DNGDD模型采用日最低气温和日最高气温代替日平均气温模拟物候期,造成其模拟效果低于基于日平均气温的SW模型。
对于叶变色期的模拟,SW模型模拟值与观测值的误差较大,DNGDD模型交叉检验的方差解释量R2最高,其值为0.580。秋季昼夜温差较大,白天和夜间气温对物候的影响会有所不同。因此,仅用平均气温模拟叶变色期会带来很大偏差。SW模型用日平均气温作为驱动变量模拟春季物候,DNGDD模型则采用日最低气温和日最高气温代替平均气温,并根据植物的特征和观测点的特点选择最优的昼夜气温的比重。因此,DNGDD模型能较准确地模拟叶变色期,而SW、Unichill模型更适用于对春季物候的模拟。
[1]Parmesan C,Yohe G.A globally coherent fingerprint of climate change impacts across natural systems[J].Nature, 2003,421:37-42.
[2]Chuine I.Why does phenology drive species distribution[J]. Philosophical Transactions(Biological Sciences),2010,365 (1555):3149-3160.
[3]Walther G R,Post E,Convey P,et al.Ecological responses to recent climate change[J].Nature,2002,416:389-395.
[4]Forrest J,Miller-Rushing A J.Toward a synthetic understanding of the role of phenology in ecology and evolution[J].Philosophical Transactions-Royal Society (Biological Sciences),2010,365:3101-3112.
[5]Qin D,Manning M,Chen Z,et al.IPCC,2007:Summary for policymakers[R]//Climate change 2007:the physical science basis.Computational Geometry,2013,18(2):95-123.
[6]Menzel A,Estrella N,Fabian P.Spatial and temporal variability of the phenological seasons in Germany from 1951 to 1996[J].Global Change Biology,2001,7:657-666.
[7]Ahas R,Aasa A,Menzel A,et al.Changes in European spring phenology[J].International Journal of Climatology, 2002,22,1727-1738.
[8]Badeck F W,Bondeau A,Böttcher K,et al.Responses of spring phenology to climate change[J].New Phytologist, 2004,162:295-309.
[9]张福春.气候变化对中国木本植物物候的可能影响[J].地理学报,1995,50(5):402-410.
Zhang F C.Possible effects of climate change on the phenology of woody plants in China[J].Acta Geographica Sinica,1995,50(5):402-410.(in Chinese)
[10]朱媛君,郝琦,清华,等.呼和浩特自然历主要物候期和物候季节变化特征及归因[J].中国农业气象,2017,38(1): 9-20.
Zhu Y J,Hao Q,Qing H,et al.Changes of main phenophases of natural calendar and phenological seasons in Hohhot[J].Chinese Journal of Agrometeorology,2017,38(1): 9-20.(in Chinese)
[11]王明,桑卫国.暖温带乔木和灌木物候变化及对气候变化的响应[J].生态科学,2020,39(1):164-175.
Wang M,Sang W G.The change of phenology of tree and shrub in warm temperate zone and their relationships with climate change[J].Ecological Science,2020,39(1):164-175. (in Chinese)
[12]刘璐,王景红,傅玮东,等.中国北方主产地苹果始花期与气候要素的关系[J].中国农业气象,2020,41(1):51-60.
Liu L,Wang J H,Fu W D,et al.Relationship between apple’s first flower and climate factors in the main producing areas of the Northern China[J].Chinese Journal of Agrometerology, 2020,41(1):51-60.(in Chinese)
[13]Cleland E E,Chuine I,Menzel A,et al.Shifting plant phenology in response to global change[J].Trends in Ecology & Evolution,2007,22:357-365
[14]Hunter A F,Lechowicz M J.Predicting the timing of budburst in temperate trees[J].Journal of Applied Ecology, 1992,2:597-604.
[15]王焕炯,戴君虎,葛全胜.1952-2007年中国白蜡树春季物候时空变化分析[J].中国科学:地球科学,2012,4(25): 701-710.
Wang H J,Dai J H,Ge Q S.The spatiotemporal characteristics of spring phenophase changes ofin China from 1952 to 2007[J].Science China: Earth Science,2012,4(25):701-710.(in Chinese)
[16]秦柏凤,霍治国,王景红,等.中国富士系苹果主产区花期模拟与分布[J].中国农业气象,2020,41(7):423-435.
Qin B F,Huo Z G,Wang J H,et al.Simulation and distribution of flower stage in main production areas of Fuji apple in China[J].Chinese Journal of Agrometeorology, 2020,41(7):423-435.(in Chinese)
[17]李荣平,张晓月,李晶.气候变化背景下紫丁香花期物候特征及其模拟研究[J].北方园艺,2018(1):109-113.
Li R P,Zhang X Y,Li J.Flowering phenology characteristics and simulation of lilacs under climate change[J].Northern Horticulture,2018(1):109-113.(in Chinese)
[18]Delpierre N,Dufrêne E,Soudani K,et al.Modelling interannual and spatial variability of leaf senescence for three deciduous tree species in France[J].Agricultural and Forest Meteorology,2009,149:938-948.
[19]Park C K,Ho C H,Jeong S J,et al.Spatial and temporal changes in leaf coloring date of Acer palmatum and Ginkgo biloba in response to temperature increases in South Korea[J].Public Library of Science,2017,12(3): e0174390.
[20]汪箫悦.基于多源数据的区域植被物候变化模拟与影响机制研究[D].北京:中国科学院大学,2018.
Wang X Y.Study on the simulation and influence mechanism of regional vegetation phenology based on multi-source data[D].Beijing:University of Chinese Academy of Sciences,2018.(in Chinese)
[21]Piao S L,Liu Q,Chen A P,et al.Plant phenology and global climate change:current progresses and challenges[J].Global Change Biology,2019,25:1922-1940.
[22]赵蓬晖,张江涛,王念.刺槐原产地分布及世界各国引种与研究概况[J].河南林业科技,2017,37(1):30-32. Zhao P H,Zhang J T,Wang N.The original distribution introduction and development of[J]. Journal of Henan Forestry Science and Technology,2017, 37(1):30-32. (in Chinese)
[23]宛敏渭,刘秀珍.中国物候观测方法[M].北京:科学出版社,1979:1-136.
Wan M W,Liu X Z.Chinese phenological observation method[M].Beijing:Science Press,1979:1-136.(in Chinese)
[24]Botta A,Viovy N,Ciais P,et al.A global prognostic scheme of leaf onset using satellite data[J].Global Change Biology, 2000,6(7):709-725.
[25]Cannell M G R,Smith R I S.Thermal time,chill days and prediction of budburst in[J].Journal of Applied Ecology,1983,20:951-963.
[26]Hunter A F,Lechowicz M J.Predicting the timing of budburst in temperate trees[J].Journal of Applied Ecology, 1992,29:597-604.
[27]Murray M B,Cannell M G R,Smith R I.Date of budburst of fifteen tree species in Britain following climatic warming[J].Journal of Applied Ecology,1989,26:693-700.
[28]Cleland E E,Chuine I,Menzel A,et al.Shifting plant phenology in response to global change[J].Trends in Ecology & Evolution,2007,22:357-365.
[29]Fu Y H,Liu Y J,De Boeck,et al.Three times greater weight of daytime than of night-time temperature on leaf unfolding phenology in temperate trees[J].New Phytologist, 2016,212:590-597.
[30]Chuine I,Cour P,Rousseau D D.Fitting models predicting dates of flowering of temperate-zone trees using simulated annealing[J].Plant Cell and Environment,1998,21:455-466
[31]王炳赟,范广洲,魏鸣,等.基于温度影响因子的植物物候模型的应用研究[J].生物数学学报,2015,30(2):253-262.
Wang B Y,Fan G Z,Wei M,et al.Application research of plant phenology model based on temperature[J].Journal of Biomathematics,2015,30(2):253-262.(in Chinese)
[32]张爱英,王焕炯,戴君虎,等.物候模型在北京旅游观赏植物开花期预测中的适用性[J].应用气象学报,2014,25(4): 483-492.
Zhang A Y,Wang H J,Dai J H,et al.Applicability analysis of phenological models in the flowering time prediction of ornamental plants in Beijing area[J].Journal of Applied Meteorological Science,2014,25(4):483-492.(in Chinese)
[33]徐韵佳,仲舒颖,戴君虎,等.1978-2014年牡丹江地区植物花期变化及模型模拟[J].地理研究,2017,36(4):779-789.
Xu Y J,Zhong S Y,Dai J H,et al.Changes in flowering phenology of plants and their model simulation in Mudanjiang,China[J].Geographical Research,2017,36(4):779-789.(in Chinese)
[34]徐琳,陈效逑,杜星.中国东部暖温带刺槐花期空间格局的模拟与预测[J].生态学报,2013,33(12):3584-3593.
Xu L,Chen X Q,Du X.Simulation and prediction of spatial patterns offlowering dates in eastern China's warm temperate zone[J].Acta Ecologica Sinica,2013,33(12):3584-3593.(in Chinese)
[35]仲舒颖,葛全胜,戴君虎,等.中国典型观赏植物花期模型建立及过去花期变化模拟[J].资源科学,2017,39(11): 2116-2129.
Zhong S Y,Ge Q S,Dai J H,et al.Development of phenological models for simulating past flowering phenology of typical ornamental plants in China[J]. Resources Science,2017, 39(11):2116-2129.(in Chinese)
[36]Berninger F.Effects of drought and phenology on GPP in:a simulation study along a geographical gradient[J].Functional Ecology,1997,11:33-42.
[37]Chen X Q,Li J,Xu L,et al.Modeling green-up date of dominant grass species in the Inner Mongolian Grassland using air temperature and precipitation data[J].International Journal of Biometeorology,2014,58:463-471.
[38]曹沛雨,张雷明,李胜功,等.植被物候观测与指标提取方法研究进展[J].地球科学进展,2016,31(4):365-376.
Cao P Y,Zhang L M,Li S G,et al.Review on vegetation phenology observation and phenological index extraction[J].Advances in Earth Science,2016,31(4):365- 376.(in Chinese)
[39]Luo X Z,Chen X Q,Wang L X,et al.Modeling and predicting spring land surface phenology of the deciduous broadleaf forest in northern China[J].Agricultural and Forest Meteorology,2014,198/199:33-41.
Comparison of Phenological Models of(L.) in the Warm- temperate Region of Eastern China
YU Pei-yang1, TONG Xiao-juan1, LI Jun2, ZHANG Jing-ru1, LIU Pei-rong1
(1. School of Ecology and Nature Conservation, Beijing Forestry University, Beijing 100083, China;2. Key Laboratory of Water Cycle and Related Land Surface Processes, Institute of Geographic Sciences and Nature Resources Research, CAS, Beijing 100101)
Vegetation phenology is a powerful indicator of the response of terrestrial ecosystems to climate change and plays an important role in water, carbon exchange and energy balance. Under the background of global warming, the changes of vegetation phenology and simulating the phenological phase of plants have been paid much attention. Up to date, the performance of most phenolgy models on spring phenolgy is better, but the simulated autumn phenology has been less accurate. In this study, meteorological data and surface phenological data (leaf bud opening date, first leaf date, first flowering date and leaf coloring date) at 10 sites were selected as the input of the SW, Unichill and DNGDD models, which were used to simulate the phenological phase of(L.). The objectives were to examine the performace of SW, Unichill and DNGDD models simulating spring and autumn phenology, and give a guide on optimum the parameters of phenology models. The odd-year data of the sites were used as the internal test, and the even-year data were used as the cross test. The simulation values of three models were compared with the observed ones for analysis to find the best one to predict vegetation phenology. The simulated annealing algorithm was applied to optimum the parameters of SW, Unichill and DNGDD models. The simulated spring and autumn phenology was compared with the measured one, and evaluate the modelling to select the best one to predict vegetation phenology.The results showed that the leaf bud opening, the first leaf, the first flowering dates of(L.) were significantly negatively correlated with average temperature in the same period. Average temperature was the main factor affecting phenology. The phenological period in spring had the strongest correlation with mean air temperature, but not significant correlation with mean daily minimum temperature and mean daily maximum temperature. Therefore, compared with Unichill and DNGDD models, SW performed better in simulating spring phenology. The variance interpretation (R2) of cross test of SW model simulating the leaf bud opening, the first leaf and the first flowering dates were 0.807, 0.876 and 0.907 separately and the root mean square error (RMSE) 6.0, 4.6 and 4.4 days. Mean temperature in the daytime and night mean temperature in autumn had different influences on phenology. There was a large deviation when leaf coloring date was simulated only using average temperature. Average temperature was replaced by daily minmum and maximum temperature in DNGDD model. DNGDD model performed well in comparison with SW model when the leaf coloring date in autumn was simulated, and R2of cross check was 0.580. Therefore, SW model is the best one to simulat the spring phenological phases, while DNGDD model perforemed well in simulating the autumn phenological phase.
; Phenology model; Spring phenology; Autumn phenology; Integrated temperature; Simulated annealing
10.3969/j.issn.1000-6362.2020.10.001
于裴洋,同小娟,李俊,等.中国东部暖温带刺槐物候模型比较[J].中国农业气象,2020,41(10):609-621
2020-06-23
同小娟,E-mail:tongxj@bjfu.edu.cn
国家自然科学基金项目(31872703;31570617)
于裴洋,E-mail:ypy0913@163.com
