洞庭湖湿地植物群落多样性及土壤有机碳储量研究
2020-10-21梁春玲
梁春玲
(商丘师范学院 测绘与规划学院, 河南 商丘 476000)
土壤有机碳(soil organic carbon,SOC)是土壤—植物生态系统的重要组成部分,对土壤物理、化学和生物特征的变化起着关键作用[1-3]。作为陆地生态的关键构成部分,土壤发挥着无可替代的作用[4],能够为植被生长发育提供必需的养分及水分,形成了土壤—植被生态子系统[5-6],同时有机碳SOC不仅影响着土壤碳库,同时制约着其理化特性,参与能量交换,成为土壤生态中不可或缺的部分[7-8]。有机碳对于促进陆地生态平衡发挥着巨大作用,作为重要的养分库,其含量也处于不断变化之中,制约土壤质量及植被生长。在土壤理化特性中,土壤养分与碳库调节效应密切相关,从而促进土壤质量的稳定。对于土壤碳库而言,其不仅拥有巨大的储量,同时具有巨大的外在影响性,其作为陆地最大的碳库,储量远超大气碳库,即使是微小的变化,都将对整个碳库施加重要影响。对于土壤有机碳而言,其在反映土壤肥力方面具有良好的效果,且其之间具有较为紧密的关系,同时能够参与到土壤的理化反应之中,作为一种动力促进着土壤碳代谢,关于土壤养分状况,制约土壤质量,同时对于碳库平衡施加着无可替代的影响,此外,其在陆地养分循环及能量交换过程中发挥着重要角色,直接制约着养分循环,成为影响土壤质量的关键因素之一,同时直接制约有机质的转化效果[9],从而在陆地碳循环过程中起着无可替代的作用。
作为国内的第二大淡水湖,洞庭湖经常出现明显的水位变化,这种变化具有明显的周期性,夏冬季节的水位变化差异较大,素有“夏季湖相,冬季河相”[10],主要原因在于水期的差异影响所致。对于雨水充足的季节,由于其水位高而呈现较为突出的湖相特点,在水位不断下降的情况下,出现不同程度的洲滩显露现象,滩地及沼泽随之增多,加之湿地植被的不断生长,从而出现了河湖交错的多种水域形态特点[11-12]。对于该区域而言,其吞吐性湖泊特点明显,水位的梯度变化尤为突出,此外,容易出现明显的洲滩淤积抬升现象,这也是该区域较为突出的地方。由于洞庭湖湿地区域广布,生物群落分布多样性明显,加之其水文条件的复杂性,环境因子的影响效应突出,因此探究该区域湿地研究具有重要的现实意义,大量学者围绕着湿地植被、水文特点等方面开展了相关研究,并取得了丰硕的研究成果,此外,不少学者围绕着湿地土壤及水质等方面开展了长期的观测研究,尤其是湿地土壤肥力、活性、微生物、酶等方面,但是对于该区域的有机碳研究相对较少[13-14]。由于有机碳关乎局地生态碳平衡,其变化将会产生巨大的关联影响,同时其关乎物质和能量循环,因此开展这方面的研究具有重要的现实意义。受地理位置的制约效果明显,对于洞庭湖湿地区域而言,其温湿气候特点较为突出,适宜的气温对于植被生长尤为有利,促进了生物的群落分布,同时也增强了其生物分布的复杂性;由于其水位呈现明显的梯度变化特点,洲滩分布较多,加之冲刷导致的淤积抬升,其呈现了较为明显的洲滩分布,随着距离的增加,植被群落分布从荫草(Sciaphilatenella)、水蓼(Polygonumhydropiper)逐渐向苔草(Carexparva)、芦苇(Phragmitesaustralis)过渡,形成了层次较为分明的植被群落。此外,微生物活动在湿地土壤质量保持过程中效果尤为显著,探究有机碳、微生物活动对于探讨该区域土壤理化特性具有重要作用。基于此,本研究选取荫草群落、水蓼群落、苔草群落、芦苇群落,进行连续3 a的实地观测分析,在对生物分布多样性加以探究的同时,注重探究有机碳分布特性,探寻其交互作用机理,从而为湿地保护提供有益参考和借鉴。
1 材料与方法
1.1 采样区域
本研究选取洞庭湖湿地,该区域面积广阔,不仅在湖南湘水等“四水”的调节方面作用突出,而且关乎长江水域的调蓄,发挥着“长江之肾”的作用,该区域主要分布大量的冲击平原,根据近些年来该地区的气象资料分析得知,其平均海拔在40 m以下,拥有多达2 625 km2的天然湖泊分布。受地理位置制约而具有较为突出的亚热带季风气候特点,四季分明,年均气温约17℃,年均降雨量超过1 400 mm;其低温、高温天气分别出现在1月、7月,月均温度分别为4.7,29℃,无霜期接近280 d。
1.2 植被调查
本试验开始于2016年8月,在洞庭湖湿地按照高程的上升,分别选取荫草群落、水蓼群落、苔草群落、芦苇群落,连续3 a的实地监测,每种植物群落置5个重复样地,每个样地面积约为50 m×50 m。按照对角线布设2条样带15个1 m×1 m的样方,共计300个样方,每个样方按照“S”型五点采样混合后收集土壤样品,然后将其杂质去除,待其自然风干,捣碎后过20目筛,选取样品50 g进行磨细处理,过10,60目筛后置于聚乙烯袋测定土壤有机碳,每个样品重复测量3次取平均值。
每个样方调查植被多样性指数[15]:
H=-∑(PilnPi)
(1)
S=(N-1)/lnN
(2)
D=1-∑(Pi)2
(3)
式中:H为Shannon-Wiener指数;S为Margalef指数;D为Simpson指数;Pi为重要值;N为物种数目。
对植被β多样性的研究过程中将从以下几个方面进行分析[15],具体如下:
βc=(G+L)/2
(4)
Cs=2j/(a+b)
(5)
式中:βc为Cody指数;Cs为Sorenson指数;G为沿生境增加的物种数;L为沿生境减少的物种数;j为共有物种数目;a为A群落物种数;b为B群落物种数。
1.3 土样采集和土壤碳储量计算
土壤有机碳含量采用外加热法[16]:
(6)
式中:SOCi为有机碳储量;Hi,BDi分别为第i层土壤的厚度、容重;Ri为砾石百分比。
2 结果与分析
2.1 不同湿地植被群落α多样性
由图1可知,荫草群落、水蓼群落、苔草群落、芦苇群落多样性各指标表现出较一致的变化规律。Margalef丰富度指数、Shannon-Wiener多样性指数和Mclntosh均匀度指数表现为:芦苇群落>苔草群落>水蓼群落>荫草群落,随着年份的增加呈逐渐增加趋势;芦苇群落和苔草群落的这3个指数显著高于水蓼群落和荫草群落(p<0.05)。对于Simpson优势度指数,则表现为相反的变化趋势,具体表现为芦苇群落<苔草群落<水蓼群落<荫草群落,随着年份的增加呈逐渐减小趋势;芦苇群落和苔草群落Simpson优势度指数显著低于水蓼群落和荫草群落。综合来看,芦苇群落环境较为复杂,其植物α多样性指标值高于其他生境群落的生物多样性指标值。
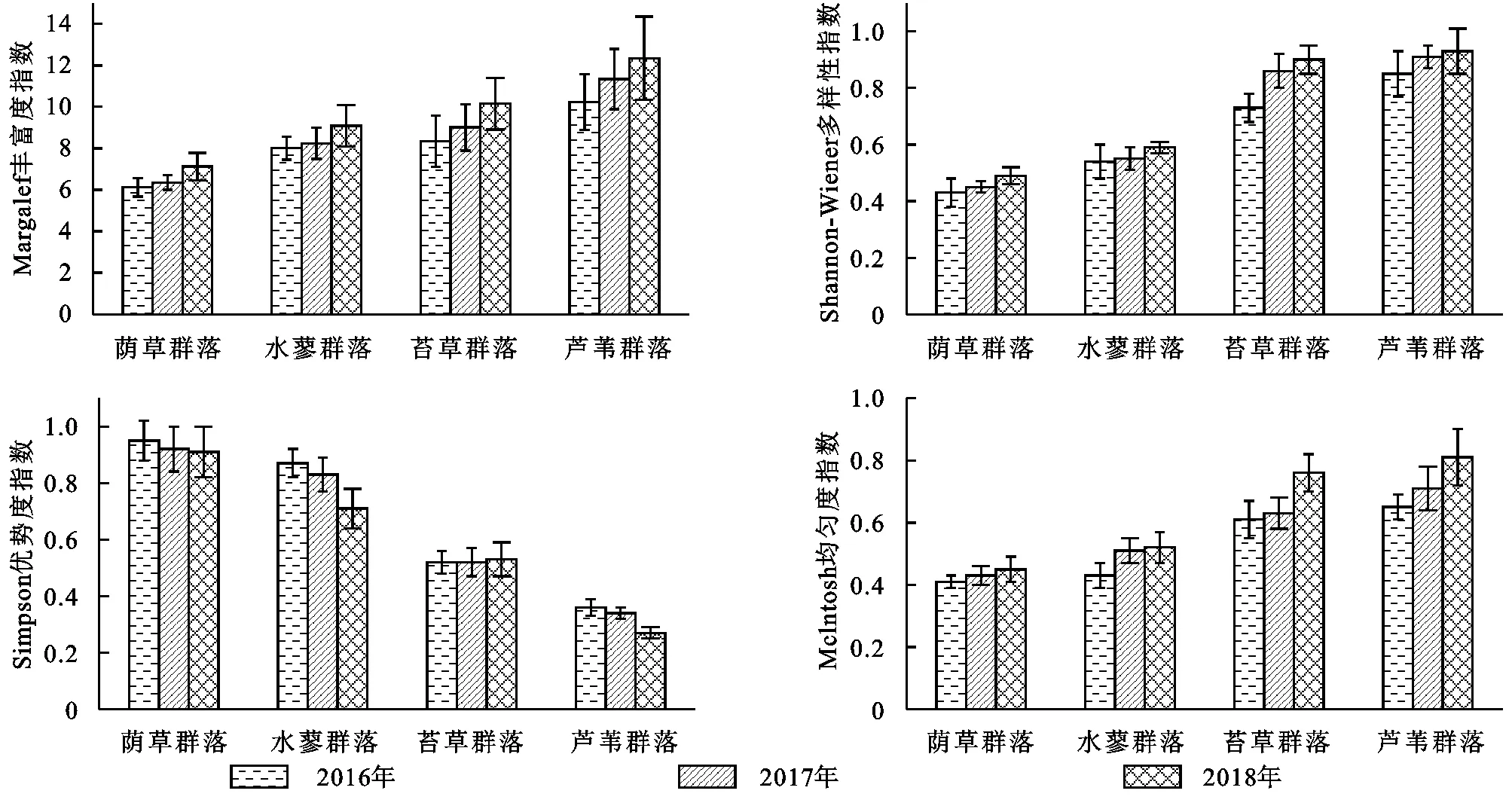
图1 不同湿地植被群落α多样性
2.2 不同湿地植被群落β多样性
Cody指数主要是通过对新增加和失去的物种数目进行比较,从而获得有关物种替代的信息。不同群落或环境梯度上不同点之间共有种越少,β多样性就越高。β多样性越大,物种替代速率越大。相反,β多样性减小,物种替代速率也减小。β多样性反映了群落结构和功能的真实信息。由图2可知,生境间Cody指数依次表现为:芦苇群落>苔草群落>水蓼群落>荫草群落,随着年份的增加呈逐渐增加趋势;Sorenson指数表现为相反的变化趋势,具体表现为芦苇群落<苔草群落<水蓼群落<荫草群落,随着年份的增加呈逐渐减小趋势。综合表明,芦苇群落相似性较大,说明生境间共有物种数多;荫草群落资源异质性明显,说明在此环境梯度上不同点之间的共有种数少,β多样性越大,物种替代速率就越大。
2.3 不同湿地植被群落土壤有机碳含量和有机碳储量
由表1可知,土壤有机碳含量和有机碳储量均呈一致的变化规律,其中以表层土壤最高,随土层深度的增加逐渐降低,其中20—40 cm以下土壤有机碳含量变化范围相对较小;60—80 cm土壤有机碳含量最低。
随剖面深度的增加,土壤有机碳储量(每20 cm)逐渐降低,以表层土壤(0—20 cm)有机碳储量最高,芦苇群落、苔草群落、水蓼群落、荫草群落土壤有机碳储量分别占土壤剖面总有机碳储量的比例为31.23%,33.45%,31.25%,32.78%;60—80 cm土壤有机碳储量最低。由图3可知,0—20 cm土壤有机碳占总有机碳储量百分比表现为芦苇群落、苔草群落较高,60—80 cm表现为水蓼群落、荫草群落较高。

图2 不同湿地植被群落β多样性
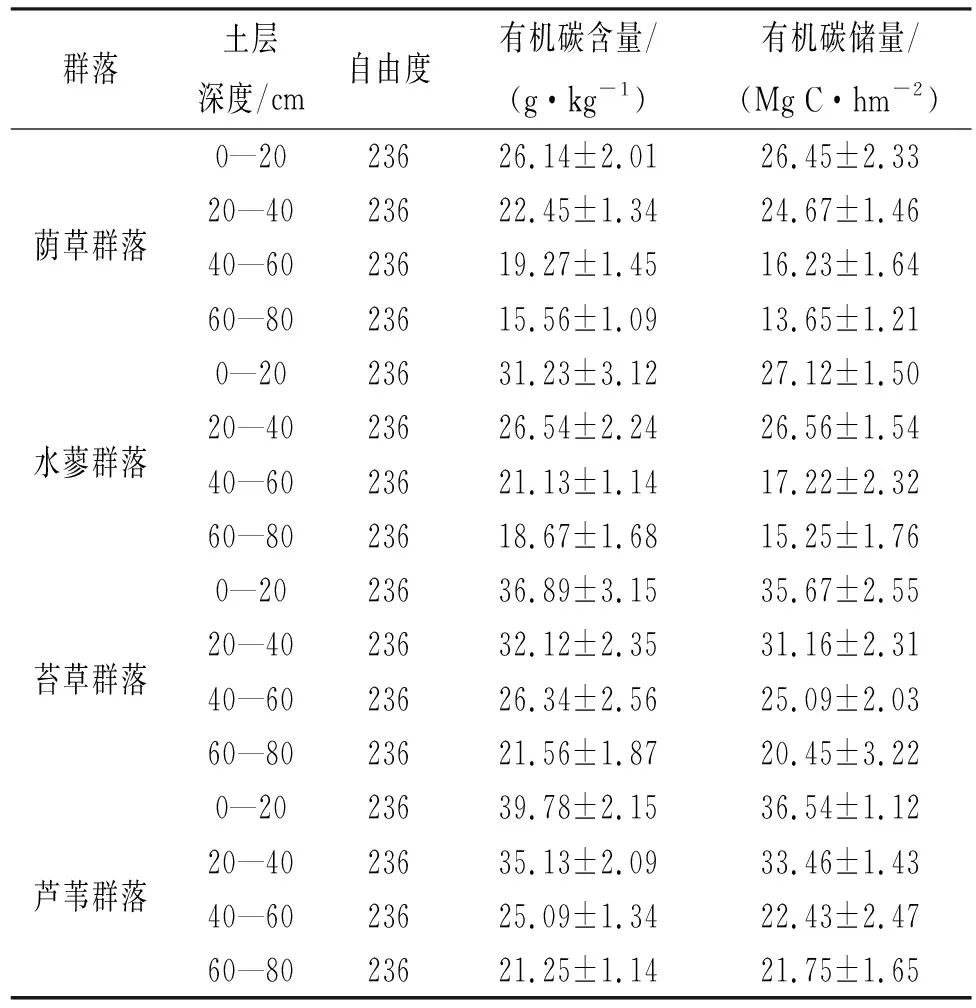
表1 不同湿地植被群落土壤有机碳含量和有机碳储量垂直分布
2.4 植被多样性与有机碳含量和有机碳储量的关系
通过各指标相关性分析表明(表2):芦苇群落、苔草群落、水蓼群落、荫草群落Margalef丰富度指数和Shannon-Wiener多样性指数与有机碳含量和有机碳储量呈显著或极显著的正相关;Cody指数和Sorenson指数与有机碳含量和有机碳储量呈显著或极显著的正相关;Simposon优势度指数和McIntosh均匀度指数则与其没有显著的相关性,并且Simposon优势度指数与有机碳含量和有机碳储量呈负相关;从相关系数绝对值来看,与有机碳含量的相关系数高于有机碳储量的相关系数,并且芦苇群落、苔草群落的相关系数绝对值高于水蓼群落、荫草群落。

图3 表层土壤有机碳储量占剖面总有机碳储量的百分比
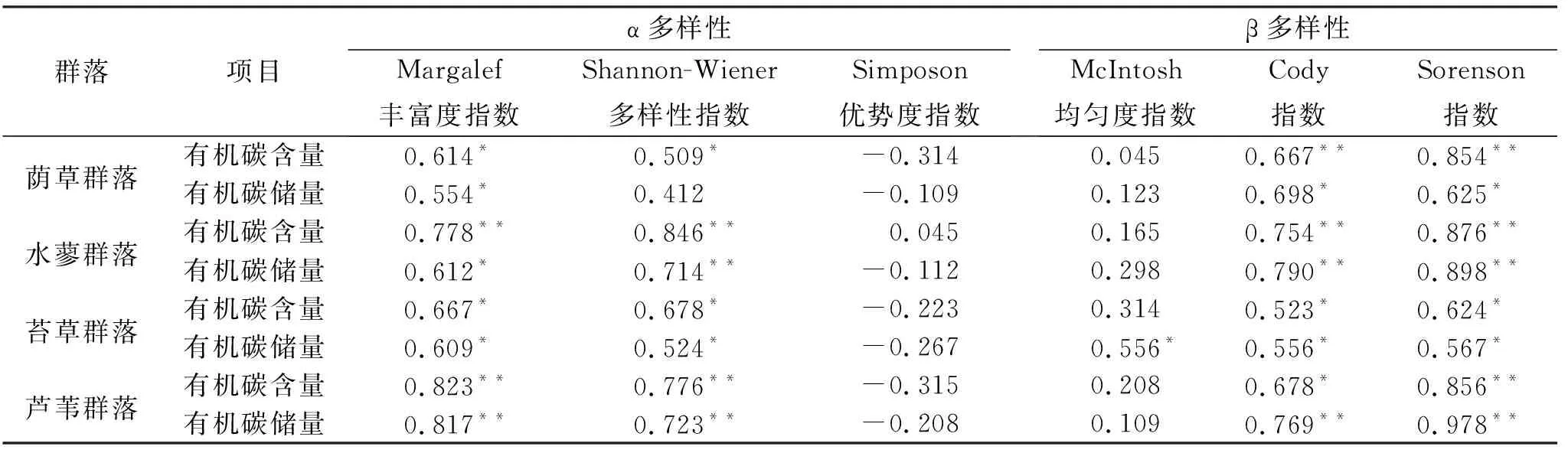
表2 植被多样性与有机碳含量和有机碳储量的关系
2.5 土层深度和植被类型对多样性和有机碳储量的影响
由表3—4可知,土层深度、植被类型和年份对多样性和有机碳储量具有显著的影响,其中植被类型、年份、植被类型×年份对有机碳含量和有机碳储量具有显著的影响(p<0.05);植被类型×年份对Margalef丰富度指数和Shannon-Wiener多样性指数具有显著的影响(p<0.05);植被类型、年份、植被类型×年份对Cody指数和Sorenson指数具有显著的影响(p<0.05)。
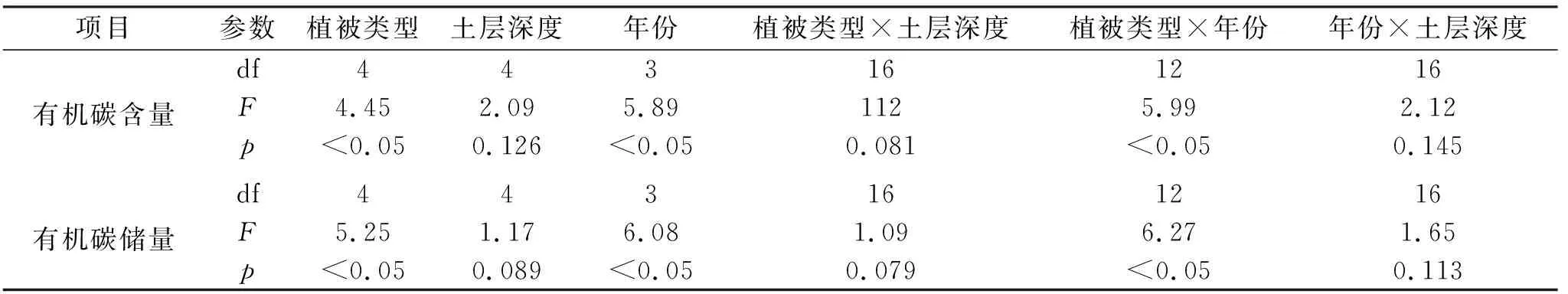
表3 土层深度和植被类型对土壤有机碳储量的影响

表4 土层深度和植被类型对植被多样性的影响
3 讨 论
物种的分布不仅直接取决于所在的生长环境,同时受制于人类的开发利用方式,环境因子及人为因素共同决定了物种群落分布,由于环境因子及人为因素具有较强的动态性,因此其群落分布也处于不断的变化之中。大量学者通过研究发现,丰度及均匀度等能够较好地体现物种分布[17-18],这在群落分布研究过程中常常加以运用。通过本研究得知,在多样性指数方面表现最好的是芦苇湿地,其次是苔草群落,而荫草群落最差,随着时间的推移,其指数也在不断增加;Cody指数亦是如此,指数最高的依然是芦苇群落。而对于Simpson指数来讲,其变化趋势与多样性刚好相反,荫草群落具有更高的优势度指数,而芦苇群落最低,在时间不断增加的情况下该指数出现明显的下降态势;对于Sorenson指数来说亦是如此。综合来看,对于芦苇湿地而言,其群落分布较为类似,这表明其存在较明显的共生物种现象,对于荫草群落而言,存在突出的异质性,物种的梯度变化较为突出。
通过研究对比分析得知,在土层深度变化的情况下,其有机碳含量也呈现较大不同,其原因是多方面的,凋落物的分布及其分解能够显著制约有机碳含量,淋溶效应的强弱也能制约其分布特点,加之根系状况及其分泌物也能施加影响,此外,微生物活动通过对有机质的降解来制约有机碳的分布[19-20]。通过对其土层分布差异的分析可以探究有机碳分布的动态变化,从而为促进局地碳平衡提供有益参考[21]。当土层深度低于20 cm的情况下,碳储量具有更明显的层次分布特点,其变化较大,而对于20—80 cm深度的土层而言,有机碳分布变化并不大,其稳定性较强,从而形成了鲜明的对比。与深层土壤相比,浅层土壤的碳密度更高,主要原因在于凋落物的分解效应增加了碳的积累,根系分布较为发达,有利于碳的积累;随着土层深度的不断增加,其活性碳分布越来越低,这与浅层土壤形成鲜明对比,最关键的影响要素在于凋落物的分解效应,在增强有机质的同时带来了更高水平的活性碳。借助于相关分析可知,对于芦苇等4种群落分布来讲,无论是丰度、Wiener指数,还是Cody,Sorenson指数,均与有机碳分布密切相关,且其正向变化关系突出,对于有机碳储量分布亦是如此。借助于双因素分析得知,植被类型及时间均对有机碳产生了制约效应,且二者交互效应对有机碳分布亦产生了突出影响,这种影响程度均达到了0.05的显著性水平;且其对丰度、Wiener指数均产生了制约效应,并且植被类型、年份及其交互作用对于物种群落分布施加着难以忽视的制约效应。
4 结 论
(1) 洞庭湖湿地植物群落多样性呈逐渐增加趋势,大致表现为:芦苇群落>苔草群落>水蓼群落>荫草群落;对于Simpson优势度指数,则表现为相反的变化趋势,具体表现为芦苇群落<苔草群落<水蓼群落<荫草群落。以上说明洞庭湖湿地植物群落多样性呈逐年增加的趋势,也表明近几年研究区域整体环境条件逐年好转。
(2) 土壤有机碳含量和有机碳储量均呈一致的变化规律,其中以表层土壤最高,随土层深度的增加逐渐降低;随剖面深度的增加,土壤有机碳储量逐渐降低,以表层土壤(0—20 cm)有机碳储量最高,呈典型的“表聚性”分布;通过各指标相关性分析表明:荫草群落、水蓼群落、苔草群落、芦苇群落Margalef丰富度指数、Shannon-Wiener多样性指数、Cody指数和Sorenson指数与有机碳含量和有机碳储量呈显著或极显著的正相关,从相关系数绝对值来看,与有机碳含量的相关系数高于有机碳储量的相关系数。由此说明洞庭湖湿地植物群落多样性的增加主要原因在于土壤有机碳含量的提升。
(3) 双因素分析表明植被类型和年份对有机碳含量和有机碳储量具有显著的影响(p<0.05),植被类型×年份对有机碳含量具有显著的影响(p<0.05);植被类型、植被类型×年份对Margalef丰富度指数(S)和Shannon-Wiener多样性指数(H)具有显著的影响(p<0.05);植被类型、年份对Cody指数和Sorenson指数具有显著的影响(p<0.05)。由此说明随年份变化,植被类型的变化是影响土壤有机碳含量和有机碳储量的主要因素。
