藏族和夏尔巴人15 个常染色体STRs 的遗传多态性
2020-10-17张致英范小伟赵一多马利锋康龙丽
张致英,范小伟,贺 娜,张 瑶,赵一多,刘 芳,马利锋,康龙丽
(1.西藏民族大学 高原相关疾病分子遗传机制与干预研究省级重点实验室,陕西 咸阳 712082;2.西藏民族大学高原环境与疾病相关基因研究高校重点实验室,陕西 咸阳 712082)
青藏高原是世界上最高的高原,平均海拔超过4000 m.人类学和遗传学研究已经证实,生活在西藏的世居人更容易适应高原低氧的环境[1-2].西藏自治区作为我国西南边陲省份,生活着藏族、蒙古族、维吾尔族、夏尔巴人等20 多个人群,各群体之间有着不同的历史背景和地理环境.藏族是青藏高原的原住民,夏尔巴人至今仍属我国未识别民族之一,对于藏族人和夏尔巴人的起源问题至今仍存有争议.到目前为止,Y染色体和线粒体DNA 的研究已经表明,现代藏族人的起源可以追溯到新石器时期的移民潮或者来源自黄河中上游流域[3-5],或者来源于北亚人群[6].夏尔巴人最初是西藏藏族人,通过对夏尔巴人历史和语言的研究发现,在500 年前部分藏族人从西藏东部移居到尼泊尔的珠穆朗玛地区,形成夏尔巴人[7].
短串联重复序列(short tandem repeat,STRs)又称为微卫星DNA,是一类广泛存在于人类基因组中的DNA 多态性基因座.该基因具有片段短、扩增效率高、判型准确等特点,被广泛应用于法医学个体识别、人类群体遗传学、多基因分子疾病诊断等领域.本研究选取了406 例藏族人和夏尔巴人,对15 个常染色体STRs 进行分析以探究这2 个群体的遗传多样性,并分析二者的亲缘关系和基因融合水平.同时将这2 个人群与世界各地不同人群进行全面比对分析,以期更好地了解藏族人和夏尔巴人的起源.
1 研究对象和方法
1.1 研究对象
本研究共采集406 份血液样本,包括西藏日喀则定日县的225 份藏族人血液样本和日喀则聂拉木县的181 份夏尔巴人血液样本. 采样遵循知情同意原则,收集的样本来自于3 代均为同一民族并且无血缘关系的健康个体.此项研究经过西藏民族大学伦理委员会批准同意.
1.2 DNA 的提取、PCR 扩增和STRs 分型
样本DNA 的提取采用Walsh 的Chelex 法,直接从白细胞中获取[8].15 个常染色体STRs 基因座具有很高的遗传标记价值,被许多国家运用在法医鉴定中,包括D8S1179、D21S11、D7S820、CSF1PO、D3S1358、TH01、D13S317、D16S539、D2S1338、D19S433、vWA、TPOX、D18S51、D5S818 和FGA.在GeneAmp 9600PCR 仪上,使用AmpF STR Identifiler PCR 扩增试剂盒(美国ABI公司)并按照产品说明进行扩增. 使用PRISM 3100 Avant DNA 分析仪(美国ABI 公司)对STR 产物进行分析,用GeneScan 3.7 和Genotyper 3.7 分析软件进行STR 的基因分型.
1.3 统计分析
应用Cervus 3.0 软件[9]进行等位基因频率、法医学参数(Ho,观察杂合度;He,期望杂合度;PIC,多态性信息含量;DP,个体识别能力;EPP,非父排除率;TDP,累积个体识别能力;CPE,累积非父排除率)以及哈迪-温伯格平衡检验的计算.用Arlequin 3.5 软件[10]计算和比较高原人群及世界范围内其他一些人群的遗传距离Fst、平均数差异、分化指数、共祖系数、Slatkins 线性Fst.用Structure 2.3.4 软件[11]推断人群结构并且将个体与预测人群进行匹配.用R 软件包和Distruct 1.1 软件[12]绘制遗传距离和群体遗传结构矩阵图.
40 个人群包括:夏尔巴人(Sherpa)、西藏定日藏族(Tibetan_Tingri)、西藏拉萨藏族(Tibetan_Lhasa)、西藏昌都藏族(Tibetan_Changdu)、青海高原藏族(Tibetan_Qinghai)、云南彝族(Yi_Yunnan)、珞巴族(Lhoba)、西藏僜人(Deng)、甘肃汉族(Han_Gansu)、陕西汉族(Han_Shaanxi)、江苏汉族(Han_Jiangsu)、云南汉族(Han_Yunnan)、广东汉族(Han_Guangdong)、东乡族(Dongxiang)、甘肃回族(Hui_Gansu)、宁夏回族(Hui_Ningxia)、喀什维吾尔族(Uygur_Kashi)、伊犁维吾尔族(Uygur_Yili)、俄罗斯族(Russian Inner-Mongolia)、朝鲜族(Korean)、畲族(She)、傣族(Thai)、葡萄牙人(Portuguese)、弗兰德人(Flemish)、希腊人(Greek)、罗马尼亚人(Romanian)、西班牙人(Spanish)、巴基斯坦人(Pakistan)、摩洛哥柏柏尔人(布里亚)(Berber_Morocco1(Bouhria))、摩洛哥柏柏尔人(阿斯尼)(Berber_Morocco2(Asni);)、埃及柏柏尔人(Berber_Egypt)、埃及穆斯林(Muslim_Egypt)、埃及科普特人(Copt_Egypt)、也门人(Yemenese)、沙特阿拉伯人(Saudi Arabian)、伊朗人(Iranian)、科威特人(Kuwaiti)、阿曼人(Omanis)、纳米比亚奥万博人(Ovambo_Namibia)、坦桑尼亚人(Tanzan).
2 结果与分析
2.1 藏族和夏尔巴人15 个STRs 等位基因的频率分布
西藏藏族人和夏尔巴人的15 个常染色体STRs等位基因的频率分布分别如表1 和表2 所示. 在藏族群体中15 个STRs 位点共检测出141 个等位基因,频率分布在0.0022~0.6222 之间. 在夏尔巴人群体中15 个STRs 位点共检测出139 个等位基因,频率分布在0.0028~0.5691 之间. 15STRs 的法医学参数如表3 所示. 在藏族群体中,TPOX 位点的PIC 低于0.5(0.470)且DP 低于0.8(0.528),D3S1358 位点的DP低于0.8(0.795),TH01 位点的DP 低于0.8(0.715),其余位点的DP 均高于0.8,PIC 均高于0.5. 在夏尔巴人群体中,除D18S51(P = 0.0361)和FGA(P =0.0016)基因座外,其余基因座均符合哈迪-温伯格平衡(P>0.05).

表1 藏族人群15 个常染色体STRs 基因频率分布Tab.1 Allele frequencies of 15 autosomal STRs in Tibetans

表2 夏尔巴人15 个常染色体STRs 基因频率分布Tab.2 Allele frequencies of 15 autosomal STRs in Sherpas

表3 藏族人和夏尔巴人的法医学参数Tab.3 Forensic parameters for Tibetans and Sherpas
2.2 遗传进化分析
中国西藏日喀则藏族人和夏尔巴人与其他已知群体间的遗传距离如图1 所示.由图1 可知,中国日喀则藏族人及夏尔巴人与其他群体配对比较的遗传距离值在0.00315~0.04203 之间. 与其他人群相比,中国日喀则藏族人、夏尔巴人和中国西藏拉萨人群间的遗传距离较短(Fst=0.00315),其次是汉族人,而遗传距离最远的是纳米比亚人(Fst=0.04203).僜人、畲族人和中国其他群体间的遗传距离Fst 均在0.02 以上.中国日喀则藏族、夏尔巴人和中国的其他群体(包括来自其他地区的藏族人、汉族人)之间的遗传距离远小于遗传距离最大的非洲群体,其次是西亚群体.

图1 藏族和夏尔巴人与世界范围其他人群配对FstFig.1 Plots of pairwise Fst of Tibetans and Sherpas and other worldwide populations
本研究使用了一种基于遗传模型的聚类方法,将藏族、夏尔巴人和其他人群之间的遗传相似性进行可视化[13].中国日喀则藏族人和夏尔巴人以及世界范围内其他一些人群的遗传结构如图2 所示.
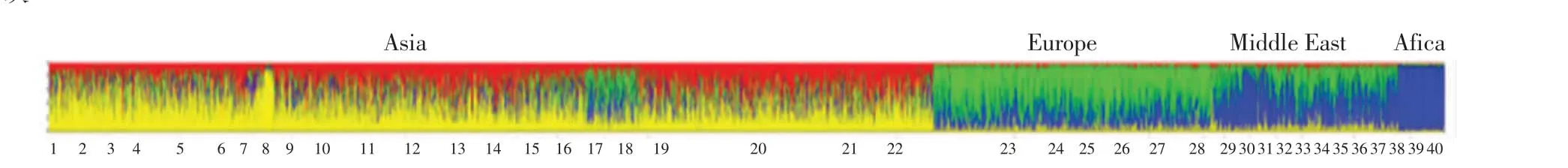
图2 西藏和夏尔巴人与世界范围内其他人群的遗传结构Fig.2 Estimated population genetic structure of Tibetan and Sherpas,and other worldwide populations
由图2 可以看出,中国藏族、中国夏尔巴人、中国汉族和中国朝鲜族具有显著的遗传相似性,而喀什和伊犁地区的维吾尔族与欧洲及中东群体具有显著的遗传相似性.中国藏族、中国夏尔巴人与非洲群体遗传相似性极不显著,这与本研究中遗传距离分析的结果较为一致.
3 讨论
STRs 群体遗传多态性可以反映群体的分化历史.本研究对西藏自治区日喀则藏族和夏尔巴人合计406人进行了15 个常染色体STR 基因分型,在藏族人群中共检测出141 个等位基因,频率分布在0.0022~0.6222 之间,在夏尔巴人群中共检测出139 个等位基因,频率分布在0.0028 ~0.5691 之间.国内外学者对STRs 位点遗传多态性的评价有不同的标准:Gill 等[14]认为DP≥0.9 的基因座具有高鉴别力,而国内学者[15-16]认为当DP >0.8 时,该基因座具有高度多态性遗传标记位点,有较高的应用价值,PIC >0.5 时遗传标记具有高度的可提供信息性.本研究中,藏族人群仅TPOX位点的PIC 低于0.5(0.470),DP 低于0.8(0.528),D3S1358 位点的DP 低于0.8(0.795),TH01 位点的DP低于0.8(0.715),其余位点的DP 均高于0.8,PIC 均高于0.5.在夏尔巴人群中除D18S51(P=0.0361)和FGA(P=0.0016)基因座外,其余基因座均符合哈迪-温伯格平衡(P>0.05).由此认为,藏族人群中TPOX、D3S1358、TH01位点和夏尔巴人群中D18S51、FGA 位点的多态性不高,其余位点均属于高度多态性遗传标记,这15个位点复合扩增后结合使用,具有很高的鉴别力,可用于法医学个体识别、亲子鉴定和群体遗传学研究等领域[17].
夏尔巴人被称为高原登山者,但对于夏尔巴人的起源仍有争议.一项通过夏尔巴人基因型数据分析结果表明,夏尔巴人和汉族人是现代藏族人的2 个祖先群体[18].也有研究通过对Y 染色体和mtDNA 多样性进行分析表明,夏尔巴人与藏族有着密切的亲缘关系,藏族人是夏尔巴人的祖先[7].最近的一项研究也认为藏族人是夏尔巴人的祖先[19].本研究从常染色体遗传角度对他们的遗传起源进行分析,结果表明,中国藏族和夏尔巴人与汉族及其他东亚群体具有显著的遗传相似性,而与欧洲或中东群体的遗传相似性不显著.
此外,本研究还发现僜人形成了一个独特的群体,这可能是由于隔离和遗传漂变的影响,因为他们是以狩猎和采集为生的数量较小的群体[20].此前研究认为,维吾尔人来源自欧亚人群,但从遗传学的角度来看,维吾尔人是东亚和欧洲人群结合交流而来[21-22].中东群体似乎是源自欧洲和非洲人群,这与Yao 等[23]的遗传结构分析结果一致.
总之,本研究的数据提供了更多关于藏族和夏尔巴人遗传起源的信息.研究结果表明,藏族和夏尔巴人与其他东亚人群在遗传上有着高度的相似性.
