植物萜烯类合成的转录调控研究进展
2020-10-16韩婷,杜方
韩 婷,杜 方
(山西农业大学园艺学院,山西太谷030801)
萜烯类物质是植物体内一种重要次生代谢物,是花香挥发物的重要成分[1],也是花器官引诱昆虫授粉或自我保护[2]的主要物质,具有较高的生态价值[3]。研究表明,萜烯类物质具有药用价值,如抗炎[4]、防腐杀菌[5]、抗疟疾[6]、抑制肿瘤活性等。但植物体内萜烯类化合物合成量较少,难以满足药用等需求,所以利用基因工程提高植物萜烯类物质的产量很有必要。多数植物体内萜烯类生物合成途径已基本明确,其中有多个限速酶且限速步骤难以确定,通过转录因子(Transcription factor,TF)激活或抑制萜类合成酶基因的表达可以促进萜烯类物质的合成,因此,萜烯类转录因子调控机制成为研究热点。本文综述了不同萜类代谢的转录调控研究现状,旨在更好地促进萜烯类物质的转录调控研究。
1 萜烯类物质及相关转录因子
萜烯简称萜,广泛存在于植物体内,按萜类分子中包括异戊二烯单位的数目可分为单萜、倍半萜、二萜、二倍半萜、三萜、四萜、多萜,常见的萜类物质如表1所示,单萜和倍半萜物质较多,其他萜类较少。

表1 常见的萜类物质分类
萜类化合物的合成是一个复杂的过程,主要是以异戊烯二磷酸作为底物,通过相应的烯萜合酶的催化作用合成[7]。异戊烯基焦磷酸(isopentenyl pyrophosphate,IPP) 和 3,3- 二甲基丙烯基焦磷酸(dimethylallyl pyrophosphate,DMAPP)是类异戊二烯合成的底物,如图1[8]所示,它们的生成主要通过2条代谢途径:一条是发生在细胞溶质中的甲羟戊酸(MVA)途径,该途径参与倍半萜、三萜等次生代谢产物的生物合成;另一条是发生在质体中的2-C-甲基-D-赤藻糖醇-4-磷酸(MEP)途径,主要参与单萜、二萜等物质的生物合成[9]。
萜类物质的释放部位包括叶片、根、花瓣和果实(表 2)。释放量随品种[10]、发育时期[11]、组织[12]等的不同而发生变化。萜类合成酶基因的表达量直接影响萜类物质的释放量,而萜类合成酶基因的表达又受到转录因子的调控,通过激活[13]或抑制[14]萜类合成酶基因的表达可以影响萜类物质的释放。如今越来越多的研究由表及里、由浅入深,从影响表型的外部因素转到内部分子调控机制,转录因子已成为萜类研究的重要线索。

表2 植物萜类释放相关信息
转录因子是能与基因5′端上游特定序列专一性结合,从而保证目的基因在特定的强度时空下表达蛋白质。从蛋白质结构分析,TF由DNA结合区、转录调控区、寡聚化位点以及核定位信号共4个功能区域组成,通过这些功能区域与启动子顺式元件结合或与其他转录因子的功能区域互作[29]。研究转录因子一般借助酵母单双杂、烟草叶片的瞬时表达、Gus染色以及双荧光素酶分析等试验进行。常见的调控萜类合成机制的TF家族有6个,包括AP2/ERF、bHLH、WRKY、MYB、NAC和 bZIP[30]。
2 植物单萜转录调控研究
研究发现,不同种类植物单萜受不同转录因子家族调控,表3给出了部分植物及其调控单萜的转录因子。PbbHLH4和PbbHLH6过表达可以增加蝴蝶兰单萜物质的释放量,香味显著增强[24]。bHLH类[31-34]、AP2/ERF 类[35-37]、WRKY 类[38]转录因子与单萜吲哚生物碱途径相关基因启动子结合后可促进长春花长春质碱或者蛇根碱的合成。AP2/ERF、bHLH、MYB、NAC、ARR-B、ARF、MIKC-MADS、HDZIP、G2-like、C2H2和CPP共11个家族的转录因子[39]都可能影响佛手柠檬烯等单萜物质的释放量。CitERF71激活萜类合成酶基因CiTPS16表达从而促进甜橙香叶醇的释放[28]。LiMYB、LibZIP、LiGeBP、LiSBP-2、LiERF-1和LiERF-2等转录因子可能会影响薰衣草中单萜物质芳樟醇和1,8-桉树脑的合成[40],而小叶章薰衣草中 AP2-EREBP、bHLH、WRKY、MYB、MADS等9个转录因子[41]可能与单萜芳樟醇和乙酸芳樟酯释放相关,推测不同的品种间释放的单萜物质不同可能是受不同转录因子调控的结果。除了上述正调控因子之外,植物中也存在一些负调控因子,如对长春花中AP2/ERF类转录因子CR1沉默后,可以促进文多灵和蛇根碱的释放量上升[42];bZIP家族的转录因子CrGBF1和CrGBF1会抑制Str基因的表达,从而降低长春碱的合成[14]。此外,大豆[43]和绿薄荷[44]中均存在MYB家族的抑制型转录因子,分别抑制长春质碱和松萜的含量。
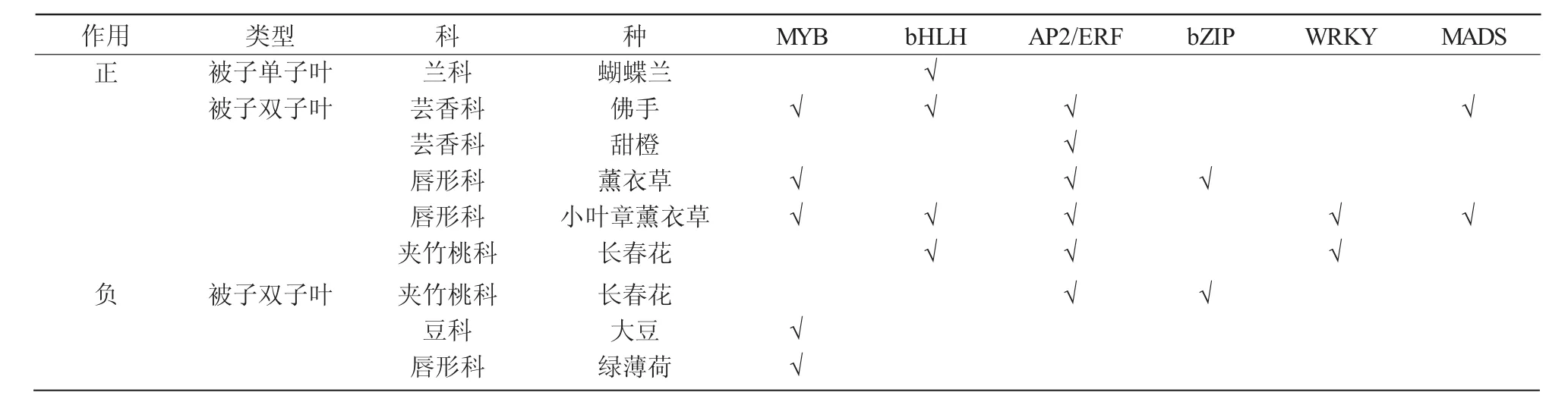
表3 部分植物单萜调控中的转录因子
3 植物倍半萜转录调控研究
青蒿素是倍半萜中的主要研究物质,由于ABA、JA、SA均会提高青蒿素的含量,所以在其试验中一般与这些生长调节剂相结合,共同分析基因功能。表4给出了调控倍半萜的转录因子,研究表明,包括 MYB 类[45-46]、AP2/ERF 类[47-49]、bZIP 类[50]、WRKY 类[51-52]、bHLH 类[53]、SPL 类[54]、NAC 类[55]等不同家族的转录因子能够激活青蒿素合成途径基因的表达,提高青蒿素的产量。MYB类转录因子与SICOS1基因共表达,可能会在和青蒿同为菊科的雪莲木香烃内酯合成途径中发挥重要的作用,但具体调控作用仍有待进一步研究[56]。R2R3-MYB转录因子抑制MsGPPS表达从而抑制绿薄荷倍半萜的含量[44]。bHLH转录因子MYC2可以直接与TPS21和TPS11基因的启动子结合从而提高拟南芥倍半萜含量,尤其是(E)-β- 石竹烯的含量[57]。δ- 杜松烯合酶(δ-cadinene synthase,CAD1)是一种倍半萜环化酶,能合成棉酚,而转录因子GaWRKY1能结合CAD1启动子中的W-box调控棉花棉酚的含量[58]。过表达转录因子HAC1能够增加毕氏菌株倍半萜物质橙花醇的含量[59]。转录因子除了单独作用,也可能存在不同家族转录因子进行共同激活或抑制基因,从而调控植物萜烯类的释放量。在细胞中过表达TwTGA1时增加了倍半萜吲哚生物碱,茉莉酸甲酯处理进一步增强了生物碱的含量,但是对TwTGA1进行RNA干扰,并没有显著影响生物碱的产量,说明还存在其他具有重叠功能的TGA[60],可能存在转录因子的共表达。

表4 部分植物倍半萜调控中的转录因子
4 植物二萜转录调控研究
二萜中丹参酮与植保素的研究比较广,表5给出了调控二萜合成的转录因子家族类型,MYB家族、WRKY家族以及bHLH家族影响丹参中的丹参酮的生成,过度表达SmbHLH10[61]、SmMYB9b[62]、SmWRKY2[63]、MYB36[64]会激活丹参酮生物合成途径相关基因的上调表达,从而增加丹参毛状根中丹参酮的含量,而SmbHLH3则会抑制丹参酮的产生[65];bZIP转录因子OsTGAP1激活OsDXS3基因的表达,过表达后导致水稻二萜类植物抗毒素的超积累[66-67];部分WRKY和bZIP家族的转录因子对植保素合成途径也有抑制作用,过表达OsWRKY76[68]、OsbZIP79[69]可以减少水稻植保素的产生。对于二萜的转录调控研究主要集中在bHLH类和WRKY类转录因子上,对于其他转录因子可能的转录调控机制研究较少。单萜和二萜主要是来自MEP途径,在调控机制等方面存在相似性,如均存在bZIP家族的负调控转录因子。

表5 部分植物二萜调控中的转录因子
5 植物三、四萜转录调控研究
对于三萜、四萜的研究还不是非常深入,表6给出了调控植物三、四萜的转录因子。据报道,三、四萜合成途径关键酶主要通过bHLH家族来进行调控,bHLH家族转录因子Bl和Bt均可以促进Bi基因的表达从而分别在黄瓜叶子和果实中提高葫芦素的含量[70];bHLH家族的TSAR1和TSAR2通过直接与HMGR1启动子上的N-box结合来促进蒺藜状苜蓿三萜皂苷的合成[71];BpbHLH9会促进白桦中桦木酸的合成[72];GubHLH3积极地调控大豆皂苷生物合成基因CYP93E3和CYP72A566,从而促进乌拉尔甘草大豆皂苷等三萜及其皂苷生物合成[73]。四萜研究主要集中于类胡萝卜素,R2R3-MYB类转录因子MYB7被发现激活AdLCY-b基因的启动子从而增加美味猕猴桃类胡萝卜素的含量[74],ERF转录因子SlAP2a[75]、SIERF6[76]减少了番茄类胡萝卜素的积累量。三、四萜类物质研究较其他萜类少,相关转录因子种类较少,主要有MYB、AP2/ERF及bHLH这3类,且研究方向大多集中在果树及蔬菜方面。

表6 部分植物三、四萜调控中的转录因子
6 结论
萜烯类物质作为一种重要的植物次生代谢物,在植物生长发育、医学、化妆品、食品等方面都具有不可缺少的作用,但其天然合成量较少。目前主要借助基因工程提高萜烯类物质的产量,但对其调控机制尚未明确,通过转录因子来调节基因表达量从而加大萜烯类物质合成量就显得很有价值。本文从单萜、倍半萜、二萜、三四萜这4个方面综述了其生物合成调控所需的转录因子。萜类物质不同,调控其产量的转录因子各有不同,但基本都属于同一转录因子家族,MYB、bHLH以及WRKY在萜类研究中最为常见。AP2/ERF类在萜烯类合成途径中也起着重要的调控作用。萜类物质的合成也会受转录因子共表达的调控。目前研究多集中在双子叶植物萜类合成调控机制,且多集中于唇形科植物,可能是因为唇形科植物含有芳香油且具有一定的药用价值,但是对可能存在潜在开发价值的单子叶植物(如百合及蝴蝶兰)研究甚少,且对转录因子作用强度的鉴定也不透彻。
随着植物基因组学的发展以及研究体系的逐渐完善,越来越多的萜烯合成途径相关基因及转录因子将被鉴定,萜烯类物质所延伸的领域势必越来越广。
