太行鸡胚PRLR基因表达与主翼羽和覆主翼羽长关系
2020-10-15杜小龙张乐超赵丽杰李兰会李祥龙
杜小龙,张乐超,赵丽杰,乔 宁,高 明,李兰会,3*,李祥龙
(1.河北农业大学 动物科技学院,河北 保定071000;2.华裕农业科技有限公司,河北 邯郸056000;3.河北省牛羊胚胎工程技术研究中心,河北 保定071000;4.河北科技师范学院,河北 秦皇岛,066004)
羽型是判定雏鸡性别的重要表型性状之一,其中主翼羽和覆主翼羽长度差异是在鸡胚生长过程中形成[1]。最早研究发现慢羽K基因和禽内源白血病毒ev21存在连锁关系,并且普遍认为ev21病毒插入导致慢羽表型的形成[2-3];后续研究重新定位K基因为包含催乳素受体(PRLR)在内的基因重复[4-7],2015年本课题组验证了PRLR基因的部分重复序列是导致羽型差异的分子基础,并且发现ev21与K基因并非紧密连锁[2,8],TAKENOUCHI等[9]也发现ev21整合不是慢羽的分子基础。PRLR基因定位在鸡Z染色体,其编码蛋白催乳素受体PRLR包含1个跨膜域[10-13]。PRLR与其配体催乳素PRL结合可以影响甲状腺激素分泌和NaCl再吸收等从而对鸡胚的发育起关键作用[14],有研究表明PRLR抑制毛囊发育[2,15]。PRLR基因转录产生多种剪切体,编码不同类型的膜结合型蛋白,其主要区别为胞外域的组成成分和序列的长度[2,16]。鸡PRLR的Ⅰ型剪切体(PRLRS1),受组织特异性启动子P1调控,主要在成年鸡小肠和肾表达,而种属特异启动子P2调控的Ⅱ型剪切体(PRLRS2)在成年鸡和胚胎组织中广泛表达,并且P2启动子与人、大鼠和小鼠广泛表达的PRLR通用启动子PⅢ具有高度同源性[17]。PRLR多种转录剪切体与其信号传导途径的多样性以及配体功能多样性、组织表达的广泛性相适应。在不同组织通过不同转录产生剪切体,发挥其各自功能[2,18]。
本研究对快慢羽太行鸡胚发育过程中翅羽皮肤组织PRLR基因的不同剪切体表达进行分析,并检测快慢羽鸡胚主翼羽和覆主翼羽长度变化,以揭示羽型形成与PRLR基因表达调控间关系,为快慢羽鸡主翼羽和覆主翼羽长度差异形成研究提供参考。
1 材料与方法
1.1 试验材料62枚18胚龄和67枚19胚龄太行鸡胚翅膀皮肤组织、主翼羽和覆主翼羽、肝脏均采集于河北农业大学太行鸡孵化室;Easy Pure Genomic DNA Kit、TransZolTMUp PlusRNAKit、TransScriptROne-Step gDNA Removal and cDNA Synthesis SuperMIX、Trans 2K Plus DNA Marker均购自北京全式金公司;Super GelRedR、2×ES Taq MasterMix(Dye)购自保定康为世纪公司;引物合成及测序由华大科技公司完成。
1.2 主翼羽和覆主翼长度测量分别收集鸡胚右翅由内向外第1,2,3根主翼羽和覆主翼羽,测量长度(mm),3根翅羽长度的平均值作为主翼羽和覆主翼羽的长度。
1.3 DNA、RNA提取及cDNA合成使用Easy Pure Genomic DNA Kit、TransZol TM Up Plus RNA Kit分别对肝脏、翅膀皮肤组织进行处理,提取DNA及总RNA。使用反转录试剂盒Trans ScriptROne-Step gDNA Removal and cDNA Synthesis Super MIX将RNA合成cDNA。
1.4 太行鸡胚羽型和性别检测参考张秀玲等[6]试验结果,合成羽型半定量鉴定引物1305和857;参考胡锐颖等[19]合成性别鉴定引物450-650。羽型和性别检测扩增体系为:DNA 0.5 μL,Mix 5 μL,上、下游引物(10 μmol/L)分别0.5 μL,ddH2O补足10 μL。PCR程序:95℃ 5 min;95℃ 30 s,退火30 s,72℃延伸 1 min 30 s,23~35个循环;72℃延伸10 min(表1)。
1.5 PRLR剪切体半定量检测
1.5.1引物设计 NCBI下载PRLR的3种剪切体序列。Ⅰ型剪切体(PRLRS1,NM_204854.1)包含外显子3(133 bp),缺少外显子8(121 bp);Ⅱ型剪切体(PRLRS2,GQ184574.1)缺少外显子3和8;Ⅲ型剪切体(PRLRS3,JX560222.1)有外显子3和8。利用Primer Premier 5.0设计引物PR2和PR3,并用NCBI数据库检测引物特异性(表1)。
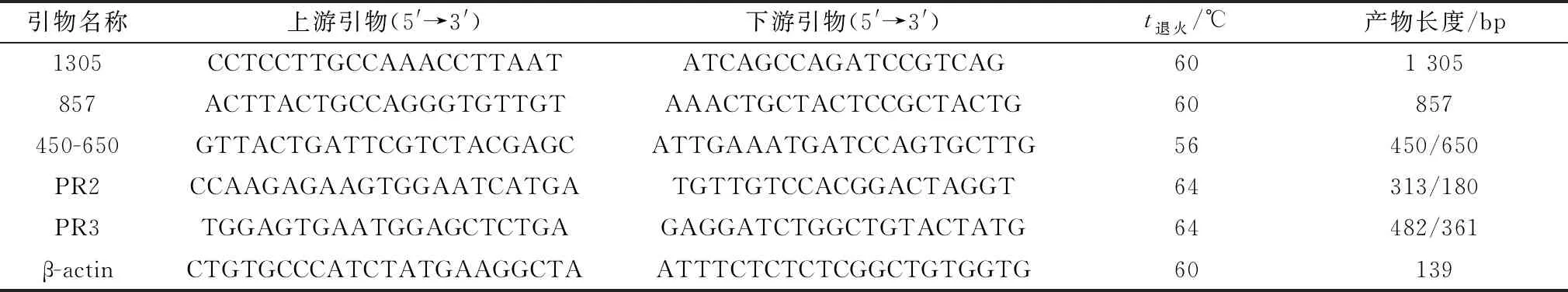
表1 试验用引物信息
利用引物PR2和PR3进行PCR扩增,检测鸡胚翅皮肤组织PRLR剪切体存在类型(图1)。PR2扩增得到180 bp产物表明PRLRS2表达,得到313 bp 产物说明PRLRS1或/和PRLRS3表达;PR3扩增得到361 bp产物表明PRLRS1或/和PRLRS2剪切体表达,得到482 bp说明PRLRS3有表达。
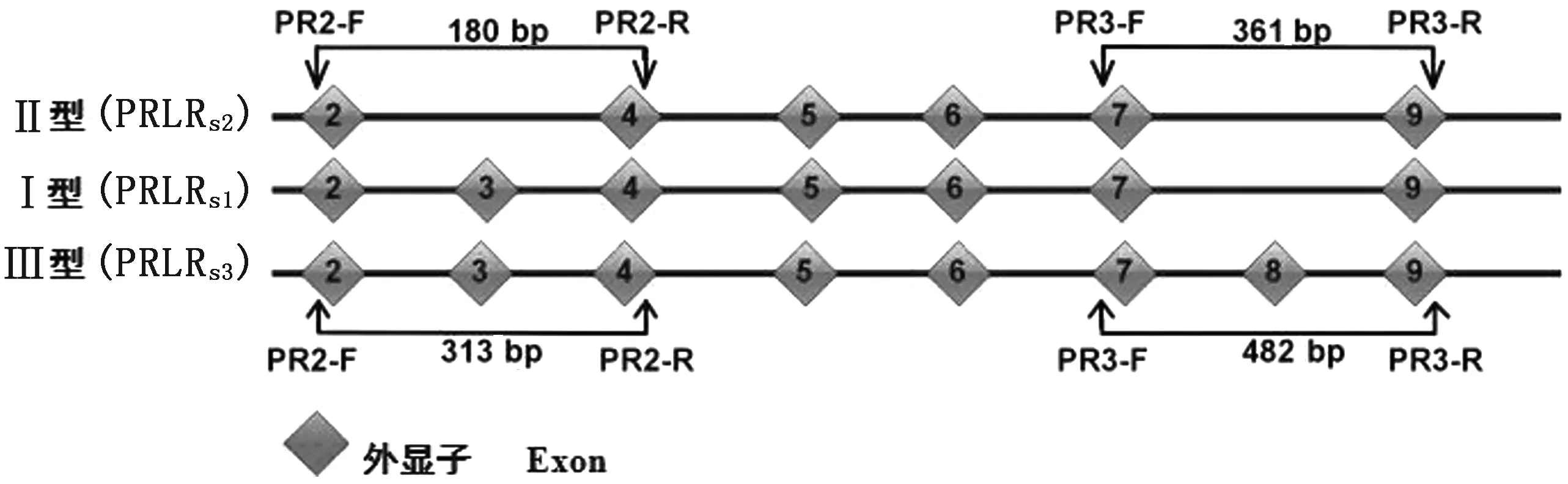
图1 PRLR剪切体与引物示意图
1.5.2PRLR剪切体表达的半定量分析 将目的片段引物设置退火温度梯度或循环次数梯度,确定引物扩增到达平台期前的最适退火温度及循环数,与β-actin内参引物在同一条件下同时扩增。分别取2 μL 扩增产物使用1.5%琼脂糖凝胶电泳,每个样本的目的产物与β-actin内参产物在同一胶孔,通过产物大小确定鸡胚翅皮肤组织PRLR剪切体类型,使用Image J软件检测分析内参条带与目的条带灰度值(亮度),以两者的灰度值比值来表示不同剪切体相对表达量。
PCR反应体系:0.4 μL cDNA,2×Es Taq MasterMix 5 μL,上下游引物(10 μmol/L)各0.4 μL,无核酸酶纯水补足至10 μL。
PCR扩增条件:94℃预变性5 min;94℃变性30 s,退火60℃ 30 s,72℃ 10 s,循环35次;72℃ 延伸10 min;4℃保存。
1.6 数据分析采用SPSS19.0一般线性模型分析胚龄、性别和羽型因素对主翼羽和覆主翼羽长和PRLR剪切体表达影响;单因素方差分析性别和胚龄对不同羽型鸡羽长变化和PRLR基因剪切体表达变化,组间进行Duncan多重比较,α=0.05为影响显著。
2 结果
2.1 DNA、总RNA提取及cDNA合成DNA、RNA和PCR产物经1%琼脂糖电泳检测,可见条带致密、整齐、清晰无拖尾,PCR扩增特异性强,没有非特异性条带和引物二聚体出现,片段长度与预期一致,完全满足后期试验。图2为肝脏组织提取的基因组DNA、皮肤组织提取的总RNA和以cDNA为模板扩增内参基因β-actin 的凝胶检查结果。

图2 DNA(A)、RNA(B)和β-actin(C)凝胶电泳检测 A.太行鸡肝脏组织提取基因组DNA;B.皮肤组织提取总RNA;C.以太行鸡cDNA为模板扩增内参基因β-actin的凝胶检查结果。M.DL5000 DNA Marker;1~5.样品
2.2 羽型和性别鉴定羽型性别鉴定检测结果显示857 bp为快羽慢羽表型,均有GHR基因扩增条带;1 305 bp为慢羽鸡PRLR重复特异条带。PCR产物经1%琼脂糖电泳检测,可见条带清晰无拖尾,PCR扩增特异性强,没有非特异性条带和引物二聚体出现,片段长度预期片段长度一致,完全满足下一步试验(图3)。

图3 太行鸡羽型(A)和性别(B)鉴定 A.羽型检测结果(5,6,8.慢羽鸡PCR产物;1,2,3,4,7.快羽鸡PCR产物);B.性别鉴定结果(1,4.公鸡;2,3,5,6,7,8.母鸡)
2.3 主翼羽和覆主翼羽长度变化一般线性模型分析18胚龄(E18)和19胚龄(E19)不同性别快慢羽太行鸡胚的主翼羽和覆主翼羽毛长,结果显示胚龄和羽型对主翼羽和覆主翼羽长度影响的主效应均极显著存在(P<0.01),而性别不影响太行鸡主翼羽和覆主翼羽长度(P>0.05)(表2)。
进一步分析E18和E19的快慢羽太行鸡胚主翼羽和覆主翼羽毛长,结果显示快羽鸡18和19胚龄的主翼羽均长于覆主翼羽1.5 mm以上(P<0.05),而慢羽鸡主翼羽略长于覆主翼羽0.4 mm以上(P>0.05)。18和19 胚龄的快慢羽鸡胚主翼羽长存在显著差异(P<0.05),快羽和慢羽鸡胚的主翼羽长在18胚龄已经表现出显著差异,快羽鸡两个胚龄的主翼羽均显著长于慢羽鸡2 mm左右。快羽鸡18胚龄的覆主翼羽长长于慢羽鸡1 mm(P<0.05),而在19胚龄时,两羽型间覆主翼羽长度没有显著差异(P>0.05)。太行鸡快慢羽型间主翼羽在18~19胚龄已经表现并维持长度差异,覆主翼羽在18胚龄同样表现出长度差异,但19胚龄两羽型间覆主翼羽长度差异基本消失,快羽鸡覆主翼羽生长速度降低(表3)。
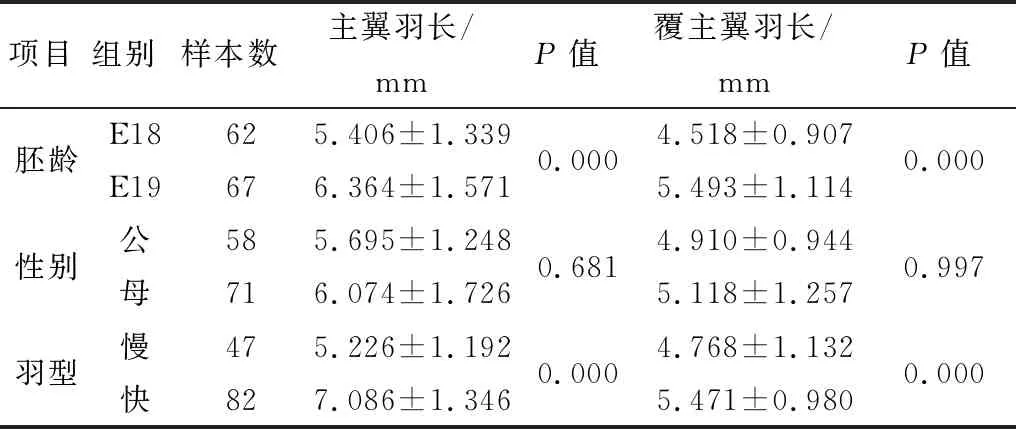
表2 胚龄、性别和羽型对太行鸡主翼羽和覆主翼羽长度主效应的影响
2.4 PRLR基因剪切体半定量结果129枚鸡胚翅羽皮肤中均检测到PRLRS1和PRLRS2剪切体的各自目的带313和180 bp以及与PRLRS1/PRLRS2共同表达目的条带361 bp,未检测到PRLRS3目的条带482 bp,说明太行鸡皮肤组织中存在PRLRS1和PRLRS2两种剪切体类型。利用目的条带313和180 bp 的灰度值分别与内参目的条带139 bp灰度值做比较,作为PRLRS1和PRLRS2剪切体的相对表达量(图4)。

表3 两种胚龄的快慢羽太行鸡的主翼羽和覆主翼羽长度变化 mm

图4 PRLR剪切体凝胶电泳检测 1~5中361 bp条带为PR3引物扩增产物;6~10中313 和180 bp条带为PR2引物扩增产物;1~10中139 bp 条带为β-actin引物PCR产物;M.DL5000 DNA Marker
2.5 羽型、胚龄和性别对PRLR剪切体表达影响胚龄、性别和羽型与PRLR两种剪切体表达量关系结果表明,18和19胚龄两种剪切体表达并没有显著性差异(P> 0.05),不同性别之间也没有显著性差异(P> 0.05),胚龄、性别对PRLRS1和PRLRS2表达都没有影响。但两种羽型间PRLRS2表达有显著差异(P< 0.05),说明PRLRS2表达可能是导致19胚龄后快慢羽羽长表型差异的主要原因,需进一步分析不同羽型的剪切体表达日龄变化(表4)。
2.6 两种羽型的PRLRS1、PRLRS2表达随日龄变化18和19胚龄剪切体PRLRS1表达量在快羽或慢羽系中并没有随时间推移发生显著性变化(P>0.05)(图5A);剪切体PRLRS2表达量在18胚龄快、慢羽系间也没有显著差异(P>0.05),但在19胚龄时慢羽表达量显著高于快羽(P<0.05)(图5B)。

表4 羽型、胚龄和性别对PRLR剪切体表达影响
3 讨论
鸡的羽毛颜色与其品种品系鉴定有关,在蛋鸡的配套系生产中利用羽型进行商品代性别鉴定,PRLR基因的重复与慢羽表型连锁,但是PRLR基因如何调控羽型形成尚不明确。羽型基因是伴性遗传基因,并影响羽毛发育脱换[20]。娄义洲等[21]发现武农I系乌骨鸡的主翼羽和覆主翼羽长度差异随生长阶段不同发生变化,第2周龄开始快慢羽鸡之间的毛长差异随着日龄增长而减小[22]。赵彩娟等[23]测定2~10周龄坝上长尾鸡翅羽长度,没有发现不同羽型公鸡2~10周龄主翼羽长度的显著差异,但是不同羽型母鸡的主翼羽长度具有显著差异。本课题组对18和19胚龄快慢羽太行鸡的主翼羽和覆主翼羽长度及PRLR基因表达进行研究,发现快羽鸡的主翼羽均长于慢羽鸡2 mm左右;18胚龄的快羽鸡覆主翼羽长于慢羽鸡,而19胚龄时快、慢羽鸡覆主翼羽长度没有差异,源于19胚龄时快羽鸡覆主翼羽生长速度降低。另外,两个胚龄快羽鸡主翼羽平均长于覆主翼羽1.7 mm左右,而慢羽鸡主翼羽仅长于覆主翼羽0.4 mm左右。认为是快羽太行鸡19胚龄覆主翼羽生长速度较慢羽鸡快、慢羽表型形成差异的原因之一。
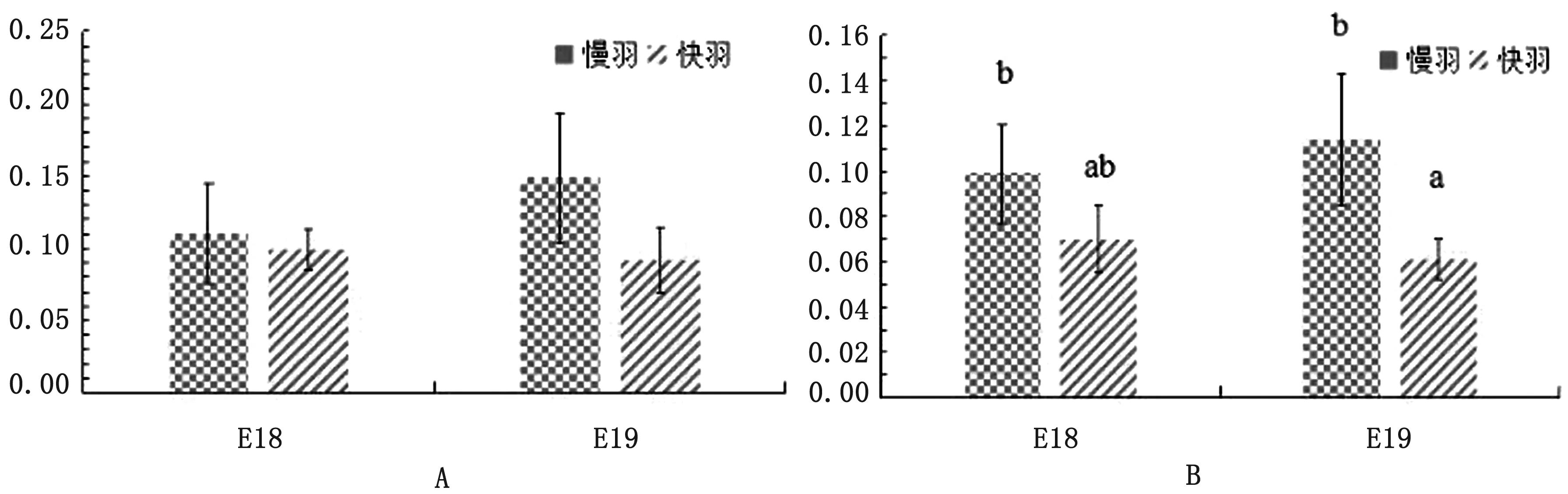
图5 两种胚龄两种羽型鸡胚皮肤组织PRLRS1(A)和PRLRS2(B)表达变化
另外,本研究发现在不同羽型太行鸡皮肤组织中不存在PRLRS3剪切体,这与BU等[24]和CHEN等[25]在鸡与鸽子上的研究结果一致。孵化期剪切体PRLRS1的表达在快慢羽间无显著差异,而PRLRS2在19胚龄的慢羽鸡表达显著高于快羽,这与19胚龄慢羽鸡的覆主翼羽生长速度的加快相对应。但剪切体PRLRS2在快慢羽鸡19胚龄的表达差异,是否影响覆主翼羽的生长变化,还需要深入细胞培养试验深入研究。冯宇等[26]发现慢羽鸡PRLR的mRNA表达高于快羽,但是蛋白表达无差异,BU等[24]发现PRLRS2与cPRL共转染不能激活HepG2细胞荧光素酶活性,说明PRLRS2在毛囊发挥作用的方式可能和PRLRS1不一样,是否PRLRS2可以和PRL结合需要进一步验证。垂体组织未形成的早期鸡胚中已有cPRL-L的广泛表达[27],PRL与PRLR相互影响,共同调控催乳素信号通路。在鼠和牛的研究中发现PRL与PRLR基因敲出或突变影响被毛的生长循环及其长度[28-30]。非哺乳动物家禽的PRLR基因表达变化,尤其是PRLRS2在慢羽鸡19胚龄的高表达可能与其调控覆主翼羽的生长有关。其中PRLRS2与PRL如何通过催乳素信号系统发挥羽毛生长作用,现在不得而知,因此关于太行鸡PRLR剪切体如何调控主翼羽和覆主翼羽生长还需进一步研究。
