卡介苗非特异性保护作用研究进展
2020-10-13陈柠孙琳申阿东何秋水
陈柠 孙琳 申阿东 何秋水
卡介苗(bacillus Calmette-Guérin)是最早研发的疫苗之一。1908年,法国科学家Albert Calmette和Camille Guérin将有毒的牛分枝杆菌培养于含胆汁、甘油、马铃薯等的培养基中,经过13年230次传代而获得的减毒活疫苗株,卡介苗由此诞生[1]。自1921年开始使用以来,截至目前,全球约有80%的国家将卡介苗列入国家计划免疫疫苗,累计接种40亿人,对于降低全球肺结核的发生起着至关重要的作用[2-3]。
自卡介苗开始使用后不久,一项在英国和美国开展的持续13年(1948—1961年)的临床随机试验表明,卡介苗接种可以降低儿童死亡率[4]。一项在西非进行的研究表明,12月龄儿童有卡介苗卡痕者的死亡率是没有卡痕者的41%,接种卡介苗可以将儿童感染性疾病的致死率降低43%[5]。卡介苗降低儿童死亡率的现象可能与卡介苗接种后能减少其他病原感染相关。近些年来,也有越来越多的研究表明,包括卡介苗在内的很多疫苗,除了对特定的病原体感染能起到保护作用外,对其他不相关的病原体感染也存在一定的保护作用,这被称为非特异性保护作用(non-specific protective effect)[6]。近期一项研究发现,卡介苗接种可以降低5岁以下儿童疟疾的发病率[7]。卡介苗免疫疗法是辅助治疗非肌肉侵入性膀胱癌的有效方法,虽然其机制尚不明确,但研究认为卡介苗诱导的非特异性保护作用是其中重要的一方面[3, 8]。除此之外,卡介苗可以降低白血病、肺癌等癌症发病的风险,也有动物实验表明,接种卡介苗可以减少糖尿病和阿尔兹海默症的发生[9-12]。笔者将着重对卡介苗对其他传染病的保护作用进行综述。
一、卡介苗对其他细菌感染的非特异性保护
卡介苗能诱导产生两类免疫反应,一类是基于抗原特异性的免疫反应,使其可以预防肺结核和非结核分枝杆菌(non-tuberculous mycobacteria,NTM)感染,另一类是依赖于细胞编程的“受训免疫(trained immunity)”,使其对肺结核以外的其他感染有一定预防作用[13]。卡介菌与NTM有相同或相似的抗原表位,所以卡介苗可能对NTM感染发挥着交叉保护作用[14]。已报道的与卡介苗有交叉保护作用的传染病包括麻风病、布鲁里溃疡等[3]。
(一)麻风病
麻风病由麻风分枝杆菌引起,经空气传播,主要损伤皮肤、外周神经、上呼吸道黏膜和眼睛,若不及时治疗将会对上述部位产生永久性损伤。根据WHO[15]统计,仅2018年,6个 WHO划分区域的159个国家共增加208 619例麻风病患者,患病率约为2/100 000。
关于卡介苗对麻风病的交叉保护作用,早在1939年就已被报道,自20世纪60年代开始的一些研究对麻风病的保护效果进行了比较系统的评估或随机临床对照试验[16],发现保护效果在20%~90%之间,这种差异可能与研究区域、人群、患者例数、疫苗菌株等不同有关(表1)。但总体来说,卡介苗对麻风病有一定的交叉保护作用。鉴于目前没有专门用于预防麻风病的疫苗,接种卡介苗是目前预防麻风病最有效的方法。WHO基于前期研究结果,建议在麻风病高发区域,在婴儿出生时就接种卡介苗[17]。对于不同卡介苗接种次数对麻风病的保护效果,研究结论也不太一致。最近的一项研究表明,与不接种卡介苗相比,接种两剂卡介苗对于麻风病患者家庭密切接触者感染麻风病有明显的保护作用,并且能够明显延长麻风病患者发病后的存活时间[18]。前期也有研究发现,接种两剂卡介苗对于麻风病的保护效果高于仅接种一剂,建议考虑在麻风病高发区域为麻风病患者密切接触者增加接种一剂卡介苗[16]。但是也有研究发现,为学龄儿童接种第二剂卡介苗对于预防麻风病并没有明显的作用[19]。
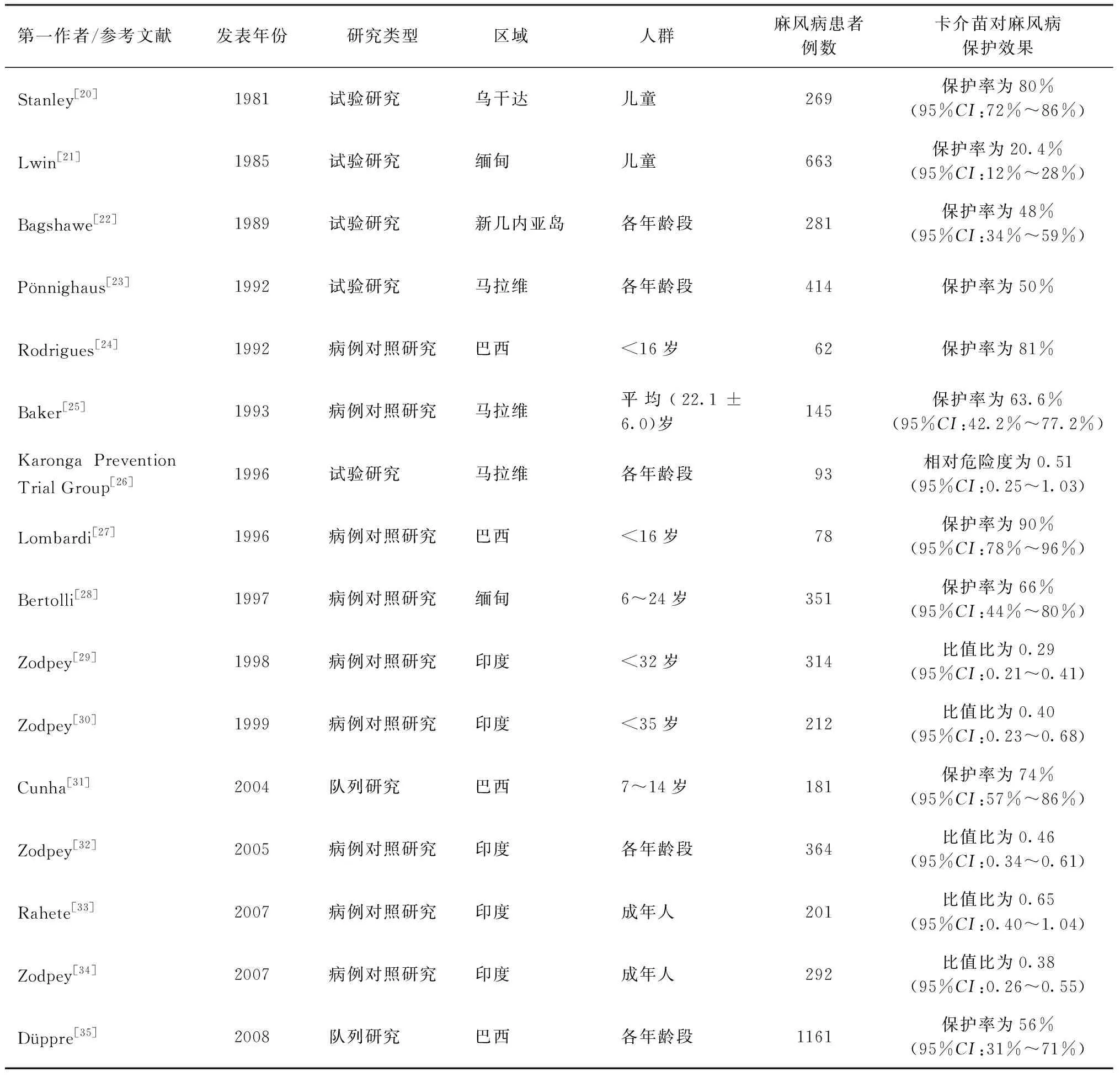
表1 卡介苗对麻风病非特异性保护的相关研究
(二)布鲁里溃疡
布鲁里溃疡(Buruli ulcer)是由溃疡分枝杆菌引起的一种慢性皮肤病,初期以皮肤丘疹、水肿为主要表现,最终发展为大面积的皮肤溃烂[36]。目前预防布鲁里溃疡除了在疫区减少高危行为(游泳、捕鱼等),卡介苗也表现出一定的保护作用,但是各个研究的结论存在差异。早在1969年发表的一篇在乌干达进行的研究表明,卡介苗对布鲁里溃疡的保护率在布鲁里溃疡高发区域为18%,在布鲁里溃疡低发区域为74%[37]。因此认为,卡介苗对降低布鲁里溃疡的发病率有一定作用。但是,2010—2013年在刚果、加纳、多哥3个国家进行的一项病例对照研究发现,卡介苗对布鲁里溃疡的发生或严重程度没有明显的保护作用[38]。鉴于目前没有完全有效的预防措施,WHO最新的卡介苗疫苗立场文件中建议在布鲁里溃疡高发区域考虑接种卡介苗[17]。
二、卡介苗对病毒感染的非特异性保护
关于卡介苗对病毒感染的非特异性保护研究较少,目前已报道的包括呼吸道合胞病毒、单纯疱疹病毒、流感病毒、黄热病毒和人乳头瘤病毒等(表2)。其中Stensballe等[39]对比呼吸道合胞病毒患儿和健康儿童卡介苗接种情况,发现呼吸道合胞病毒患儿未接种比例高于健康儿童,Arts等[40]发现,接种卡介苗可以降低黄热病毒疫苗接种后的病毒血症。其他还有一些研究是将卡介苗用于病毒性感染疾病的治疗,如Hippmann等[41]研究发现,卡介苗可以在短期内促进单纯疱疹病毒的清除,Podder等[42]发现,接种卡介苗可以有效治疗人乳头瘤病毒引起的疣。上述研究结果表明,卡介苗可以在一定程度上预防某些病毒感染或提高某些病毒性感染疾病的治疗反应。但是有些研究患者例数较少,不能有效代表群体的水平,如2015年一项关于流感病毒的研究中,卡介苗接种组和安慰剂接种组均为20例,2016年关于人乳头瘤病毒的病例系列报告仅为7例[43-44]。
对于目前在全球肆虐流行的新型冠状病毒肺炎(corona virus disease 2019,COVID-19)病毒,荷兰、英国、德国和澳大利亚4个国家将于近期启动一项临床试验,试图证明卡介苗对其是否有预防保护作用[45]。Curtis等[46]认为接种卡介苗可能可以降低COVID-19暴露后的病毒血症,从而减轻其症状并促进其康复,而Hamiel等[47]研究了以色列COVID-19病毒PCR阳性患者中卡介苗接种情况,认为卡介苗不能有效保护COVID-19的感染。由于目前缺少足够的临床研究结果,WHO表示不推荐卡介苗用于COVID-19的预防保护[48]。
三、相关机制研究
(一)异源性T细胞免疫
不相关的疫苗或病原体可能会存在相似的抗原表位,T细胞会识别结构相似的抗原表位,由此造成的异源性T细胞免疫是卡介苗对其他病原体的非特异性保护作用的机制之一[6, 50]。甚至有研究表明,氨基酸序列不同的表位生物化学特性可能很相似,T细胞可以识别氨基酸序列明显不同的T细胞表位[6]。这可以在一定程度上说明卡介苗的非特异性保护作用,但很难完全解释其有如此广泛的非特异性保护现象。
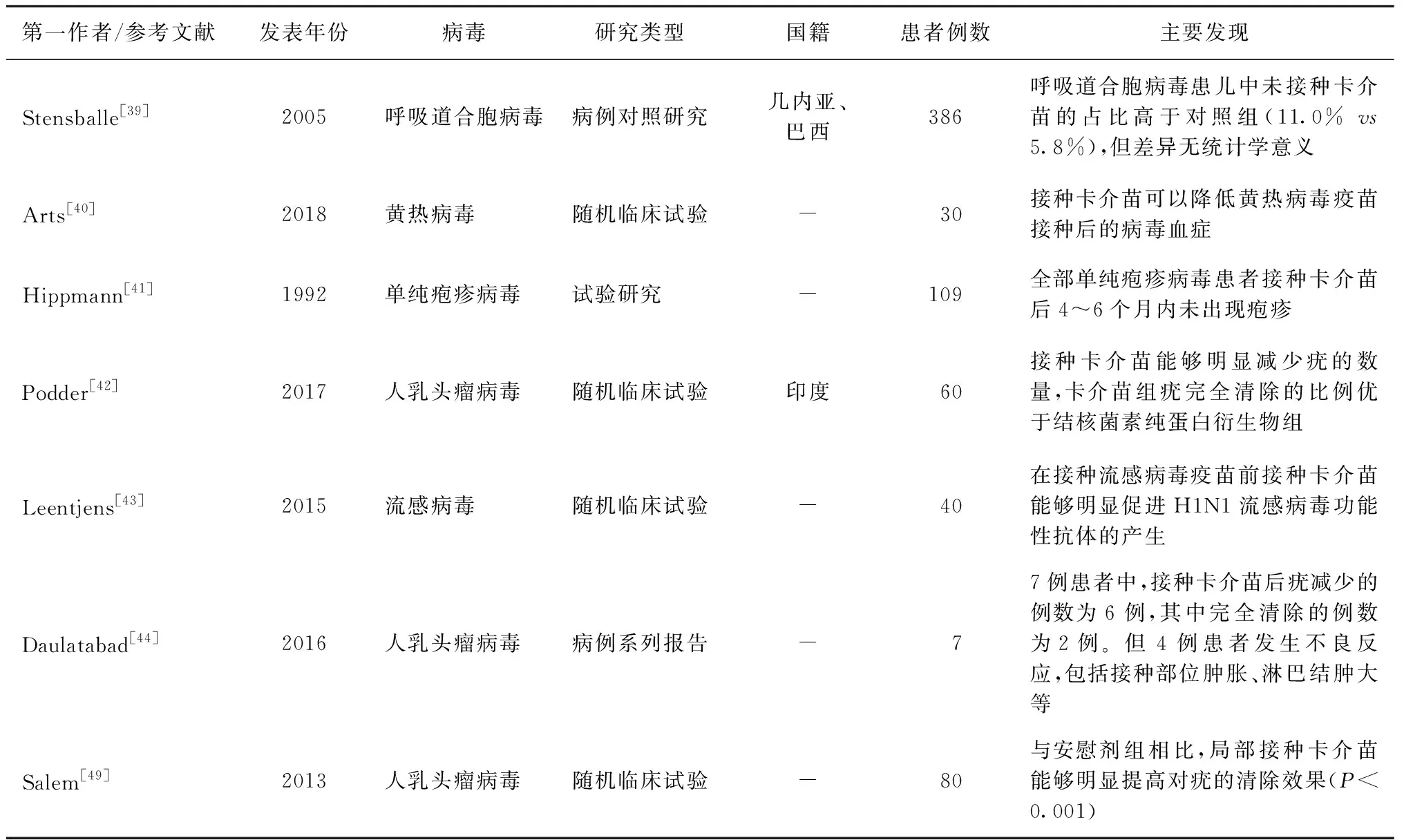
表2 卡介苗对病毒非特异性保护的相关研究
Kleinnijenhuis等[51]认为,卡介苗长期的非特异性保护作用主要是由于长期的辅助性T细胞1(Th1细胞)和Th17细胞效应作用,在卡介苗接种前和卡介苗接种1年后分别采集外周血淋巴细胞,分别用结核分枝杆菌、白色假丝酵母和金黄色葡萄球菌进行刺激,发现Th1细胞因子和Th17细胞因子有明显增加,说明长期的卡介苗非特异性保护作用在一定程度上依赖于异源性Th1/Th17反应。Ugolini等[52]发现卡介苗可以激活Toll样受体8(TLR8),产生白细胞介素(IL)-12 p40,从而影响下游滤泡辅助性T细胞(Tfh)反应,但是热灭活的卡介苗不能达到同样的效果。该研究为了证实TLR8在卡介苗的非特异性保护作用中是否发挥作用,在人群中进行了TLR8基因多态性的流行病学调查,发现rs3764880位点G等位基因(将启动子ATG变为GTG)在健康对照中的比例明显高于肺结核患者,且这种差异仅存于卡介苗接种的人群中,这说明该等位基因G能够提升卡介苗接种带来的保护作用,同时也表明在卡介苗接种后TLR8在促进Tfh细胞反应过程中发挥了重要作用。另外,也有研究者对接种卡介苗和未接种卡介苗的儿童进行研究,比较两组儿童接种其他疫苗后的抗体反应,发现接种卡介苗组再接种其他疫苗,其抗体浓度明显高于未接种卡介苗直接接种其他疫苗组,说明卡介苗可以促进其他疫苗接种后的体液免疫反应[53]。
(二)受训免疫
一般认为只有适应性免疫反应可以建立免疫记忆,但是越来越多的研究表明,在初次感染或接种疫苗后,固有免疫细胞在病原体再次入侵时可以迅速产生炎症因子,促进清除感染,这被称为“受训免疫”[54]。
为了证实卡介苗引起的受训免疫是否依赖于适应性免疫系统,有研究用缺乏B细胞和T细胞的严重联合免疫缺陷(SCID)小鼠进行试验,在接种卡介苗2周后用白色假丝酵母进行攻击,结果发现卡介苗接种的SCID小鼠的存活率明显高于生理盐水对照组,这说明卡介苗引起的非特异性保护作用并不依赖于B细胞和T细胞[55]。Arts等[40]进行的一项关于卡介苗对黄热病毒疫苗影响的研究发现,接种卡介苗可以激活单核细胞并产生IL-1β,从而降低黄热病毒疫苗接种后的病毒血症,该结果也支持了卡介苗可以引发固有免疫系统的受训免疫。
一些研究认为,卡介苗接种后单核细胞表型发生了变化。Bekkering等[56]在细胞水平建立了研究受训免疫的方法,在该研究中首先用卡介苗刺激单核细胞进行“训练”,再用脂多糖等刺激后检测细胞因子水平,发现IL-10和IL-1Ra均有一定程度的升高。Loenhout等[55]在接种卡介苗后的2周和3个月分别收集血液中的单核细胞,发现用NTM或其他不相关的病原体(金黄色葡糖球菌和白色假丝酵母)进行刺激,IL-1β和肿瘤坏死因子-α(TNF-α)的水平均会明显升高,且两个时间点采集的单核细胞中CD14+单核细胞均增加,且表达TLR4的CD14+单核细胞的比例明显升高,说明接种卡介苗会引起单核细胞的表型改变,同时伴有强烈的促炎反应。但是这种改变在接种后1年开始下降[51]。近期也有研究发现,卡介苗在骨髓细胞生成的早期发挥作用,并影响造血干细胞分化成单核细胞和巨噬细胞的表型,通过调节造血干细胞进行受训免疫也可能是卡介苗非特异性保护的原因[57-58]。
另外,表观遗传修饰和代谢驱动也是受训免疫形成的原因。Loenhout等[55]发现在接种卡介苗后3个月,与接种前相比,单核细胞中H3组蛋白第4位赖氨酸(histone H3 at lysine 4,H3K4)的三甲基化明显升高,从而提高IL-1β和TNF-α基因的转录水平,这也是接种卡介苗后受训免疫形成的原因之一。另外一项研究也发现,上述H3K4表观遗传修饰是由代谢改变造成的,在接种卡介苗后,单核细胞中糖酵解明显上调,谷氨酰胺代谢轻微上调,这些代谢改变影响了组蛋白H3K4的三甲基化和H3组蛋白第9位赖氨酸(histone H3 at lysine 9,H3K9)的三甲基化[59]。
综上所述,卡介苗的非特异性保护作用非常广泛,不仅可以减少其他NTM和病毒的感染,也可以优化其他疫苗接种的免疫反应[45-46]。卡介苗广泛的非特异性保护作用也提示,任何国家或地区在决定终止大规模接种卡介苗,或用新的疫苗取代卡介苗时,应将卡介苗的非特异性保护作用考虑在内。目前,关于卡介苗的非特异性保护机制尚不明确,受训免疫和异源性T细胞免疫可能分别是卡介苗的短期和长期非特异性保护作用的原因之一[51]。由于卡介苗对其他一些不相关的病原体产生非特异性保护的同时也会存在一些不良反应[44],因此在利用其非特异性保护作用的过程中应在其发挥的作用和不良反应之间做出平衡。自新中国成立以来,卡介苗在我国广泛应用,有效降低了结核病的发病率。但其对中国其他细菌和病毒感染的间接预防保护效果有待进一步探索。
