低温胁迫对黄秋葵幼苗相关生理特性的影响
2020-10-12陈瑶瑶严良文刘智成张双照郑作芸黄发茂刘晓佩
陈瑶瑶,严良文,刘智成,张双照,郑作芸,黄发茂,刘晓佩
(福建省龙岩市农业科学研究所,福建 龙岩 364000)
黄秋葵(Abelmoschusesculentus)原产于非洲,因其风味独特,营养丰富,保健效果佳,成为大众餐桌上的佳品[1-3]。随着生活水平的不断提高,人们对蔬菜新、奇、特的要求也越来越强,黄秋葵给种植户带来了良好的经济效益,是近年来南方地区早春蔬菜栽培的优势品种之一。黄秋葵喜温暖,对低温的反应很敏感[4],在早春低温下其植株生长受到抑制,影响了产量与品质,降低了农民的收益[5]。在低温胁迫下,植物自身会产生对逆境的适应性反应,据报道,低温胁迫后植物体内的SOD、POD、APX、CAT活性,MDA含量及可溶性蛋白含量会发生变化[6-8]。低温导致植物体内可溶性蛋白含量及膜脂过氧化物的变化已在茄子[9-10]、苦瓜[11-12]、黄瓜[13-14]等蔬菜上得到证实[15-16]。但关于黄秋葵幼苗在低温伤害后相关生理生化变化的系统研究报道极少。本试验以黄秋葵耐寒性较好的南洋品种为试材,结合福建闽西地区自然条件下的低温,研究了不同低温胁迫和持续时间条件下黄秋葵叶片膜脂过氧化产物 MDA含量、可溶性蛋白质含量、保护性酶活性的变化情况,旨在为后期探讨减轻低温危害途径和方法提供技术支持,为探索黄秋葵抗寒生理机制提供依据。
1 材料与方法
1.1 试验材料及处理方法
供试黄秋葵品种为南洋,半耐霜冻,耐冷性较好。试验在龙岩市农科所设施大棚内进行,种子经浸种催芽后播种于穴盘中,每钵播1粒种子,常规管理。当幼苗具有3~4片真叶时,挑选生长一致、健壮的植株各40株,转移至光照培养箱中,于 25 ℃/20 ℃(昼/夜)下生长2 d后将昼/夜温度调为:CK:25 ℃/20 ℃;T1:20 ℃/15 ℃;T2:15 ℃/10 ℃;T3:10 ℃/5 ℃,在光周期12 h/d下进行低温处理,分别于低温处理的第1、3、5、7 天选取植株第3~4片展开叶,混合后,测定各项生理生化指标,每处理3次重复。
1.2 测定项目及方法
超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性、抗坏血酸过氧化物酶(APX)活性、丙二醛(MDA)含量和可溶性蛋白质含量的测定按苏州科铭生物技术有限公司提供的检测试剂盒说明书介绍的方法进行。
1.3 统计分析
用SPSS 17.0软件进行数据分析。
2 结果与分析
2.1 低温胁迫对幼苗叶片可溶性蛋白含量的影响
在低温胁迫的过程中,可溶性蛋白质含量均随着温度下降而上升,但随着胁迫时间延长其含量有下降的趋势;当胁迫时间达到第5天时T2、T3处理间的可溶性蛋白质含量差异不显著(图1)。T3处理的可溶性蛋白质含量在各个阶段显著高于其它处理;T1和T2的可溶性蛋白质含量在胁迫第3天,T3在胁迫第5天时达到最大值,且均显著高于对照,分别比对照增加了77.32%、188.66%和287.45%。因此,在低温胁迫过程中,低温导致叶片中的可溶性蛋白质含量增加。

图1 低温胁迫下黄瓜秋葵叶片可溶性蛋白质含量的变化
2.2 低温胁迫对幼苗叶片MDA含量的影响
由图2可知,在低温胁迫过程中,MDA含量均呈上升的趋势,常温培养的黄秋葵幼苗叶片组织中MDA含量无明显变化;低温胁迫后,3个处理的MDA含量前期增速缓慢,至胁迫第3天时,T2和T3处理间均无显著差异,至第7天时 T1、T2和T3均达到最大值,分别比对照提高了55.86倍、107.73倍和129.82倍且各处理间差异极显著。

图2 低温胁迫下黄秋葵叶片MDA含量的变化
2.3 低温胁迫对幼苗叶片SOD活性的影响
在低温胁迫过程中,SOD活性随着温度降低和胁迫时间延长均表现为先升高后降低的变化趋势。低温胁迫处理第3天时各处理达最大值,分别为对照的84.97%、66.27%和39.09%;在处理第7天时SOD活性均降到最低值(图3)。
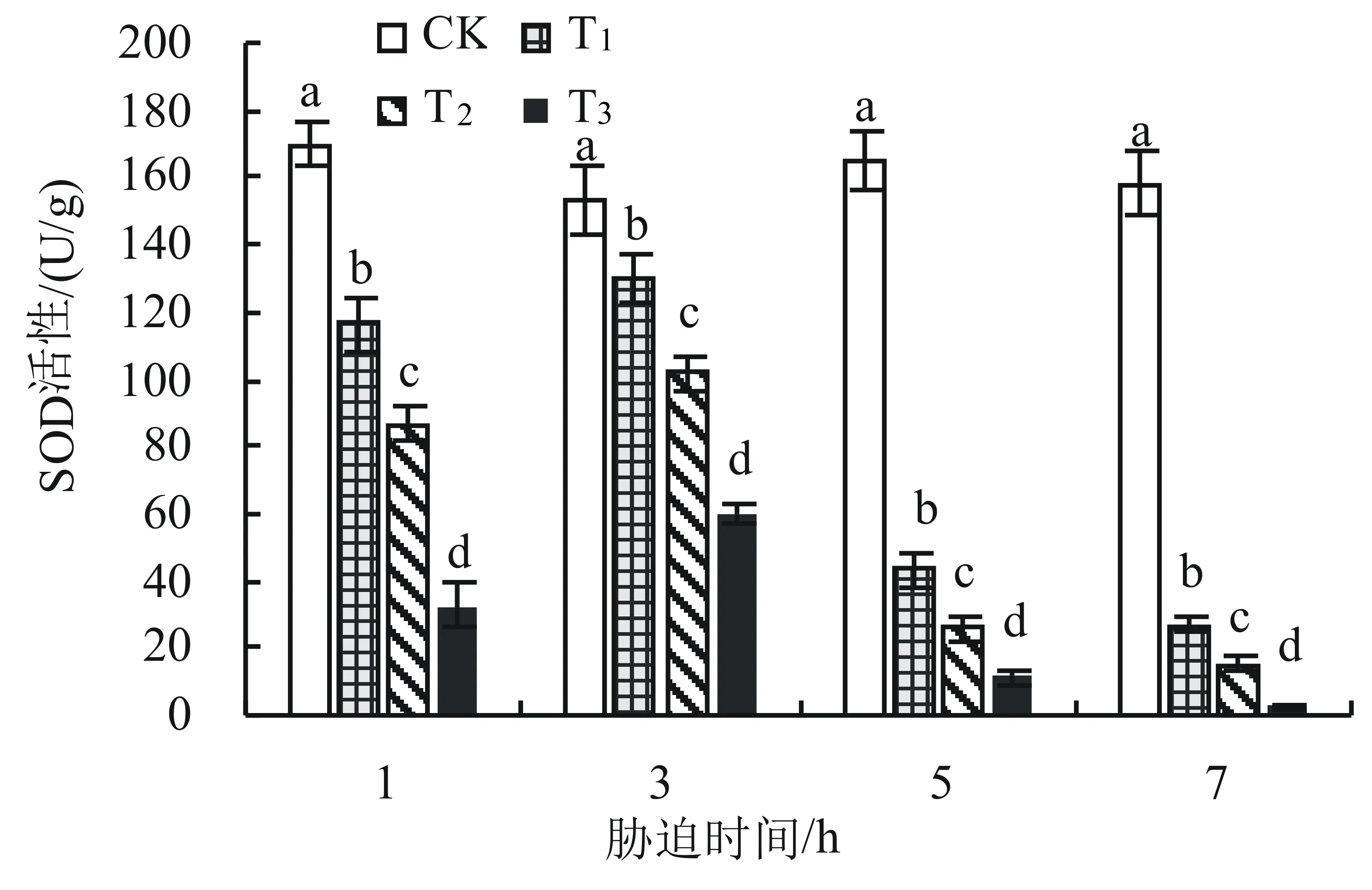
图3 低温胁迫下黄秋葵叶片SOD活性的变化
2.4 低温胁迫对幼苗叶片POD活性的影响
由图4可知,在低温胁迫开始后,各处理POD活性的变化趋势一致,在处理至第3天时急剧上升,而后平稳上升,但与对照组相比POD活性仍呈现显著差异。T1、T2和T3在胁迫7 d时的POD活性分别比同期对照增加464.5%、479.6%和866.3%,且T1处理与T2处理间无显著差异。
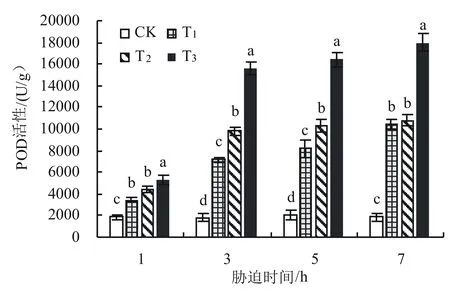
图4 低温胁迫下黄秋葵叶片POD活性的变化
2.5 低温胁迫对幼苗叶片CAT活性的影响
低温胁迫下,CAT活性变化与SOD相似,随着温度降低和胁迫时间延长呈先升高后降低趋势。与CK相比,胁迫第7天时,T1、T2和T3处理的CAT活性均达到最低值,分别为同期对照的16.02%、7.79%和3.03%。从胁迫第3天开始T2与T3的CAT活性无显著差异(图5)。

图5 低温胁迫下黄秋葵叶片CAT活性的变化
2.6 低温胁迫对幼苗叶片APX活性的影响
由图6可知,经低温胁迫后,APX的活性均呈先上升后下降的趋势,且温度越低APX活性越大。在胁迫处理第1天时T3活性达到最高;T1、T2处理以胁迫第3天时活性最高,分别比同期对照提高了6.0倍、6.63倍和10.6倍,且显著高于对照。

图6 低温胁迫下黄秋葵叶片APX活性的变化
3 讨论与结论
渗透调节是植物抵御低温逆境的一种重要生理机制,而可溶性蛋白则是植物体内重要的渗透调节物质之一[17-19]。本试验中,黄秋葵幼苗叶片在低温胁迫前期的可溶性蛋白含量迅速增加,但是随着胁迫时间的延长,其含量反而下降,这一试验结果与前人研究的结论相似[20-21],说明在一定的胁迫范围内,低温可以诱导黄秋葵叶片产生更多的可溶性蛋白,可溶性蛋白可以束缚细胞的水分从而减少失水,降低低温胁迫给植株带来的伤害。
膜脂过氧化的最终产物丙二醛(MDA)会对生物膜系统产生严重的损伤[22-23]。本试验发现随着低温胁迫程度加大和处理时间的延长黄秋葵幼苗叶片中MDA的含量呈显著增加的趋势,表明低温胁迫加剧了叶片细胞膜的过氧化程度。各处理的MDA含量在胁迫过程前期增速较缓,且无显著差异,说明前期受到低温伤害的程度较小;而后期MDA含量急剧升高,则有可能是低温已经严重伤害黄秋葵幼苗。
SOD、POD、CAT和 APX作为活性氧的酶促脱毒系统能清除植物体的活性氧自由基,在正常情况下,植物体内抗氧化酶系统中所产生的活性氧不足以使植物受到伤害,因为植物体内有一套行之有效的清除系统,但是这种低温逆境可以彻底打破这种高度平衡的过氧化状态,导致植株受到严重的损伤[24-25]。超氧化物歧化酶SOD是植物细胞抗氧化防御系统的第一道防线[26]。在本试验中,黄秋葵幼苗在低温胁迫期间SOD活性变化呈现先上升后降低的趋势,该研究结果与其他植物的研究结果一致[27-28]。SOD活性前期升高这可能是植物应对低温逆境时的自我保护,增强植物对低温的耐受力;但是随着低温时间的延长及低温程度加大,植物体内则无法及时清除毒害物质,这就会对植株产生严重的毒害作用。逆境下CAT与SOD活性变化相似,但是CAT活性的变化幅度不同于SOD活性,低温胁迫过程中CAT活性增幅高于SOD 活性增幅,这可能是这两种酶对低温条件有着不同的适应机制,这有待于进一步研究。过氧化物酶POD普遍存在于植物体内,它与呼吸作用、光合作用及生长素的氧化等都有着密切的关系。因此,POD 的活性能很好地反映出植物的生长状况。在低温胁迫下,前期黄秋葵的POD合成急剧增加,可能是植物抗逆自我保护,以降低活性自由基的积累,保护植物细胞免受氧化伤害。APX是植物体内清除H2O2的关键酶[5,29-30]。该试验结果表明低温胁迫下黄秋葵幼苗叶片APX活性呈现先升后降的趋势,APX活性的升高说明黄秋葵抵御膜脂过氧化的能力提高;而后期APX活性下降则可能是低温已经严重伤害黄秋葵幼苗。
综上所述,适当低温可以提高抗氧化保护酶系统相关酶活性,促进叶片中MDA和可溶性蛋白含量增加。低温胁迫下幼苗叶片中的可溶性蛋白、丙二醛(MDA)含量、过氧化物酶POD、超氧化物歧化酶SOD、过氧化氢酶CAT及抗坏血酸氧化酶APX的活性与对照差异显著,这些生理指标与黄秋葵的抵御低温能力密切相关,因此可以作为耐低温性鉴定的指标。本试验也证明,低温胁迫3天内细胞受到低温伤害的程度较小;随着低温时间延长低温会严重伤害黄秋葵幼苗。这些结果可为后期探讨人为介导缓解低温危害提供参考。
