光照度对绿球藻生长和营养成分的影响
2020-09-27王伟隆黄旭雄
纪 鹏,王伟隆,戴 颖,蒋 刚,黄旭雄,2,3
(1.上海海洋大学,农业农村部鱼类营养与环境生态研究中心,上海201306;2.上海海洋大学,农业农村部淡水水产种质资源重点实验室,上海201306;3.上海海洋大学,水产科学国家级实验教学示范中心,上海201306)
微藻种类繁多、分布广泛,是水域生态系统中重要的初级生产者。与传统农作物相比,微藻具有光合效率和表面积生产力高,营养需求简单,占用土地面积少等优势。微藻在固定CO2、净化废水、调节水质方面发挥着重要作用。微藻中含有丰富的色素、脂质、碳水化合物和脂肪酸等营养成分,可作为各种增值产品的原料,如生物柴油、颜料、生物医药和营养品等[1-2]。
微藻的生长率和生物量生产力通常取决于它们指数期内将生物量翻倍的能力[3]。藻类的生长受多种环境因子的影响[4-5],如温度、盐度、光照、pH等,其中光照是促进微藻细胞生长及其代谢活性的首要因素[6-8],是微藻将CO2转化为有机物质的必要能源,不仅影响微藻的光合速率和产量,而且影响藻细胞的生物化学组分,许多研究表明光照对色素、脂质和碳水化合物等细胞组分含量有所影响[9-10]。在藻类的长期进化中,不同种类的微藻对光照的适应存在差异,因此,适宜的光照度对微藻的生长和目标产物的积累都至关重要。
绿球藻(Chlorococcumsp.)能够利用有机碳源进行兼养培养,具有耐高温[11]和高氨氮[12]的特性,在养殖废水净化处理和水产动物养殖方面具有很大的开发价值和前景,关于绿球藻生长所需的温度、碳源和营养方式等影响因子已经做了研究,但其对于光照度的需求还未有过探究。本研究分别在不同的光照度下对绿球藻进行兼养培养,测定并分析了光照度对绿球藻生长、色素、脂质、碳水化合物和脂肪酸的影响,为绿球藻的大规模培养和产业化应用提供相应的理论依据。
1 材料与方法
1.1 试验设计
实验用绿球藻来自上海海洋大学生物饵料培养实验室。取处于对数生长期的绿球藻藻液,经3 000 r/min离心收集藻细胞,然后接种到1 L的灭菌三角烧瓶中,初始密度为5×106cells/mL,实验共设计6个光照度,分别为0、1 500、3 000、4 500、6 000、7 500 lux,每个梯度3个平行,共18个样品,实验所用培养基为改良f/2培养基,主要成分为(mg/L):C6H12O6500,NaNO3150,NaH2PO48.8,ZnSO4·4H2O·46,CuSO4·5H2O 20,MnCl2·4H2O 356,FeC6H5O7·5H2O 7800,NaMoO4·2H2O 14.6,Na2EDTA 8700,CoCl2·6H2O 24。实验在光照培养箱中进行,培养温度(35±1)℃,光周期12L ∶12D,每天定时摇藻4次,培养7 d。
1.3 测定指标和方法
1.3.1 绿球藻细胞密度、特定生长率和生物量的测定
实验期间,每天采用XB-K-25血球计数板测定绿球藻细胞密度,每个样品测定3次,并计算绿球藻的特定生长率K。实验结束后,将100 mL藻液通过微孔滤膜(孔径0.45 μm)抽滤,之后将滤膜置于85 ℃烘箱中烘干至恒重,并计算生物量(B)。
K=(lnNt-lnN0)/t
(1)
B(g/L)=(W1-W0)/V
(2)
式中,N0:初始藻细胞密度(cells/mL);Nt:经过时间t后藻细胞密度(cells/mL);t:培养时间(d);W0:微孔滤膜的质量(g);W1:含有藻液的微孔滤膜的质量(g);V:测定生物量所需藻液体积(L)。
1.3.2 绿球藻营养成分含量的测定
实验结束后,取一定体积的藻液,离心后用去离子水清洗三次,获取藻泥,并用甲醇提取绿球藻色素,采用分光光度计法[13-14]测定色素含量,根据以下公式计算叶绿素a(Pa)、叶绿素b(Pb)、类胡萝卜素(Pc)含量,最后,根据藻细胞密度和生物量换算成百分含量。
Pa(μg/106cells)=(-8.096 2×A652+16.516 9×A665)×V1/V2/细胞密度×106
(3)
Pb(μg/106cells)=(27.440 5×A652-12.168 8×A665)×V1/V2/细胞密度×106
(4)
Pc(μg/106cells)=4×A480×V1/V2/细胞密度×106
(5)
式中,A480、A652、A665分别是在波长为480、652、665 nm下测定的吸光度;V1:甲醇体积(mL);V2:测定藻液体积(mL)。
之后,将剩余藻液离心,并用去离子水反复清洗3次,将获得的藻泥置于-40 ℃下冷冻干燥,用于测定碳水化合物和总脂含量,碳水化合物含量测定采用苯酚-硫酸法[15],总脂含量采用氯仿甲醇法[16],之后采用BF3甲酯化法[17]测定脂肪酸含量,使用Agilent-7890A型气相色谱仪联合Agilent-5975C质谱仪分析脂肪酸,根据脂肪酸标准品的分析图谱、保留时间以及质谱的分析对脂肪酸进行定性分析,并按照峰面积归一化法计算各脂肪酸的相对含量。
1.4 数据处理与分析
所有数据以平均值±标准差(Mean±SD)表示,通过单因素方差分析和Duncan’s多重比较来评估各组实验数据之间的差异,P<0.05表示数据之间有显著性差异。所有数据都使用SPSS 17.0进行分析和处理。
2 结果与分析
2.1 不同光照度对绿球藻生长的影响
由图1可知,不同光照度对绿球藻生长影响不同,绿球藻在无光条件下(0 lux)没有生长,第3天藻细胞密度出现降低的趋势,显著低于其它各组;在光照条件下,1 500和3 000 lux组绿球藻经过1天的生长延缓期后开始进入指数增长期,之后进入稳定期,分别在第6天和第4天达到最大值24.69×106和22.93 ×106cells/mL,明显高于其它各组;随着光照度的进一步增加,4 500、6 000和7 500 lux组延缓期延长,绿球藻细胞密度降低。因此该绿球藻在无光条件下无法生长,在低光照条件下生长较好,高光照条件下生长受到抑制。

图1 不同光照度对绿球藻细胞密度的影响
由表1可知,绿球藻在光照度为0 lux时,特定生长率最小,为0.03,显著低于其它各组,随着光照度的增加,特定生长率先增大后减小,在光照度为1 500 lux达到最大值0.24,显著高于6 000和7 500 lux组。随着光照度的增加,绿球藻生物量先增大后减小,0 lux组生物量显著低于其余各组,当光照度为1 500和3 000 lux时,绿球藻具有较高的生物量,分别为0.47、0.44 g/L,显著高于其它各组。由此可知,绿球藻最佳生长所需光照度为1 500~3 000 lux。

表1 不同光照强度对绿球藻特定生长率、生物量和类胡萝卜素/叶绿素的影响
2.2 不同光照强度对绿球藻营养成分含量的影响
绿球藻细胞色素含量受光照度的影响,由图2可知,随着光照度的增加,叶绿素a、叶绿素b和类胡萝卜素均表现为先增加后降低的趋势,光照度为3 000 lux时色素含量较高,叶绿素a、叶绿素b和类胡萝卜素含量分别为0.41%、0.06%、0.10%,与1 500 lux组无显著差异,但显著高于其它各组。在光照条件下,随着光照强度的增加,绿球藻类胡萝卜素/叶绿素比值增大(见表1),7 500 lux组达到最大值0.23,显著高于1 500、3 000和4 500 lux组。

图2 不同光照强度对绿球藻色素含量的影响
由图3可知,随着光照度的增加,总脂含量在0 lux组有最大值17.53%,显著高于其它各组,随着光照度从1 500增加到7 500 lux,总脂含量无显著变化;碳水化合物含量在0 lux组显著低于其它各组,其余各组之间无显著差异。

图3 不同光照强度对绿球藻总脂和碳水化合物的影响
光照度影响绿球藻的脂肪酸类型和含量,对绿球藻脂肪酸组成测定结果见表2,绿球藻中主要含有C16和C18脂肪酸,其中C16 ∶0、C18 ∶2n6(亚油酸LA)、C18 ∶3n3(亚麻酸ALA)含量较高。随着光照度的增加,C14 ∶0、C16 ∶0、C18 ∶0、C16 ∶1、C17 ∶1含量降低,从而导致饱和脂肪酸(SFA)和单不饱和脂肪酸(MUFA)含量减小;0~3 000 lux,多不饱和脂肪酸(PUFA)含量随着光照度的增大逐渐提高,当光照度大于3 000 lux后不再有显著性变化;随着光照度的增加,LA含量先增大后减小,在光照度为1 500 lux时达到最大值25.46%,ALA含量随着光照度的增加而增加,当光照度大于4 500 lux时不再变化,维持在43%~44%左右。
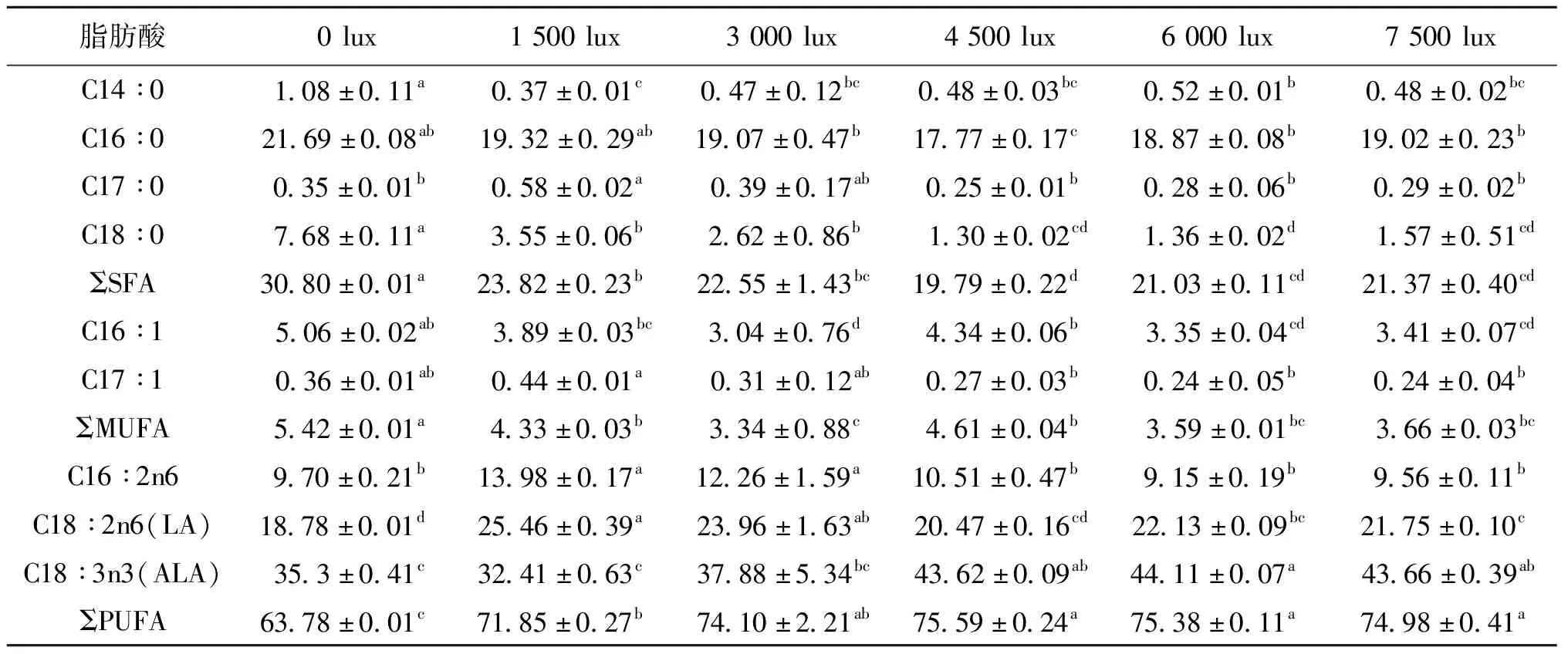
表2 不同光照强度对绿球藻脂肪酸组成的影响(%总脂肪酸)
3 讨论
本研究结果表明该绿球藻在无光照条件下无法生长,特定增长率几乎为0,说明该绿球藻不能进行异养生长。当光照度为1 500 lux时,绿球藻细胞密度、特定生长率和生物量提高并达到最大值,随着光照度增加到3 000 lux,绿球藻不再进一步生长,当光照度从4 500 lux增加到7 500 lux,绿球藻生长下降,表明低光照促进绿球藻生长,高光抑制生长,这与大多数微藻具有相同的特性,如斜生栅藻(ScenedesmusobliquusCNW-N)[18]、微绿球藻(Nannochloropsissp.)[7]、凯氏拟小球藻(Chlorellakessleri)[19]等。在无光或低光照度下,微藻生长往往处于光限制状态,光照度的增加会促进微藻的生长,直到达到饱和光照度。然而,当光照度超过饱和光度时,通常会导致Fv/Fm比值急剧下降,对光合系统Ⅱ(PS Ⅱ)造成光氧化损伤,即发生光抑制现象[20-21],降低光合效率,从而降低生物量的生产,这与Khoeyi等[22]、Chen等[23]的研究结果相一致。根据实验结果可以推测绿球藻达到光饱和点所需光照强度为3 000 lux左右,绿球藻培养所需适宜的光照强度为1 500~3 000 lux。
色素是微藻捕光和传递光量子的重要成分,与光合作用效率密切相关。绿藻中主要存在叶绿素和类胡萝卜素两类色素。本研究结果表明,当光照度从0增加到3 000 lux,绿球藻叶绿素a、叶绿素b和类胡萝卜素含量均不断提高,这是由于在光限制条件下,藻类可以通过增加光合单位的数量及光捕获色素蛋白复合体的大小来增加细胞的色素[24-25],提高采光能力,使光合速率维持在较高水平。当光照度超过3 000 lux,绿球藻各色素含量开始逐渐下降,其原因主要是在高光照度下,细胞不需要较高的色素来捕获光量子,并通过降低色素含量减少对光的吸收,从而避免光氧化损伤[26-27]。类胡萝卜素除了具有捕光功能外,在保护藻细胞免受光抑制中发挥着主要作用,本研究结果表明,在光照条件下,随着光照度的增加,类胡萝卜素/叶绿素之比逐渐增大,这与对杜氏盐藻(Dunaliellasalina)[28]的研究结果类似。当受到过度光照胁迫时,藻类往往能够增加类胡萝卜素的组成比例来应对自身的氧化损伤,参与光保护过程。因此,在持续高光度下,叶绿素比类胡萝卜素下降更快,所以导致类胡萝卜素/叶绿素比值增大。
脂质和碳水化合物是微藻主要的储能形式[29],研究表明微藻将获得的能量用于脂类和碳水化合物的生物合成过程存在竞争关系,与脂质相比,碳水化合物的合成所需的ATP和NAD(P)H更少,微藻会将碳和能量倾向于碳水化合物的合成[30]。本研究结果表明,在无光照条件下脂质和碳水化合物含量相近,绿球藻无法利用光照和葡萄糖获取能量,不存在这种竞争关系,但在光照条件下,绿球藻总脂含量降低,碳水化合物含量升高,差异增大,说明绿球藻能够将从光照和葡萄糖中获得的能量优先转化为用于碳水化合物的合成。
C16和C18系列脂肪酸是藻类细胞合成的主要脂肪酸[31]。光照度对微藻脂肪酸组成和含量的影响因种而异,大多数研究表明较高的光照度能够促进SFA和MUFA的合成,降低PUFA的合成[31-33]。本研究结果表明,随着光照度的增加,绿球藻SFA和MUFA均出现了不同程度的下降趋势,而PUFA含量却显著提高,尤其是ALA,这与Krzeminska等[32]研究结果不同,但与Liu等[34]研究结果一致。光能够诱导基质中的pH、Mg2+和NADPH的变化,最终影响和调节脂肪酸合成过程中关键酶的活性[35]。PUFA是光合膜的重要组成成分,在维持光合膜的结构完整和功能方面发挥着重要作用,随着光照度的增加,微藻能够通过加速D1蛋白的合成来完成脂肪酸的去饱和,从而提高藻细胞对光的耐受能力[36-37]。另一方面,碳链的去饱和过程中需要分子氧的参与[38],随着光照度的增加,绿球藻光合效率提高,产生更多的分子氧,加速碳链的去饱和,从而促进PUFA的合成,当光照达到饱和点后,PUFA不再增加。因此,在低光照度下,有利于绿球藻SFA和MUFA的合成,在高光照度下,有利于PUFA的合成。
综上,绿球藻在光照度为3 000 lux生长时可获得较高的细胞密度和生物量,同时,绿球藻色素、碳水化合物和PUFA含量也维持在较高水平。
