甘露聚糖酶和纤维素酶对玉米-小麦-豆粕型日粮的体外消化评价
2020-09-27刘芦鹏王占彬马文锋
毛 培,刘芦鹏,王占彬,马文锋*
(1.兰州大学 草地农业科技学院,甘肃 兰州 730020;2.河南科技大学 动物科技学院,河南 洛阳 471023)
目前,以植物原料为基础的畜禽日粮中,存在较高含量的非淀粉多糖,如甘露聚糖和纤维素等[1]。这一类多糖经动物采食后可以与食糜中的水分子结合,增加胃肠内容物的黏度,抑制机体对营养物质的利用效率[1-2]。由于单胃动物不能分泌非淀粉多糖酶,因此在畜禽生产中主要通过外源酶的方式进行添加。研究表明,在日粮中添加一些非淀粉多糖酶,如甘露聚糖酶和纤维素酶等,可以破坏植物原料的细胞壁、释放其包裹的营养物质、提高日粮养分与胃肠消化酶的接触面积、专一性的将部分多糖分解为小分子物质,改善肠道消化环境,促进对日粮营养物质的消化吸收[3-5]。
在评价外源酶的作用效果时,可采用动物试验法和体外酶解方法[6-7]。在通过动物试验法进行评价时,可以直观地检测添加酶制剂对动物生长性能的影响。然而,动物试验法周期较长,也需保证相当数量的动物群体才能完成,因此难以评价多种酶制剂的作用效果[8]。近年来研究发现,体外酶解方法可以合理地模拟动物消化环境,同时具备试验周期短、重复性高等诸多优点,可以很好地反映添加酶制剂对日粮营养物质消化率的影响[8]。因此,本试验主要采用体外酶解法,分别以干物质和粗蛋白质酶解率、还原糖生成量为评价指标,研究了甘露聚糖酶和纤维素酶对玉米-小麦-豆粕型日粮营养物质酶解效率的影响。
1 材料与方法
1.1 酶制剂
试验所用甘露聚糖酶和纤维素酶均为单酶,购自河南仰韶生化工程有限公司。酶活性分别为甘露聚糖酶50 000 U/g、纤维素酶10 000 U/g。
1.2 试验设计及日粮组成
本试验采用单因子试验设计方法,每个酶分别设置5个浓度梯度,每个梯度分别设置6个重复,其中甘露聚糖酶的添加浓度分别为5、15、25、35、45 U/g,纤维素酶的添加浓度分别为1、3、5、7、9 U/g。
试验日粮组成及营养见表1。粉碎小麦、豆粕、玉米和麸皮各200 g,过60目筛网后,用四分法分别获得四种原料各50 g、20 g、20 g和10 g,混匀后置于包装袋内密封,备用。当取样时,采用十万分之一天平称取试验日粮样品。
1.3 体外酶解操作过程
准确称取1 g左右的饲料样品(精确至0.0001g),置于透析袋后编号,置于150 mL三角瓶中,加入20 mL HCl-NaCl缓冲液(0.1 mol/L,pH2.0)和0.1 g的胃蛋白酶(Sigma,P7000),用封口膜封紧瓶口。在厌氧条件下,将三角瓶置于39℃恒温摇床内振荡6 h,震荡频率为70 r/min。

表1 试验日粮组成和营养成分Table 1 Ingredient composition of the experimental diets
模拟小肠酶解过程:体外消化6 h后,暂停摇床,取出三角瓶打开封口膜,向三角瓶中准确加入20 mL磷酸缓冲液(0.1 mol/L,pH 7.0)和2.5 mL胰蛋白复合酶溶液,在溶液pH 7.0、厌氧环境下,封好三角瓶的瓶口,置于39℃恒温摇床中震荡24 h,震荡频率为70 r/min。
还原糖生成量测定:24 h后取出三角瓶中透析袋,室温静置20 min,吸取1 mL上清液于试管中,加入DNS检测液2 mL,于水浴锅中煮沸5 min,取出,冷却至室温,加入9 mL蒸馏水,混匀。移液枪移取混合液于比色皿中,在540 nm光波处测定各溶液的吸光度值,通过还原糖标准曲线,计算上清液中的还原糖含量。
还原糖生成量(mg/g)=(C×VT)/(m×VS) ×1000
式中:C:标准曲线所查糖量(mg);VT:提取液的体积(mL);m:样品的质量(g);VS:测定时的溶液体积(mL)。
干物质和粗蛋白酶解效率的测定:使用绝干称重的滤纸对三角瓶中的酶解残渣真空抽滤,将抽滤后的残渣和滤纸置于105 ℃烘箱中烘干至恒重,使用半自动定氮仪测定粗蛋白含量。干物质和粗蛋白酶解率的计算公式如下:
干物质酶解率(%)=(M0-M1)/M0×100
粗蛋白质酶解率(%)=(M0×CP0-M1×CP1)/(M0×CP0)×100
式中:M0:饲料样品干物质重量(g);M1:未酶解残渣干物质重量(g);CP0:饲料样品粗蛋白含量(%);CP1:未酶解残渣粗蛋白含量(%)。
1.4 数据分析
以重复为单位,采用SAS(2001)中GLM模型进行数据分析,通过线性(Linear)和二次(Quadratic)效应评估不同浓度甘露聚糖酶和纤维素酶对日粮还原糖生成量、干物质和粗蛋白酶解效率的影响,处理间显著性判定水平为P<0.05,数据平均值以LSMEANS表示。
2 结果与分析
2.1 添加甘露聚糖酶对日粮还原糖生成量、干物质和粗蛋白酶解效率的影响
由表2可知,试验日粮添加甘露聚糖酶时,可以显著提高试验日粮的还原糖生成量(线性和二次,P<0.05)。当甘露聚糖酶添加量分别为25、35 U/g时,还原糖生成量显著高于对照组和5 U/g剂量添加组(P<0.05)。
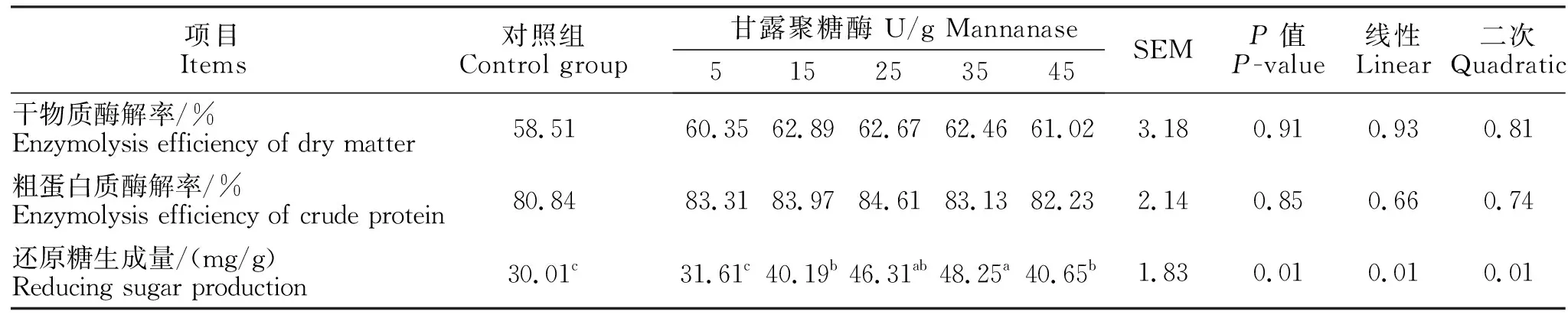
表2 添加甘露聚糖酶对日粮还原糖生成量、干物质和粗蛋白酶解率的影响Table 2 The effects of mannanase on the reducing sugar production, enzymolysis efficiency of dry matter and crude protein in the diet
由表3可知,随着甘露聚糖酶添加比例的提高,还原糖生成量与甘露聚糖酶含量存在线性和二次方程关系。本试验中关于甘露聚糖酶的最佳添加量由线性方程推导得出,以还原糖为评价指标:该方程为Y=45.07-0.860(20.69-x),推导得出甘露聚糖酶的最适添加量为20.7 U/g。

表3 甘露聚糖酶添加水平与还原糖生成量的关系Table 3 The correlation between the additional level of mannanase with reducing sugar production
2.2 添加纤维素酶对日粮还原糖生成量、干物质和粗蛋白酶解效率的影响
由表4可知,添加纤维素酶对干物质和粗蛋白酶解率无显著影响,但能显著提高日粮的还原糖生成量(线性和二次,P<0.05)。当纤维素酶的添加量为5、7、9 U/g时,还原糖生成量显著高于其余各组(P<0.05)。
由表5可知,随着日粮中纤维素酶添加比例的提高,还原糖生成量与纤维素酶含量存在线性和二次方程关系。本试验中关于纤维素酶的最佳添加量由线性方程推导得出,以还原糖为评价指标:该方程为Y=51.85-2.625(6.297-x),推导得出纤维素酶的最适添加量为6.3 U/g。
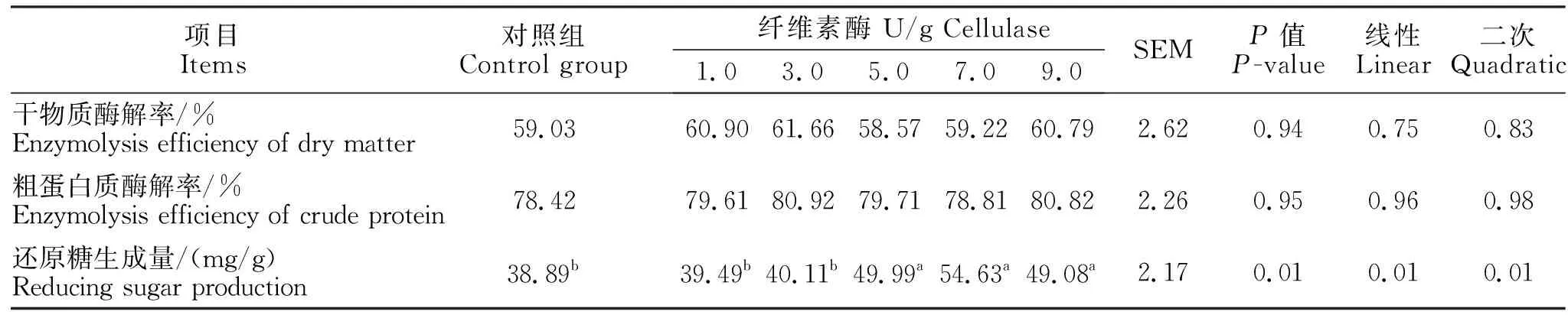
表4 添加纤维素酶对日粮还原糖生成量、干物质和粗蛋白酶解效率的影响Table 4 The effects of cellulase on the reducing sugar production, enzymolysis efficiency of dry matter and crude protein in the diet

表5 纤维素酶添加水平与还原糖生成量的关系Table 5 The correlation between the additional level of cellulase with reducing sugar production
3 讨 论
酶制剂是从动物、微生物或植物中提取的具有生物催化活性的一类蛋白质,分子量较高,具有高效性和专一性的特点,其在适宜的温度、pH等环境条件下具备较高活力,可按实际生产需要进行合理使用[9]。日粮的植物原料结构较为复杂,其所含的多糖类物质难以被动物消化吸收,因此在饲料中通常添加多种非淀粉多糖酶,才能更好的发挥酶制剂的水解作用[10]。然而,非淀粉多糖酶的种类较多,不同种类酶的来源、作用位点和专一性底物也存在差异[11]。
目前,甘露聚糖酶和纤维素酶均已应用于不同领域,如医药、纺织、食品和饲料行业等[12-14]。在豆粕型日粮中甘露聚糖含量较高,添加甘露聚糖酶可以专一性的提高甘露聚糖的利用效率,消除其对动物消化道带来的抗营养作用[14]。研究表明,在蛋鸡或仔猪日粮中添加适宜剂量的甘露聚糖酶,可以提高蛋鸡的产蛋量和平均蛋重[15],或可以提高仔猪的平均日增重,同时降低仔猪腹泻率[14]。纤维素酶的来源较为广泛,其中微生物源的纤维素酶生产成本较低,应用领域更为广泛[12-13]。在日粮中添加纤维素酶,可以在一定程度上刺激动物机体内源酶的分泌,打破细胞壁结构,使消化酶与饲料中营养物质充分接触,进而改善饲料的养分利用率,同时也可改变纤维多糖的物理结构,进而改变纤维源的系水力,降低肠道内食糜的黏度,降低肠道负担,从而提高动物生长性能[13]。
本试验中,当甘露聚糖酶和纤维素酶的添加浓度分别由5 U/g提高至45 U/g或由1 U/g提高至9 U/g时,可以显著提高日粮还原糖的生成量,然而对粗蛋白质和干物质酶解率无明显影响,相关原因尚不清楚,有待于进一步的研究。这也说明在评价甘露聚糖酶和纤维素酶的体外作用效果时,还原糖生成量指标更为敏感,与酶的添加剂量存在效应关系。也在一定程度上表明,在日粮中添加甘露聚糖酶和纤维素酶时,其发挥作用的主要机制是将大分子糖苷键分解为小分子糖,进而提高营养物质利用率[16-17]。
4 结 论
甘露聚糖酶和纤维素酶的添加浓度分别由5 U/g提高至45 U/g和由1 U/g提高至9 U/g时,显著提高了日粮的还原糖生成量(线性和二次,P<0.05)。以还原糖生成量为评价指标,甘露聚糖酶和纤维素酶添加的最适添加浓度分别为20.7 U/g和6.3 U/g。
