食源性单核细胞增生型李斯特菌lmoF2365_0032基因克隆及序列分析
2020-09-24刘扬扬马勋康立超李红欢钱凌霄江婉琳阮婷玉
刘扬扬 马勋 康立超 李红欢 钱凌霄 江婉琳 阮婷玉
摘要:对食源性单核细胞增生李斯特菌LM5567的lmoF2365_0032基因进行克隆和序列分析,利用PCR方法对lmoF2365_0032基因进行扩增,将扩增产物连接到pMD19-T载体上,筛选阳性菌株进行测序比对分析。结果显示,扩增获得的lmoF2365_0032基因序列长度为811 bp,克隆得到lmoF2365_0032基因的阳性转化子;生物信息学分析表明,lmoF 2365_0032蛋白属于跨膜蛋白,无信号肽序列;二级结构中无规则卷曲和延伸链结构占比较大,分别是37.86%和36.89%;序列比对结果表明,LM5567菌株lmoF2365_0032基因的核苷酸序列与02-6680菌株(4b,奶酪,加拿大)、10-0811菌株(1/2b,螃蟹,加拿大)的相似性均为99.7%;与10-0809菌株(4b,粪便,加拿大)、81-055菌株(4b,脑脊髓液,加拿大)、02-1103菌株、02-1792菌株(4b,奶酪,加拿大)、81-0861菌株(4b,卷心菜,加拿大)的相似性均為99.8%;LM5567 lmoF 2365_0032基因编码的氨基酸序列与上述菌株相似性均为94.7%。试验成功克隆了lmoF 2365_0032基因,可为进一步探究lmoF2365_0032基因功能奠定基础。
关键词:食源性单核细胞增多李斯特菌;lmoF2365_0032基因;PCR;生物信息学分析
中图分类号:S852.61
文献标志码:A
文章编号:1002-1302(2020)16-0074-05
单核细胞增生李斯特菌(Listeria monocy-togenes,LM)是重要的食源性人兽共患李斯特菌病的胞内寄生菌,与大肠杆菌、沙门氏菌、志贺氏菌并称为世界四大食源性致病菌[1-2]。LM可突破宿主的肠道屏障、血脑屏障和血胎屏障,临床上引起人和多种动物的脑炎、脑膜炎、败血症、流产及发热性胃肠炎等[3]。李斯特菌病具有较高的死亡率,免疫功能不全者、孕妇、新生儿及胎儿都是李斯特菌病的易感群体,该病在动物中主要见于牛、羊、骆驼等反刍动物[4-5]。因此,它对公共安全和畜牧业发展危害极大。
LM基因组中存在多个毒力因子基因簇,这些基因簇编码的蛋白在感染机体过程中发挥着重要作用。arc基因由4部分组成,分别是arcA、arcB、arcC和arcD,它们分别编码精氨酸脱亚胺酶系统(arginine deiminase system,ADS)中的精氨酸脱亚氨酶(ADI)、鸟氨酸转氨甲基酶(OTC)、氨甲酸激酶(CK)和精氨酸鸟氨酸逆向转运体[6]。arc基因编码的蛋白在LM调节酸平衡中发挥重要作用,2016年Maury等对104株不同来源的代表性LM菌株进行全基因组序列分析,挖掘出15个高毒力分离株具备而参考菌株不具备的假定毒力因子基因/基因簇,其中包括精氨酸鸟氨酸逆向转运体基因,但是与以往研究结果不同的是,编码精氨酸鸟氨酸逆向转运体的基因除了arcD外,还有3个未知功能的基因,其中就包括lmoF2365_0032基因[7]。为此,对笔者所在实验室保存的LM进行了检测,发现54株食源性单核细胞增生李斯特菌中只有19株可以检测到lmoF 2365_0032基因。arc基因编码的未知功能的蛋白是否与精氨酸代谢、抗酸应激以及毒力有关,目前尚不清楚。
本试验克隆分析编码精氨酸/鸟氨酸逆向转运体的lmoF2365_0032基因,并对其进行生物信息学分析,旨在为进一步探究食源性单核细胞增生李斯特菌精氨酸鸟氨酸逆向转运体基因的功能奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒 单核细胞增生李斯特菌LM5567菌株分离自新疆农垦科学院;大肠杆菌(Escherichia coli) DH5α,购自北京全式金生物技术有限公司。
1.1.2 主要试剂和仪器 PCR Mix、ddH2O、DL 2 000 DNA Marker,均购自广州东盛生物科技有限公司;脑心浸液培养基(BHI),购自青岛高科技园海博生物技术有限公司;LB培养基,购自青岛高科技园海博生物技术有限公司;细菌基因组DNA提取试剂盒,购自天根生化科技(北京)有限公司;PCR仪(Bio-Rad MyCycler thermal);电泳系统和凝胶成像系统(BIO-RAD-2000),购自美国Bio-Rad公司。
1.1.3 引物设计与合成 根据文献[4]中提供的Gene (Locus tag),运用Primer 5.0软件自行设计编码LM5567菌株精氨酸鸟氨酸逆向转运蛋白的lmoF2365_0032基因的特异性引物,引物序列为5′-TTTGGACCGACTTGATTG-3′(F)、5′-ACCCACAAATGCGATAGA-3′(R),预期扩增片段长度为811 bp,引物由北京六合华大基因科技有限公司合成。
1.2 方法
1.2.1 菌株培养和基因组DNA的提取 将LM5567菌株从-80 ℃保存条件下取出,接种于BHI平板上,37 ℃ 培养16~18 h,挑取单菌落接种于BHI液体培养基中,37 ℃、180 r/min培养16~18 h,采用 1.5 mL 试管收集菌体,参考细菌基因组DNA提取试剂盒说明书提取LM5567菌株的全基因组DNA,于-20 ℃条件下保存备用。
1.2.2 PCR检测 PCR扩增体系:模板DNA 2 μL,上下游引物各1 μL,2×PCR Mix 10 μL,加ddH2O至体积20 μL。PCR扩增程序:95 ℃预变性5 min;95 ℃变性40 s,50 ℃退火45 s,72 ℃延伸 1 min,72 ℃再延伸10 min,共30个循环;4 ℃保存60 min。
1.2.3 PCR扩增产物的检测 PCR扩增产物的电泳检测:琼脂糖凝胶加溴化乙锭(EB)染色,将 10 μL PCR扩增产物及DL 2 000 marker上样后进行电泳,电压为150 V,电流为 50 mA,25 min后在凝胶成像仪下观察电泳结果。
1.2.4 目的基因克隆及测序 PCR扩增产物经琼脂糖凝胶电泳检测后,纯化回收目的条带,将回收产物与pMD19-T载体过夜连接,采用冷热激法将连接产物转化至大肠杆菌DH5α感受态细胞中,涂布于含Ampr(100 μg/mL)的LB固体培养基平板上,37 ℃培养 12~16 h。筛选出阳性克隆菌,将鉴定的阳性克隆菌送北京六合华大基因科技有限公司测序。
1.2.5 生物信息学分析 利用ProtParam软件(http://web.expasy.org/ protparam/)分析基因序列的基本理化性质,利用SignalP 4.1 Server软件(http://www.cbs.dtu. dk/services/SignalP/)预测蛋白质的信号肽;利用TMHMM软件(http://www.cbs.dtu.dk/services/ TMHMM-2.0/)在线分析蛋白质跨膜结构;利用SOPMA软件(https://npsa-prabi.ibcp.fr/cgi- bin/npsa_automat.pl?page=npsa_sopma.html)和SWISS-MODEL软件(http://swissmodel.expasy.org/)分别预测蛋白质二级结构和三级结构。
2 结果与分析
2.1 lmoF2365_0032基因PCR扩增和克隆测序
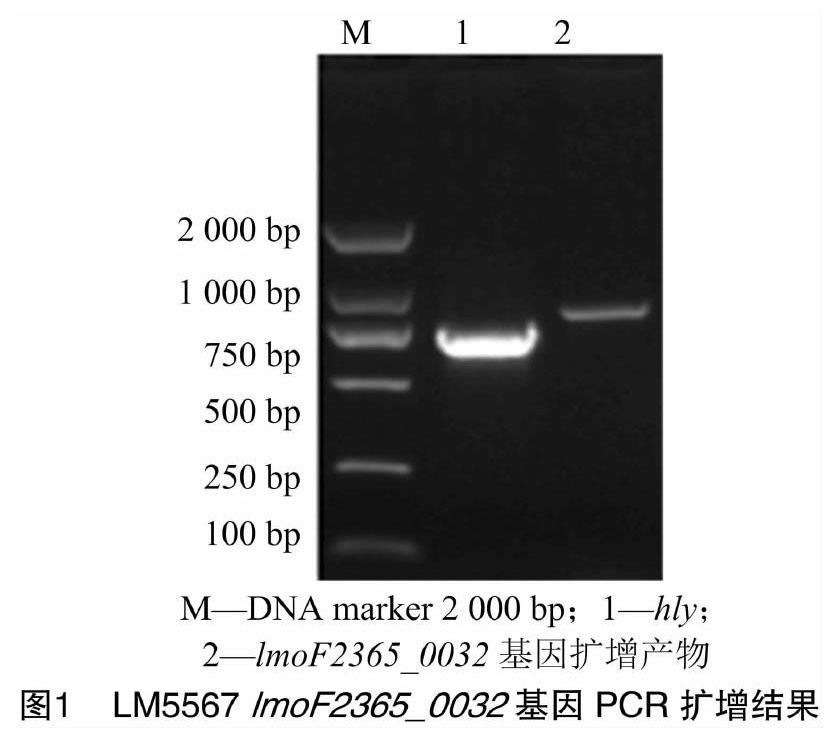
利用特异性引物扩增lmoF2365_0032基因,PCR产物经1%琼脂糖凝胶电泳检测,在811 bp处有单一清晰条带,与lmoF2365_0032基因扩增产物预期大小一致(图1)。经克隆转化后,将验证为阳性菌的质粒提取物送至北京六合华大基因科技有限公司测序,经测序进一步验证,成功获得序列长度为811 bp的lmoF2365_0032基因阳性转化子。
2.2 lmoF2365_0032编码蛋白的理化性质分析
利用ExPASY在线软件中编码ProtParam工具分析LM5567 lmoF2365_0032基因编码蛋白的理化性质,得出该蛋白理论等电点为8.51,分子量为 22 973.62,原子组成是C1 050H1 685N253O314S3,脂肪系数是101.75,总疏水性平均数是-0.124,负电荷氨基酸残基(Asp+Glu)22个,正电荷氨基酸残基(Arg+Lys)24个,不稳定系数是23.31(<40为稳定蛋白),属于稳定蛋白。
2.3 lmoF2365_0032编码蛋白信号肽和跨膜区分析
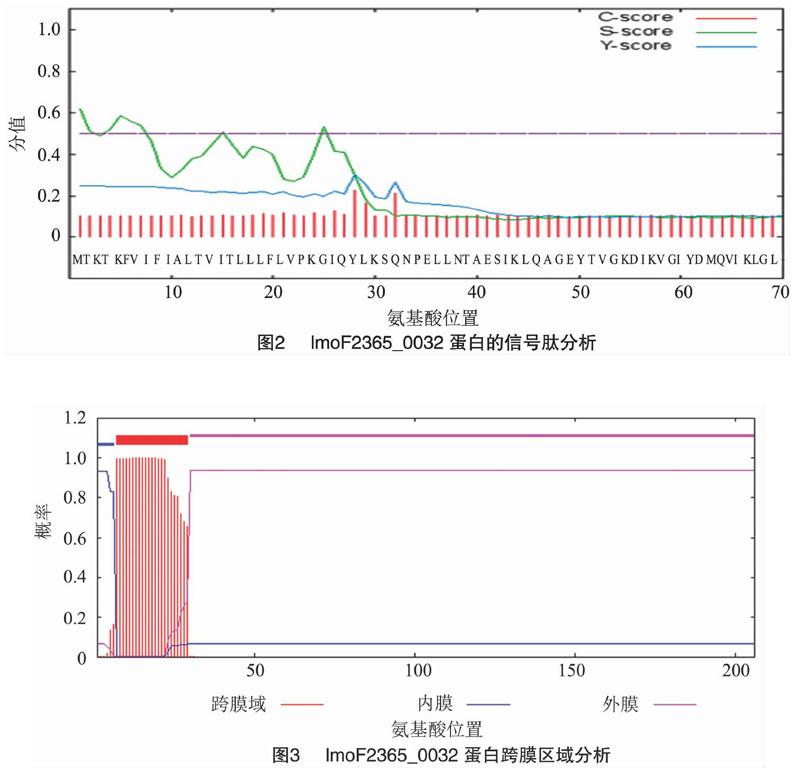
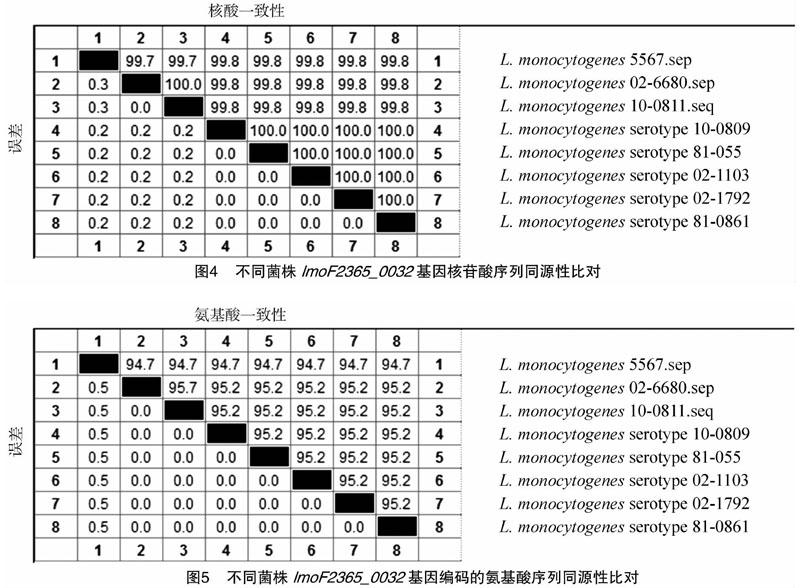
利用SignalP 4.1 Server软件(http://www.cbs.dtu. dk/services/SignalP/)预测lmoF2365_0032蛋白质的信号肽,结果(图2)表明,lmoF2365_0032蛋白无信号肽,属于非分泌蛋白。利用TMHMM软件(http://www.cbs.dtu.dk/services/ TMHMM-2.0/)分析蛋白质跨膜结构,结果(图3)显示,lmoF2365_0032为跨膜蛋白,跨膜区位于7~29氨基酸处。
2.4 lmoF2365_0032基因核苷酸序列同源性比对
将测序得到的结果进行序列拼接,得到LM5567菌株的lmoF2365_0032基因的核苷酸序列,将其与GenBank上公布的6株不同来源的lmoF2365_0032基因的核苷酸序列进行同源性比对,结果(图4)显示,LM5567菌株的lmoF2365_0032基因的核苷酸序列与02-6680菌株(4b,奶酪,加拿大)、10-0811菌株(1/2b,螃蟹,加拿大)相似性均为99.7%;与10-0809菌株(4b,粪便,加拿大)、81-055株(4b,脑脊髓液,加拿大)、02-1103菌株、02-1792菌株(4b,奶酪,加拿大)、81-0861菌株(4b,卷心菜,加拿大)的相似性为99.8%。
2.5 lmoF2365_0032基因序列编码氨基酸同源性比对
利用DNSstar软件推导出的LM5567菌株lmoF2365_0032基因编码的氨基酸序列与GenBank上公布的6株不同来源的lmoF2365_0032基因編码的氨基酸序列进行同源性比对,结果(图5)显示,LM5567菌株lmoF 2365_0032基因编码的氨基酸序列与02-6680菌株(4b,奶酪,加拿大)、10-0811菌株(1/2b,螃蟹,加拿大)、10-0809菌株(4b,粪便,加拿大)、81-055菌株(4b,脑脊髓液,加拿大)、02-1103菌株、02-1792菌株(4b,奶酪,加拿大)、81-0861菌株(4b,卷心菜,加拿大)的相似性均为94.7%。
2.6 lmoF2365_0032蛋白的二级和三级结构预测
通过SOPMA软件对LM5567菌株lmoF2365_0032蛋白的二级结构进行预测,结果(图6)发现,lmoF2365_0032蛋白的二级结构包含15.53%的α-螺旋(含32个氨基酸),37.86%的无规则卷曲(含
78个氨基酸),36.89%的延伸链结构(含76个氨基酸)和9.71%的β-转角(含20个氨基酸)(图6)。通过SWISS-MODEL分析平台预测该蛋白的三级结构(图7),进一步验证了二级结构的预测。
3 讨论
单核细胞增生李斯特菌广泛存于自然界、食品加工各环节,能够耐受高盐、高渗、高温、酸性和碱性环境。其中,在酸性环境中的存活尤为关键,LM经污染的饲草料和食品等进入人和动物机体后,首先遭遇的就是胃肠道的低pH值环境,被巨噬细胞吞噬后,吞噬体内也有一个酸化过程,过酸环境不利于该细菌的存活,因此单核细胞增生李斯特菌必须具备有效维持pH值平衡的机制,以保证其在酸性环境中存活[8]。arc基因编码的精氨酸脱亚胺基酶系统(ADS)中的蛋白在LM调节酸平衡中发挥重要作用[9]。lmoF2365_0032基因是编码精氨酸鸟氨酸逆向转运蛋白的基因之一,因此推测其有助于细菌在酸性条件下存活。
ADS存在于革兰阳性菌和革兰阴性菌中。ADS作用的发挥需要一个转运蛋白和3个酶,首先由arcD编码的精氨酸鸟氨酸逆向转运蛋白从细胞外摄取精氨酸进入细胞内,接着由arcA编码的精氨酸脱亚氨酶(ADI)将精氨酸分解为鸟氨酸和氨,鸟氨酸在arcB编码的鸟氨酸转氨甲酰基酶的作用下生成鸟苷和氨甲酰磷酸盐,然后由arcC编码的氨甲酸激酶将氨甲酰磷酸盐的磷酸基团转移给二磷酸腺苷(ADP)产生能量腺嘌呤核苷三磷酸(ATP)[10]。由此可见,一方面细菌可通过ADS摄取胞外的精氨酸产生ATP,促进细菌生长繁殖,另一方面当细菌处于酸性环境时,ADS产生的氨可与细胞质中的氢离子结合为铵离子,从而提高细胞质中的pH值,在一定程度上减轻酸应激的伤害。
ADS除了在抗酸应激中发挥重要作用外,在许多细菌的致病性中也具有重要作用。Evans等的沙门菌生长动力学试验表明,arcA参与鞭毛的生物合成、趋化因子的基因表达调控[11]。Ma等在禽大肠杆菌(APEC)雏鸭模型中发现,APEC野毒株致死率高于arcA缺失株;与野毒株相比,arcA缺失株脑组织的载菌量大幅度下降,说明arcA基因的缺失可显著降低APEC在雏鸭模型中的毒力[12]。Thao等通过血清杀伤试验表明,arcA是嗜血杆菌抗血清和逃避补体的必要条件[13]。Gupta等发现,肺炎链球菌arcD的缺失增强了A549细胞和巨噬细胞的吞噬能力,且免疫荧光检测发现,该突变体还表现出荚膜受损,表明arcD不仅可能在抗吞噬中发挥作用,而且可能参与荚膜的形成[14]。陈健舜等构建了李斯特菌10403的arcA、arcB、arcD缺失株,发现缺失株对Hela细胞的黏附和侵袭没有影响,但是会降低小鼠的脾内载菌数,使半数致死剂量(LD50)高于亲本株,说明arcA、arcB和arcD与细菌毒力相关[15]。但食源性单核细胞增生李斯特菌lmoF2365_0032蛋白的功能未见相关报道。
本研究采用PCR方法从食源性单核细胞增生李斯特菌LM5567菌株中克隆了lmoF2365_0032基因,经测序、拼接、分析比对得到,lmoF2365_0032基因的全长序列为811 bp。同源性比对分析显示,LM5567菌株lmoF2365_0032基因的核苷酸序列与4b血清型菌株相似性较高,为99.8%,其次与1/2b血清型菌株相似性为99.7%,其编码的氨基酸序列相似性为94.7%。LM5567菌株lmoF2365_0032蛋白的理化性质分析显示,该蛋白是一种偏碱性(等电点理论值为8.51)、稳定(不稳定指数为23.31<40.00,为稳定性蛋白)、亲水性蛋白质。lmoF2365_0032蛋白结构域分析显示,该蛋白为跨膜蛋白,无信号肽区域,属于非分泌蛋白;lmoF2365_0032蛋白的二级结构预测结果显示,lmoF2365_0032蛋白的二级结构包含15.53%的α-螺旋(含32个氨基酸),37.86%的无规则卷曲(含78个氨基酸),36.89%的延伸链结构(含76个氨基酸)和9.71%的β-转角(含20个氨基酸),其中以无规则卷曲和延伸链结构为主;lmoF2365_0032蛋白具有高度亲水性,由此推断该蛋白与抗酸应激有关。
本研究首次克隆了lmoF2365_0032基因,并利用生物学软件对其结构和功能进行预测,为今后研究该蛋白与LM5567菌株致病作用的关系提供了理论基础。
参考文献:
[1]丘增萍,谢 强,况 伟,等. 2013年梅州市餐饮业自制食品微生物污染检测与分析[J]. 中国药事,2015,29(4):353-356.
[2]马 微,潘秀霞,李苏龙,等. 进口俄罗斯食品中食源性微生物的检测分析[J]. 食品安全质量检测学报,2016,7(12):5069-5073.
[3]Swaminathan B,Gerner-Smidt P. The epidemiology of human listeriosis[J]. Microbes and Infection,2007,9:1236-1243.
[4]张春琳,张家国. 单增李斯特菌增殖的影响因素研究[J]. 食品研究与开发,2011,32(7):48-53.
[5]吴学林,曹树珠,马 勋. 单核细胞增生性李斯特菌srtA基因的克隆与原核表达[J]. 石河子大学学报(自然科学版),2015,33(2):196-200.
[6]Sakanaka A,Kuboniwa M,Takeuchi H,et al. Arginine-ornithine antiporter arcD controls arginine metabolism and interspecies biofilm development of Streptococcus gordonii[J]. Journal of Biological Chemistry,2015,290(35):21185-21198.
[7]Maury M M,Tsai Y H,Charlier C,et al. Uncovering listeria monocytogenes hypervirulence by harnessing its biodiversity[J]. Nature Genetics,2016,6(48):308-311.
[8]Cotter P D,Hill C. Surviving the acid test:responses of gram-positive bacteria to low pH[J]. Microbiology & Molecular Biology Reviews Mmbr,2003,67(3):429-453.
[9]Wong S M S,Michael F S,Cox A,et al. ArcA-regulated glycosyltransferase lic2B promotes complement evasion and pathogenesis of nontypeable Haemophilus influenzae[J]. Infection & Immunity,2011,79(5):1971-1983.
[10]叶保国,滕士勇,李新白,等. Arc基因表达及甲基化与幼龄鼠和成年鼠空间记忆形成的相关性[J]. 中国生物制品学杂志,2013,26(3):363-367.
[11]Evans M R,Fink R C,Vazquez-Torres A,et al. Analysis of the ArcA regulon in anaerobically grown Salmonella enterica sv. typhimurium[J]. BMC Microbiology,2011,11(1):58.
[12]Ma J L,An C X,Jiang F W,et al. Extraintestinal pathogenic Escherichia coli increase extracytoplasmic polysaccharide biosynthesis for serum resistance in response to bloodstream signals[J]. Molecular Microbiology,2018,110(5):689-706.
[13]Thao S,Brandl M T,Cater M Q. Enhanced formation of shiga toxin-producing Escherichia coli persister variants in environments relevant to leafy greens production[J]. Food Microbiology,2019,84:103241.
[14]Gupta R,Yang J,Dong Y M,et al. Deletion of arcD in Streptococcus pneumoniae D39 impairs its capsule and attenuates virulence[J]. Infection and Immunity,2013,81(10):3903-3911.
[15]陳健舜,方维焕,张才乔. 精氨酸亚胺酶系统对单增李斯特菌致病力的影响及其分子作用机制[C]//2010中国博士后生命科学学术论坛,2010.