水稻OsCatB敲除突变体的构建及耐逆性初步分析
2020-09-24赵李剑唐冬英刘选明林建中
刘 珊,赵李剑,刘 聪,邓 勇,黄 剑,唐冬英,刘选明,林建中
(湖南大学生物学院植物功能基因组学和发育调控湖南省重点实验室,中国湖南长沙410082)
植物在生长发育过程中常常会遭受一些不良因素的侵袭,从而造成植物正常生长状态被破坏甚至死亡。我们把导致植物生长发育过程被打断或破坏的环境条件称为逆境胁迫[1~3]。根据胁迫来源的不同,逆境胁迫分为两大类,即生物胁迫和非生物胁迫。其中,最常见的高温、高盐、高碱等属于非生物胁迫,会导致植物体内产生大量的活性氧(reactive oxygen species,ROS),如超氧阴离子自由基(O2.-)、羟基自由基(·OH)和过氧化氢(H2O2)等[4~6]。ROS本身能够作为一种重要的信号分子参与许多生物学过程,包括耐受生物与非生物逆境。但是,ROS的过量积累会氧化并破坏膜脂、蛋白质和核苷酸等细胞组分,若不能及时清除将会严重影响植物新陈代谢,抑制植物生长与发育,甚至可能直接造成植物的死亡[7~10]。
为了消除ROS的毒害,植物已进化出了一套有效清除ROS的酶和非酶清除机制。ROS的清除酶类包括超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)、过氧化氢酶(catalase,CAT)、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)和谷胱甘肽过氧化物酶(glutathione peroxidase,GPX)。ROS的非酶清除分子,即抗氧化分子,主要为抗坏血酸(ascorbic acid,AsA)、还原型谷胱甘肽(glutathione,GSH)、生育酚和黄酮类化合物等。这些酶类和非酶类分子协同作用使ROS保持动态平衡,维持ROS在细胞信号传导中所需要的正常水平[11]。CAT是一个非常重要的抗氧化酶,能够催化H2O2降解为H2O和O2,在植物非生物逆境响应中起着关键作用。有研究表明,植物在逆境条件下产生的过量ROS一般都是由CAT来大量清除的[12]。CAT虽然在细胞内的浓度与亲和力偏低,但是清除效率高,反应快,在植物处于逆境胁迫时,可及时为植物减低氧化损伤[13]。水稻中的CAT家族共含有3个同工酶成员,分别为CatA、CatB和CatC,在拟南芥中也包含3个成员,即Cat1、Cat2和Cat3[14]。大麦和烟草的遗传研究显示,大麦Cat2对叶片的生长发育很重要,而烟草中的Cat2则可清除由氧化胁迫产生的H2O2[15]。相关研究对拟南芥Cat2基因的T-DNA插入突变体展开分析发现,Cat2负责拟南芥中大部分的叶片过氧化氢酶活性,是光呼吸的重要参与者[16]。在水稻中,CatB属于第三类过氧化氢酶,大量在非光合组织、愈伤组织、根和种子中表达,其可能与脂肪酸β-氧化合成有关[17]。但是,关于CatB基因在水稻抗逆境中的作用尚未有人作出清楚的阐述。
本研究首先建立了OsCatB的CRISPR/Cas9基因敲除体系,并筛选鉴定到纯合突变体catb。然后通过模拟盐、高温和氧化胁迫,分析了突变体catb的生理和生化表型,发现在高温和氧化胁迫下突变体catb的存活率比野生型(wild type,WT)显著降低,体内生理指标CAT酶活与野生型相比也显著降低,同时突变体在表型上也呈现出一种更容易受损伤的状态。研究结果初步说明,缺失了CatB基因的水稻突变体在氧化和热胁迫下,其生长状态和抗逆境能力都低于野生型,意味着Os-CatB参与了水稻的氧化和热胁迫响应。
1 材料与方法
1.1 供试材料
水稻材料为粳稻品种Kitaake(Oryza sativa L.cv.Kitaake),用于水稻转化、克隆、RNA提取等实验。水稻种子保存于4℃冰箱。
1.2 CRISPR/Cas9载体的构建
根据 Liu等[18]的方法,利用 CRISPR-P 2.0(http://crispr.hzau.edu.cn/)在线工具设计靶标。以OsCatB的第二个外显子作为基因编辑的靶点,遵循gRNA设计基本原则筛选出单靶标CatB-U6a:5′-AACAACTCCGCCCTCACCGT-3′;采用 Primer 5.0软件设计引物 CatB-U6a-F和 CatB-U6a-R(表1),克隆靶标片段;将靶标连上CRISPR/Cas9中间载体(华南农业大学刘耀光教授提供),随后利用内切酶Nde I和BamH I(ThermoFisher公司,美国)切下含有靶标的CRISPR表达原件片段;将目标片段与植物载体pCAMBIA1301-GUS(实验室保存)连接后转化进大肠杆菌(E.coli)DH5α感受态中,挑选阳性克隆,交由生工生物工程(上海)股份有限公司进行测序确认。
1.3 水稻转化
水稻转化方法在文献[19]的基础上略有修改,具体如下:将重组质粒pCAMBIA1301-GUS经电击法转入农杆菌EHA105(本实验室保存),再通过农杆菌浸染粳稻品种Kitaake愈伤组织,获得Os-CatB敲除突变体catb。
1.4 PCR鉴定
参照文献[20]的方法,提取转化株系幼苗(T1)叶片的DNA,根据表达载体上的潮霉素筛选标记基因(Hyg)设计引物Hyg-F和Hyg-R(表1),然后通过PCR扩增和电泳检测进行阳性转基因植株的鉴定。提取已鉴定的阳性植株(T1)叶片DNA,根据基因编辑靶标位点附近的序列设计引物CatBTC-F和CatB-TC-R(表1),然后通过PCR扩增目的片段,交由生工生物工程(上海)股份有限公司进行测序,最后将测序结果与野生型CatB的序列进行比对以确认具体编辑方式。
1.5 GUS组织化学染色
参考Jefferson等[21]的方法配制染色液,将催芽6 d的水稻幼芽(T1)浸泡于GUS染色液中,随后置于37℃培养箱黑暗处理1~2 d,期间每隔3 h观察其颜色变化。
1.6 免疫印迹检测
免疫印迹检测参照Zhou等[22]的方法进行,主要步骤如下:将catb突变体的植物总蛋白质上清经SDS-PAGE电泳后恒流转移到PVDF膜上,用5%脱脂奶粉室温封闭1 h,弃封闭液;然后使用CatB的多克隆抗体(本实验室制备)室温孵育1 h(或4℃过夜),TBST洗膜3次;再使用二抗(鼠抗IgG,HRP)室温孵育1 h,TBST洗膜3次;TBS洗膜1次后显影。
1.7 苗期的逆境处理
盐胁迫处理:将生长21 d的突变体和野生型幼苗置于NaCl浓度为140 mmol/L的水培营养液[23]中,处理8 d,随后用正常的水培营养液恢复培养20 d,观察植株表型并统计野生型和突变体的存活率。
热胁迫处理:以45℃持续16 h(光照)和35℃持续8 h(黑暗)的交替温度处理生长21 d的突变体和野生型幼苗7 d[24],然后于28℃恢复培养20 d,观察植株表型并统计存活率。
氧化胁迫处理:参考文献[25]的方法使用甲基紫精(methylviologen,MV)(Sigma公司,美国)对水稻发芽材料进行氧化胁迫处理。略作修改的方法如下:将催芽5 d后的水稻材料分别转移到正常的1/2 MS培养基(对照组)和含有2 μmol/L MV的1/2 MS培养基(实验组)中处理6 d,然后观察表型并拍照。
1.8 CAT酶活和H2O2含量的测定
对于高温和盐胁迫处理的生长21 d的水稻幼苗,分别取开始处理时的 0 d、1 d、3 d、5 d、7 d和 0 d、2 d、4 d、6 d、8 d 的样品,使用 pH 7.5 的0.02 mmol/L磷酸缓冲液作为基本的植物上清蛋白质提取液。使用CAT检测试剂盒(S10051;碧云天公司)和H2O2含量检测试剂盒(BC3590;索莱宝公司)进行测样,设置三组平行样。
1.9 统计学处理
2 结果与分析
2.1 OsCatB敲除突变体catb的获得及鉴定
CRISPR/Cas9表达载体的示意图如图1A所示,其中含有由35S启动子驱动表达的潮霉素筛选标记基因(Hyg)和GUS报告基因(GUS),而单靶点gRNA则由OsU6启动子驱动。测序结果显示,该CRISPR/Cas9表达载体构建成功。通过农杆菌介导水稻转化法,获得56株T1杂合子。潮霉素抗性基因Hyg的PCR鉴定结果显示,阳性植株中目的基因大小与预期相同,而阴性对照植株中无相应条带,初步确定这些阳性植株为目的转基因植株(图1B)。由于在T-DNA区段中含有35S驱动的GUS报告基因(图1A),为了进一步快速鉴定阳性植株,我们采用GUS染色液对萌发种子进行了染色筛选,发现凡是含有Hyg标记基因的萌发种子均着色,而不含有Hyg标记基因的萌发种子则无法着色(图1C)。靶点测序及比对结果发现,3个独立的转基因株系在PAM(protospacer adjacent motif)附近发生了3种方式的编辑:catb-1在目标位置缺失了胞嘧啶(C);catb-2在目标位置缺失了52个碱基;catb-3在目标位置插入了鸟嘌呤(G)(图1D)。为进一步了解上述3个catb突变体中CatB蛋白的表达情况,我们采用抗CatB的多克隆抗体进行了免疫印迹检测,发现3个突变株系中CatB蛋白的积累水平均比野生型显著降低(图1E)。需要说明的是,catb突变体在CatB位置仍可检测到微弱的信号,但是其信号显著弱于野生型,这可能是由于所用抗CatB的多克隆抗体非特异性地检测到了部分CatA和CatC。综上可知,文中所构建的catb突变材料为真正的OsCatB基因敲除突变体,可用于后续的生理和生化等表型分析。

表1 PCR引物序列Table 1 Nucleotide sequences of PCR primers
2.2 突变体catb的耐盐性分析
选取突变体catb-1和catb-3作为实验材料,以未转化的Kitaake作为野生型对照,在长日照培养箱培养至3周左右时,用含140 mmol/L NaCl的水培营养液进行盐胁迫处理。表型观察结果显示,野生型和突变体经盐胁迫处理后,表型并无明显差别(图2A),说明OsCatB在水稻的盐响应中不起作用或作用不明显。恢复培养20 d后进行存活率统计,结果显示catb-1和catb-3的存活率维持在15%左右,比野生型的存活率(16%左右)略微下降,但是仍没有显著差异(图2B)。已知CAT对于H2O2的消除和氧化还原的维持非常重要[26],为了研究catb突变体在应对盐胁迫时对体内CAT 酶活的影响,分别取盐处理 0 d、2 d、4 d、6 d和8 d 5个时间点下幼苗的根和地上部分,检测其CAT酶活和H2O2含量。结果如图2C~F所示,未受胁迫处理(0 d)时,catb突变体与野生型的CAT酶活和H2O2含量几乎无明显变化;在140 mmol/L NaCl处理下,野生型的CAT酶活有所增加,虽然catb中CAT酶活增强的幅度不明显,但与野生型相比基本无显著性差异,而H2O2含量变化与之相反,且差异也基本不显著。该结果说明,OsCatB的敲除对水稻盐胁迫响应的影响不明显,猜测CAT家族成员间具有一定的功能冗余,即当OsCatB缺失时,其他成员(OsCatA和OsCatB)可能存在一定的补偿机制,以保持体内的H2O2含量不变。
2.3 突变体catb的耐高温分析
21 d大小的突变体和野生型幼苗经45℃16 h和35℃8 h循环处理7 d后,置于28℃光照培养箱恢复培养20 d。表型观察发现,catb-1和catb-3与野生型在高温处理7 d以后的生长状态有明显差异,野生型还尚存一小部分绿色的叶片,但是突变体已经全都变黄呈现枯萎状(图3A)。恢复培养20 d后统计其存活率发现,catb突变体的存活率(5%~8%)显著低于野生型(15%)(图3B),说明突变体比野生型更易遭到热胁迫的损伤和破坏,OsCatB正调控水稻的热胁迫响应。为了进一步探究OsCatB的敲除对植株在热胁迫下的生理生化的影响,我们测定了热处理0 d、1 d、3 d、5 d和7 d 5个时间点下幼苗根部和地上部分的CAT酶活与H2O2含量。检测结果显示,高温处理时catb突变体和野生型的根与地上部分的CAT酶活均有升高的趋势,但是catb突变体中CAT酶活升高的趋势显著低于野生型,这种差异在根中更显著(图 3C~D)。该结果与已有文章的报道[17]一致,因为水稻OsCatB主要在非光合组织、愈伤组织、根和种子中表达。同时研究结果也发现,在正常条件(0 d)下野生型与catb突变体株系的CAT酶活和H2O2含量均无显著的差异,只有在高温处理之后,catb突变体的CAT酶活才开始显著低于野生型,而其H2O2含量则显著高于野生型(图3C~F),这些数据与图3A的表型特征相符合。由此我们初步推测,水稻植株受到热胁迫时体内会积累超量的ROS(主要是H2O2的形式),由于catb突变体中OsCatB的功能缺失而无法及时清除超量H2O2,最终导致其耐受性下降。同时,该结果也进一步说明,OsCatB基因可以在一定程度上增强热胁迫的抗性,在加强水稻细胞对抗热胁迫中发挥重要的作用。
2.4 突变体catb的耐氧化胁迫分析
于前所述,当catb突变体处于盐和热胁迫下时,其CAT酶活和H2O2含量与野生型相比均发生了变化,而且这种变化幅度在热胁迫下更大,因此我们猜想水稻OsCatB可能通过及时清除过量积累的ROS来正调控水稻的氧化胁迫响应。为了证实该猜想,我们使用甲基紫精(MV)模拟氧化胁迫来处理发芽5 d的水稻材料,结果显示,对照组中野生型和catb突变体在植株高度上无明显差异,但是在2 μmol/L的MV处理6 d后,突变体的生长受到显著抑制,与野生型相比呈现出一种更矮小萎黄的状态(图4)。由此,我们初步认为OsCatB也正调控水稻的氧化胁迫响应。
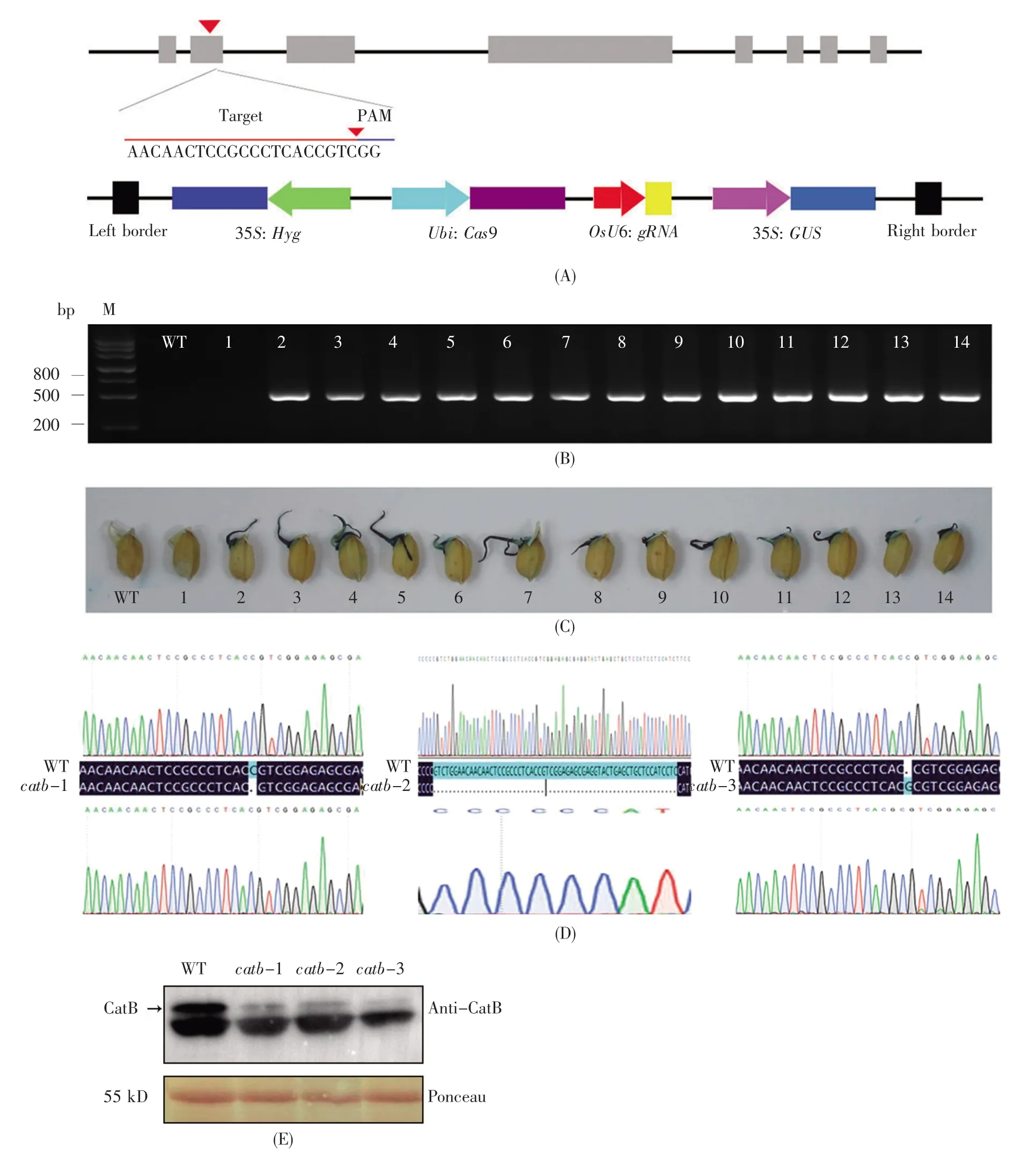
图1 catb突变体的创制与鉴定(A)CRISPR/Cas9表达载体的构建;(B)转化苗中潮霉素抗性基因(Hyg)的PCR检测。M为DNA分子量标准,WT为野生型,1~14为不同转基因株系的T1代幼苗;(C)发芽种子的GUS染色鉴定;(D)3种突变OsCatB基因的目标区间的测序图谱和序列;(E)免疫印迹检测突变体OsCatB蛋白的表达。Fig.1 Creation and identification of catb mutants(A)Construction of CRISPR/Cas9 expression vector;(B)PCR analysis of the hygromycin resistant gene Hyg in transgenic lines.M represents DNA marker,WT represents wild type plants,and 1~14 represent the T1seedlings of various transgenic lines;(C)GUS staining identification of germinated seeds;(D)Chromatograms and sequences of three mutated OsCatB genes in the desired region;(E)Immunoblot analysis of OsCatB in catb mutants.
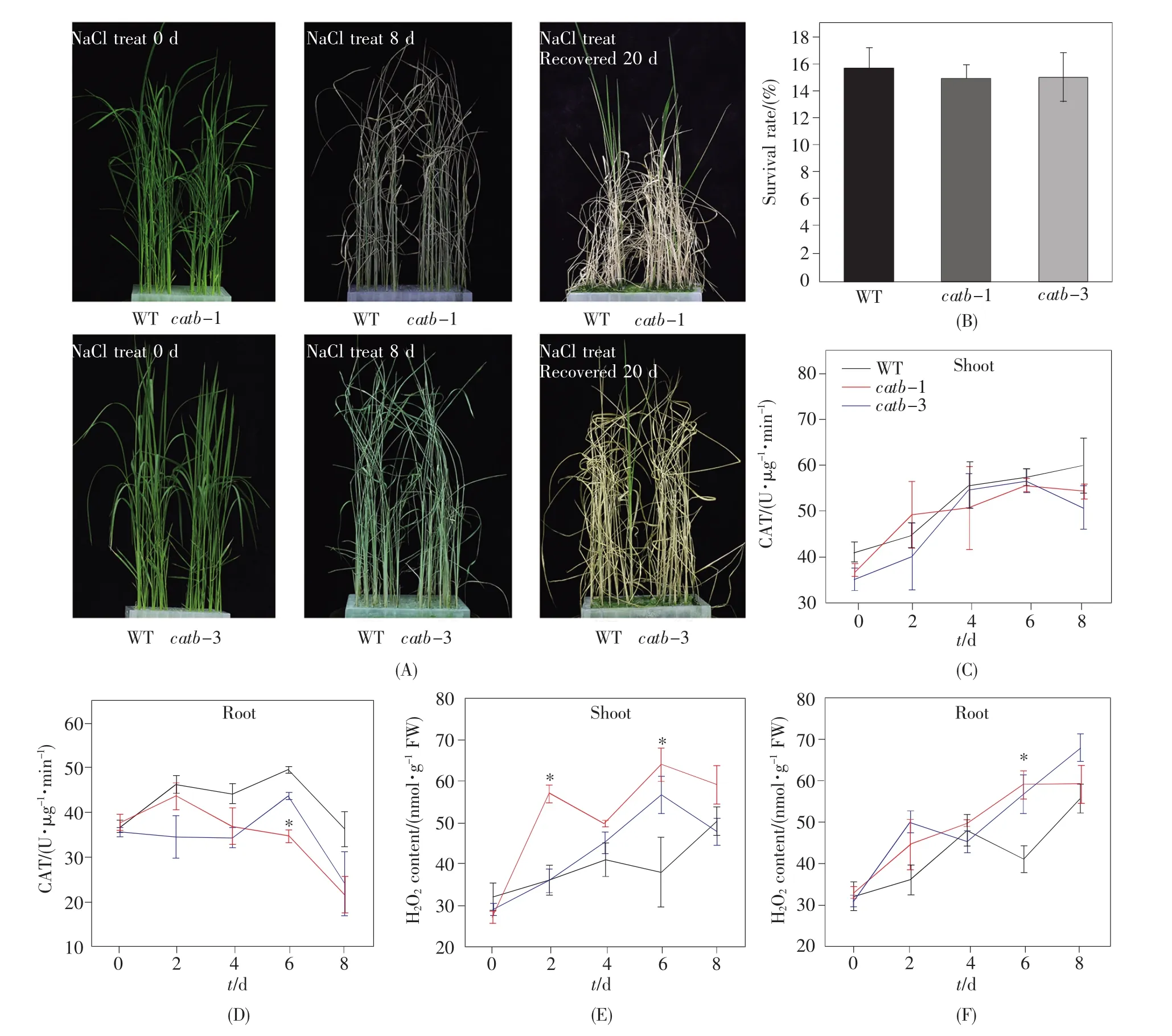
图2 幼苗期catb突变体对盐胁迫的响应分析(A)盐胁迫下catb和野生型幼苗的表型。生长21 d的水稻幼苗用140 mmol/L的NaCl处理8 d,随后用常规营养液恢复培养20 d;(B)恢复培养20 d后catb和野生型幼苗的存活率;(C~D)catb和野生型幼苗地上部分和根部在不同盐处理时间的CAT酶活性;(E~F)catb和野生型幼苗地上部分和根部在不同盐处理时间的H2O2含量。图B~F中的数值代表的是平均值±标准差(n=3,*表示与 WT 相比 P≤0.05,t检验)。Fig.2 Response analysis of catb mutants to salt stress at seedling stage(A)Phenotype of catb and WT seedlings under salt stress.Twenty-one-day-old seedlings were treated with 140 mmol/L NaCl for 8 d,and then recovered for 20 d;(B)Survival rates of catb and WT seedlings after recovery for 20 d;(C~D)CAT activities of shoots and roots of catb and WT seedlings treated with salt stress at the indicated time points;(E~F)H2O2contents of shoots and roots of catb and WT seedlings treated with salt stress at the indicated time points.Data in B~F are presented as mean ± standard deviation(n=3,*P≤0.05 vs.WT,Student’s t-test).
3 讨论
在植物生长与发育过程中,盐、高温和干旱等逆境胁迫条件下产生的离子毒害和渗透胁迫会打破其ROS生成和清除之间的平衡,使体内产生的ROS多于代谢的ROS,造成氧化应激[27]。过氧化氢酶(CAT)是一类非常重要的抗氧化酶,能够消除植物中因逆境胁迫而积累的ROS,使之维持合适浓度,因此在植物的非生物逆境响应中起着关键作用[28]。为了探究CatB对逆境的响应情况,我们建立了OsCatB基因的CRISPR/Cas9基因敲除体系,并成功获得了OsCatB基因敲除的3个独立纯合突变株系catb-1、catb-2和catb-3(图1)。由于catb-2的种子量不够,所以在后续实验中我们仅选取catb-1和catb-3用于逆境胁迫的生理及生化分析。
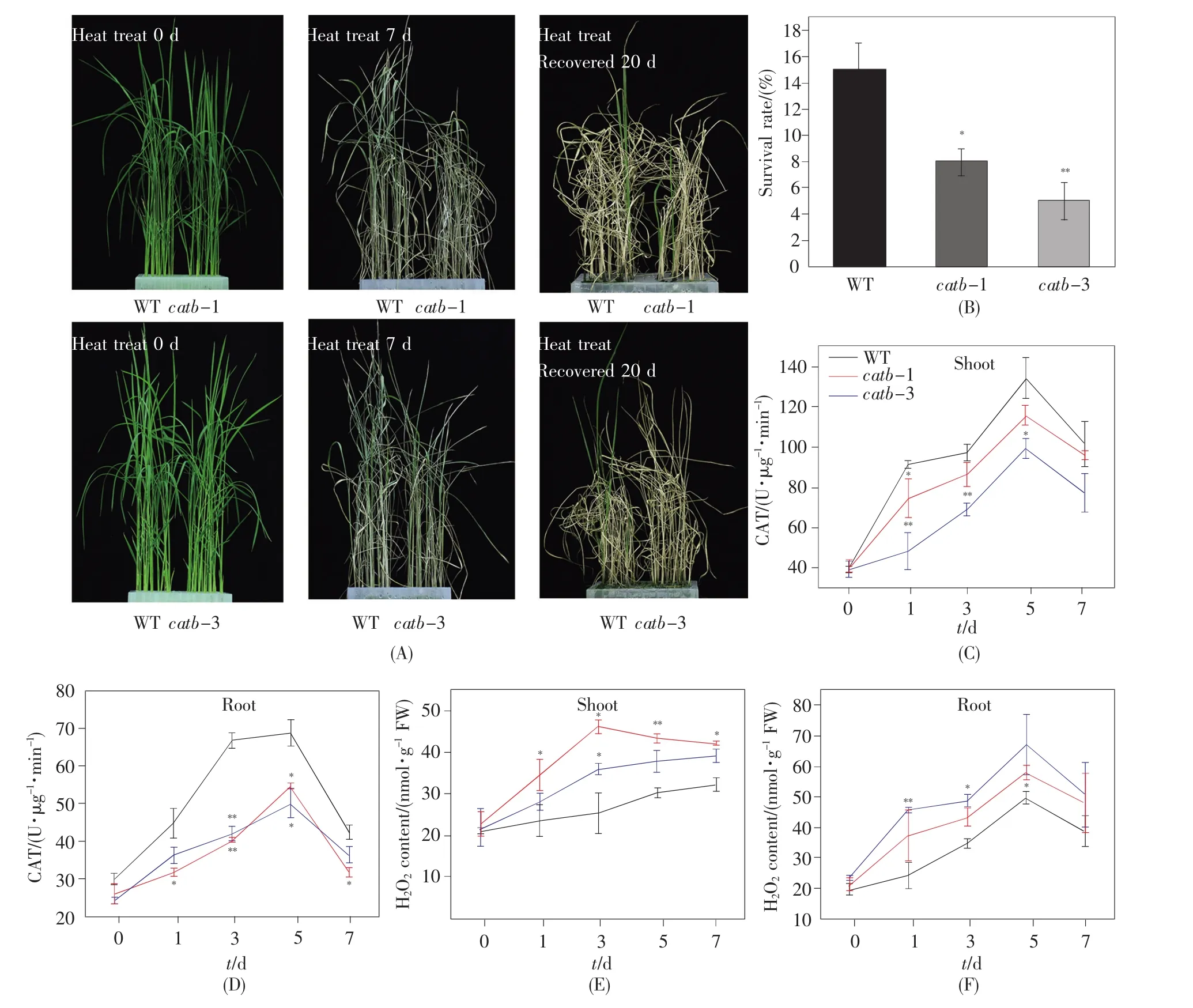
图3 幼苗期catb突变体对热胁迫的响应分析(A)热胁迫下catb和野生型幼苗的表型。生长21 d的水稻幼苗用45℃16 h(光照)和35℃8 h(黑暗)循环处理7 d,然后28℃恢复培养20 d;(B)恢复培养20 d后的catb和野生型幼苗的存活率;(C~D)catb和野生型幼苗地上部分和根部在不同热处理时间的CAT酶活性;(E~F)catb和野生型幼苗地上部分和根部在不同热处理时间的H2O2含量。图B~F中的数值代表的是平均值±标准差(n=3,t检验),*和 **分别表示与 WT 相比 P≤0.05、P≤0.01。Fig.3 Response analysis of catb mutants to heat stress at seedling stage(A)Phenotype of catb and WT seedlings under heat stress.Twenty-one-day-old seedlings were treated with the temperature cycle of 16 h at 45℃(light)and 8 h at 35℃(dark)for 7 d,and then recovered at 28℃for 20 d;(B)Survival rates of catb and WT seedlings after recovery for 20 d;(C~D)CAT activities of shoots and roots of catb and WT seedlings treated with heat stress at the indicated time points;(E~F)H2O2contents of shoots and roots of catb and WT seedlings treated with heat stress at the indicated time points.Data in B~F are presented as mean ± standard deviation(n=3,Student’s t-test).*and**represent P≤0.05 and P≤0.01,respectively.

图4 幼苗期catb突变体对MV的响应分析(A)MV处理下catb和野生型材料的表型。催芽5 d的水稻材料用正常的1/2 MS培养液或含有2 μmol/L MV的1/2 MS培养液处理6 d。标尺=1 cm;(B)0 μmol/L或2 μmol/L MV处理6 d后catb和野生型幼苗的高度。图中数值代表的是平均值±标准差(n=20,t检验),*和 **分别表示与 WT 相比 P≤0.05、P≤0.01。Fig.4 Response analysis of catb mutants to MV at seedling stage(A)Phenotype of catb and WT seedlings treated with MV.Following pre-germination treatment for 5 days,the germinated seeds were transplanted into either 1/2 MS medium or 1/2 MS medium supplemented with 2 μmol/L MV for 6 d.Bar=1 cm;(B)Seedling heights of catb and WT seedlings treated with 0 μmol/L or 2 μmol/L MV for 6 d.Data are presented as mean ± standard deviation(n=20,Student’s t-test).*and**represent P≤0.05 and P≤0.01,respectively.
CAT家族在水稻中有CatA、CatB和CatC等3个成员,而在拟南芥中也有Cat1、Cat2和Cat3等3个成员,各成员在不同逆境响应中起着不同的作用[14]。在拟南芥中,Cat3主要正调控干旱胁迫响应,其可以被钙依赖蛋白激酶CPK8磷酸化而激活,进而降解因干旱积累的H2O2,以增强植株的抗旱性[29]。近期,Zhou等[23]发现CatC能正调控水稻的耐盐性,在水稻体内ROS的清除中起着重要作用。CatC在盐胁迫条件下被类受体胞质激酶STRK1磷酸化而激活,激活的CatC可清除因盐胁迫诱导积累的过量H2O2,从而提高水稻的耐盐性。在本研究中,盐胁迫处理下OsCatB基因敲除突变体catb的表型与野生型对比不明显,只显示出微弱的盐敏感性;同时,CAT酶活和H2O2含量的差异也不显著(图2C~F),甚至在5个时间点中存在几个点的野生型和突变体数据很相近。因此,我们猜测在清除因盐胁迫累积的ROS时,CatB仅起辅助作用,而起主导作用的还是CatC。此外,我们还分析了在高温与氧化胁迫下野生型和catb突变体的表型、CAT酶活和H2O2含量,发现与野生型相比,突变体均表现出对高温和氧化胁迫的超敏感,而且体内的CAT酶活显著降低,H2O2的累积量显著升高(图3和图4)。因此,我们可以推测,水稻体内OsCatB基因的敲除,导致了由高温和氧化胁迫所积累的ROS无法及时清除,从而造成植物自身的氧化损伤。该结果也同时说明,OsCatB具有正调控水稻耐高温和耐氧化胁迫的能力,在作物的遗传改良中具有一定的应用潜力,但是与其相关的分子作用机理仍需要进一步研究。
总之,本研究初步发现OsCatB对水稻盐胁迫响应的调控不明显,主要参与了高温和氧化胁迫响应,并正调控水稻的高温和氧化胁迫耐受性。同时,该研究为进一步探究OsCatB参与逆境响应的分子机制和培育抗逆水稻品种提供了一定的理论指导和遗传材料。
