达尔文主义的“日食”
2020-09-23郭建崴
郭建崴
我们已经用连续几期介绍了自《物种起源》问世以后半个世纪左右的时间段里一些学者对捍卫与发展进化论所作出的贡献。但这并不代表那个时代的全景。
实际上,“达尔文主义”的整个理论体系在很长的一段时间内并没有完全被更多的学者充分理解与全面接受,即使那些支持进化论的人。大家普遍认可的只是进化本身以及“共同祖先”等观念,其他方面则在不同学者那里存在着不同的疑惑与分歧。其中,最大的挑战是针对“自然选择”以及一脉相承而来的“渐变”理论。尤其到了1882年达尔文逝世之后,除了华莱士、魏斯曼以及生物统计学家威尔登(Raphael Weldon,1860~1906)和皮尔森(Karl Pearson,1857~1936)等少数支持者和发展者之外,自然选择学说几乎被漠视甚至抛弃,更多学者试图用其他的机制来说明进化如何产生。直到20世纪30年代后期至40年代,当一批新的学者将进化论与遗传学统一而催生了“现代达尔文主义”之后,上述状况才终于扭转。“现代达尔文主义”的开创人之一、“达尔文的斗犬”之孙朱利安·赫胥黎(Julian Huxley,1887~1975)将这一段“自然选择蒙难史”称为“达尔文主义的‘日食’”。
“日食”期间,围绕着对达尔文的批评形成了许许多多的观点和学说,其中最具广泛影响的有三种:新拉马克主义、直生论和突变论。
新拉马克主义
前文曾经介绍过,拉马克认为,各类简单的生物体在生命史上的不同时间以“自然发生”的方式起源于无机界,不同类型的原始生命通过漫长岁月的进化,逐渐发展成为各类复杂的生物。推动这种进化的动力,首先是生物具有一种不断地增加结构复杂性和完美性的天生趋势,其次是生物具有对环境变化的反应能力。当环境不变时,生物进化就是一种完美化的发展;如果环境改变,生物就产生适应环境的需要和行为习性并引起有关的适应性进化。而进化的机制,则是器官的用进废退和获得性状遗传。
拉马克生前和逝世后相当长的一段时期,其进化论不但没有被接受,甚至常常被作为反面教材。究其原因,一方面是因对神学的冲击而遭致基督教阵营的反对,另一方面是其科学上的“先天不足”。但是当达尔文以《物种起源》促使越来越多的人对生物进化事实广泛接受之后,一些对自然选择学说不满意的学者却又从拉马克的一些观点中找到了共鸣,从而出现了“新拉马克主义”学派。
“新拉马克主义”一词由美国学者帕卡德(A.S.Packard)创造,阵营中包括有著名古生物学家科普(E.D.Cope)和奥斯本(H.F.Osborn)以及爱米尔(T. G. H. Eimer)、唐得克(Le Dantecl)、耐格里(C.Ngeli)、汪德比尔特(Wintrebert)等一干当时的大家。获得性状遗传的观点是这个学派的核心,但是对拉马克学说本身包含的其他诸多要素,不同学者却是各持己见。在今天看来,这些见解或缺少事实的证据和实验的证明因而多为推测,或在认识上存在片面性。
例如,科普认为生物体具有其固有的作为进化原动力的生长力,耐格里也以类似的观点提出了“完成化原理”,发展了定向进化的思想。科普把生物的变异分出理化变异和动作变异两类,认为前者是由环境的影响直接促成的,而后者是因生物受环境因素刺激而发生运动(动作),这些运动(动作)又引发了变异。进而科普认为,动作变异与生物的需要和意识密切相关,意识的根源在生命起源的时代即已存在。科普因此被称为是“心灵拉马克主义”的代表。耐格里则主张进化的原因是来自于生命内在的冲动。
奥斯本认为生物进化有一个预定的方向或目的,不管是有利还是有害,它们都沿着一定的方向发展。这些学者非但没有克服拉马克主义原有的生物由“内在愿望”驱动而“努力”向上发展的错误观念,反而在这条歧途上渐行渐远。
爱米尔倒是不相信心灵的作用,也不赞成用进废退的原理,只是坚信生物的进化主要靠环境的力量和获得性状遗传。对于定向进化,他认为其原因是动物的生活状况和环境的影响,如气候、食物等的影响;生物本身并非完全被动,它们各有适合于自身的个性,体现在某类变化的方式常为某类生物所固有。
客观地说,新拉马克主义在19世纪后期到20世纪初期的盛行体现了科学发展“螺旋式上升”的规律,因为自然选择学说自身在当时也面临对一些科学难题无法给出另人满意解答的困境,即便是达尔文,也没有将获得性状遗传排除在进化机制之外。身为达尔文主义者的斯宾塞和海克尔也认为,只有把自然选择理论与新拉马克主义结合才能正确地解释进化的原理。直到20世纪30年代后期至40年代,综合进化理论(也称为现代达尔文主义)确立之后,获得性状遗传的理论在真正的生物学领域才被最终抛弃,新拉马克主义也就随之偃旗息鼓。
然而在前苏联,新拉马克主义却因一个带有强烈政治色彩的人物——李森科(T. D.Lysenko)而续命。出于意识形态对抗的目的,在斯大林的大力支持下,李森科从1935年开始运用新拉马克主义理论发起了挑战“西方遗传学”的“阶级斗争”。他汇集了前辈学者米丘林在进行作物品种改良和引进的长期试验中的一些经验,并结合自己的一些想法而提出了“米丘林-李森科理论”,认定环境影响可以造成生物适应性的定向改变,否认“资本主义的”遗传学所揭示的遗传规律,进而把科学扭曲为一场政治闹剧,压制生物学在前苏联的发展长达30年之久,直到1964年才随着赫鲁晓夫的下台而结束。
直生论
直生论也是一种反对自然选择学说的理论,认为生物进化是有方向的直线式历程,不论环境条件如何,生物只是沿着既定的方向进化;而决定进化方向的动力,完全来自于生物体自身的潜在力量,自然选择对此并无作用;一些趋势一旦启动就不会停止,即使最终导致该类生物的灭绝。例如,菊石外壳卷曲的过程一直发展,直至最后的物种完全将身体封闭在里面而导致整个类群的灭绝;剑齿虎在进化历程中无法停止牙齿的增长,猛犸象也不能中止长牙的生长,最终它们各自的家族便灭绝于因此造成的生活障碍。
直生论源于拉马克主义,最早由爱米尔在1885年提出来的时候就是指定向进化,后来经科普、奥斯本和罗莎(D.Rosa)等学者的助力而得到发展。到了后来,直生论因过于强调内在的种系动力对进化的驱动作用,认为这种进化是非适应性的,与环境也没有关系,因而实际上也偏离了拉马克主义。
对于一个支系的动物,如果把它们按由古到新的顺序排列起来,确实可以看到其形态朝着一定方向发生变化的趋势。例如马的进化,随时代的变迁体型逐渐增大、指趾数逐渐减少,同时牙齿越来越复杂化;又如鲸,在进化的过程中骨盆逐渐退化、后肢最终消失。这种“定向进化”现象在比较解剖学上也有所表现,例如从鱼类向两栖类、爬行类和鸟类与哺乳类比较过去,脊椎骨的骨化度增强、头骨数目减少、心脏房室分化增加,等等。但是直生论的解释是错误的。
随着化石资料的日益丰富,古生物学家发现马的进化并不是单一直线式的,而是发生过分支。而且,上述那些现象完全可以用自然选择学说来解释。“直生论”最大的死穴是所谓的“内在动力”,没有任何相应的物质基础。
直到1925年,作为直生论者的美国古生物学家R.S.卢尔还在用爱尔兰麋(学名为巨大角鹿,Megaloceros giganteus)的例子攻击达尔文:“自然选择不能解释超特异性。”他所谓的“超特异性”就是对于爱尔兰麋的巨角这样的器官“由于选择达到完美状态后,却不能度过日常的危险情况而保障生物体生存下来”这种“超常”的“特异性”的定义。在他看来,爱尔兰麋是由形态较小、角较小的鹿进化而来,虽然最初角是有适应意义的,但是随着这一支系的进化,角的生长不受限制地越变越大;到了爱尔兰麋的阶段,角已经大到不再具有适应性了——大角使头骨因为重负而弯曲,而当大角碰到树就容易卡在树杈间、在湿地上活动时也容易使其陷入泥潭,它们便因大角的累赘而纷纷死掉并灭绝了。
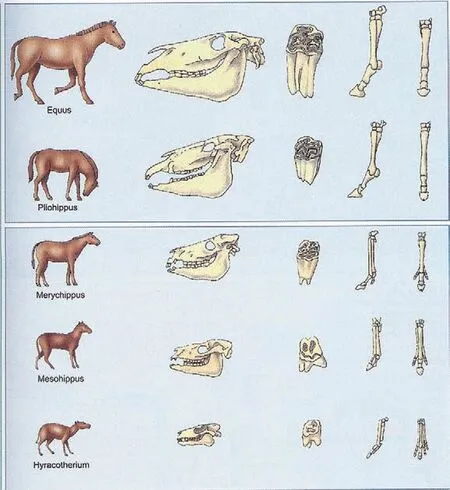
马的进化(王大锐供图)主干,被直生论拿来当作佐证。实际上在很多节点上进化出来的不仅仅是一个物种而是多个。因此马类家族的进化在整体上依然呈现为“进化树”的状态而非单一一条直线
今天来看这样的解释犹如天方夜谭。首先,从20世纪60年代开始,动物行为学家逐渐发现,角可以作打斗的武器,也可以在一定场合用作仪式性行为。角大的鹿说明地位高,可以获得雌性。因为保证生殖成功是最大的进化优势,所以对于角的选择压力很大,因而大的角无疑是具有适应意义的。再者,如果角是适应的,那么这种巨鹿在爱尔兰的灭绝原因为何呢?地质学、古生物学和古环境学的发展揭示,巨鹿在爱尔兰繁荣的时间很短,是在末次冰期的所谓阿累鲁间冰期,处于两个最冷期之间的短暂温暖期,从12000年前到11000年前仅仅延续了1000年。在此前一个冷期,爱尔兰与不列颠以及欧洲大陆因海平面降低而连接起来,爱尔兰麋便迁徙到了爱尔兰;在阿累鲁期,爱尔兰麋很好地适应了当地当时草木繁茂的广阔原野;但1000年过去之后,它们既不能适应伴随下一个寒冷期的到来而变成的亚北极冻土环境,也不能适应末次冰期结束后随着冰盖退去而发展起来的茂密森林。
灭绝最通常的原因就是由于物种不能足够迅速地适应气候等环境条件或物种间竞争条件的改变,爱尔兰麋如斯,猛犸象亦然,三叶虫、菊石、恐龙、剑齿虎……,哪一个物种或物种群(笔者以“物种群”指代“种系”或“支系”,以便于普通大众能够更易理解)不如此呢?达尔文主义坚信,动物不会主动发展出有害的结构,但进化也并不保证有用的结构在改变的环境中继续适应。爱尔兰麋可以说是自己曾经成功的牺牲品,同样的案例在古生物史上可谓是层出不穷,但,都与直生论无缘,都是自然选择的结果。
突变论
前文谈到,孟德尔的工作及其遗传学说在1900年被三位植物学家通过各自对植物杂交的研究而“重新发现”。从此以后,孟德尔遗传学进入了突飞猛进的发展时期。
达尔文的自然选择学说是建立在连续变异的基础上的,认为微小的、连续的变异是进化的基础。孟德尔所研究的生物性状却具有不连续变异的特征,即某一性状的不同表现形式之间存在着显著的差别;另一方面,物种之间也是不连续的。因此自然会使人联想到,通过变异产生新物种的过程是基于不连续的变异造成的,也就是说,进化是由不连续变异引起的。
一时间,孟德尔遗传学把达尔文主义逼上了绝路,在孟德尔遗传学蓬勃发展的形势下,由不连续突变引起进化的突变论思想占了上风。
突变论最初由孟德尔遗传定律的再发现者之一、荷兰植物学家和遗传学家德弗里斯(Hugo Marie de Vrier,1848~1935)提出。德弗里斯早年从事植物生理学研究,在渗透压方面取得卓著成果。1873年他发表的两篇关于攀援植物运动机制的记录,还被达尔文在《攀援植物的运动和习性》一文中加以赞赏。后来他转向遗传学,在进行多年的月见草实验过程中,发现偶尔会突然出现新的类型。他把这种现象称为“突变”,并于1901年提出生物进化起因于骤变的‘突变论’,而后,他以《突变论》(1901~1903,共两卷)、和《物种和变种,它们通过突变而起源》(1905)两部论著,夯实了自己“突变论创始人”的地位。对于自然选择学说,德弗里斯认为自己的突变论只是对其进行了修正而不是完全否定,当新的物种由突变产生之后,自然选择将对其起到保留或是淘汰的作用。有意思的是后来的学者发现,德弗里斯在月见草中发现的“突变”是染色体重组的结果而并不是真正的基因突变。
实际上,“骤变”的思想早有萌芽。在达尔文在世时以及逝世后相当长一段时间,很多生物学家都认为生物进化是以跳跃的方式进行的,新的形态和新的器官源于大的跃变(骤变),而非微小变异在自然选择的作用下缓慢而逐渐地累积。由于缺少足够的过渡型化石而出现化石记录的不连续性,一些古生物学家尤其认可这种观点,即使是自称为“达尔文斗犬”的赫胥黎,也始终不支持达尔文的“渐变”思想。在1860年发表于《威斯敏斯特评论》的文章中,赫胥黎宣称物种转变是以跳跃的方式而非达尔文认为的渐变方式发生的;自然始终做着跳跃,无论过去还是现在。另外,赫胥黎把自然选择一直当作“假说”而不是“理论”。不过,赫胥黎坚持把对达尔文学说的异议限制在学术圈内,对外则统一口径,打击和扑灭所有反达尔文的思想,坚持一切都在宣扬达尔文主义的口号下进行。
回到20世纪初,突变论到了遗传学大师摩尔根(Thomas Hunt Morgan,1866~1945)那里几乎已登峰造极。
摩尔根是美国遗传学家、胚胎学家和进化生物学家,因发现了染色体的遗传机制而创立了染色体遗传理论,并于1933年获得了诺贝尔生理学或医学奖。他及其团队在实验室研究果蝇时,发现了大量的突变。正是对这些突变的研究证明了基因(即孟德尔的“遗传因子”)位于染色体上,从而给魏斯曼的种质连续学说以证明和落实,彻底否定了获得性状遗传。同时,摩尔根学派认为基因突变是产生新性状的唯一动力,突变压力驱动生物向某个特定的方向突变,自然选择仅仅起到淘汰有害突变的消极作用。这种非适应性的突变压力被一些直生论者与“内在动力”联系起来,当成了否定自然选择的武器。
一时间,在突变论学派眼里,达尔文主义同拉马克主义都已是明日黄花。
幸好,达尔文主义不是即将燃尽的恒星,而更像是被月球遮挡了光芒的太阳。后来是如何“月移见日”的呢?“解铃还需系铃人”。请关注下一回:“遗传学——从挑战达尔文到创建现代达尔文主义”。
