若尔盖高原沙化沼泽区植物群落物种组成及其驱动因素
2020-09-17韩大勇杨永兴
韩大勇, 杨永兴
1 伊犁师范大学生物与地理科学学院, 伊宁 835000 2 同济大学环境科学与工程学院长江水环境教育部重点实验室, 污染控制与资源化研究国家重点实验室, 上海 200092 3 浙江双良商达环保有限公司, 杭州 310013
植物群落与环境之间的关系是群落生态学研究的基本内容,其所要回答的基本问题包括:(1)沿环境梯度植物群落种类组成及相关生态过程是如何变化的?(2)何种关键环境因子影响上述变化[1-3]。其中,尤其以植物群落与土壤环境因子的关系广受关注,诸如土壤养分[4-5]、含水量[6-8]和微地形地貌[9-10]等。土壤沙化作为陆地生态系统中最严重的生态环境问题之一,可直接改变土壤物理结构和化学性质,诸如土壤粒度,土壤水分和养分含量等[11-12],由此对植物群落物种组成及其分布产生深远影响。以往研究表明,在土壤沙化过程中,植物群落通常表现出物种多样性降低、生物量下降、群落结构失调[13-16]、牧用价值降低[17]等特征。了解沙化过程中植物群落的变化规律以及如何科学地恢复沙化地植被,是生态学者和生产管理者亟需回答的问题。
若尔盖高原是我国沼泽面积最大、分布最集中、发育最典型的高原沼泽区,为长江、黄河的重要水源涵养地。自20世纪70年代以来,受气候暖干化与人为排水、过度放牧、滥采滥挖等多重干扰,引发严重的局部次生沙化过程[18-22]。据有关数据显示,若尔盖高原沙地面积约从1966年的24.14 km2增加至2006年的96.71 km2,年平均增长率达3.44%,尤以1994—2001年间年均增长率最高,达6.09%[23]。以往就本区沙化成因及治理对策等方面在国内有过较多报道[18-19,24-25],但迄今仍缺少对本区土壤沙化过程中植物群落变化特征的深刻认识,也缺少沙化地植被恢复的群落学依据。本研究是人为干扰下若尔盖高原沼泽植被退化过程与机理系列研究的一部分,以往本课题组曾对该区人为排水和过度放牧干扰下沼泽植被退化过程与机理分别进行过报道[20-22],本文主要研究土壤沙化过程中植物群落分布和种类构成的变化及其与土壤环境因子的关系,为沙化地植被恢复提供参考。
1 研究区概况与研究方法
1.1 研究区自然概况
研究区位于青藏高原东缘若尔盖高原的典型宽谷谷地,海拔3300—3600 m,地理位置101°36′—103°30′E,32°20′—34°00′N,为第四纪新构造抬升运动强烈隆起区的相对沉降区之一[26]。地质构造为刚性较强、稳定的若尔盖地块,岩层为三迭统砂、页岩互层,夹有薄层灰岩[18-19]。地貌类型主要为谷地和缓丘,相对高度50—100 m[26]。年平均气温0.7—1.1℃,最暖月7月平均气温10.9—11.4℃,最冷月1月平均气温-10.5—-7.9℃。年平均降水量650—750 mm,主要集中在6—8月,约占年降水量的1/2。研究区内河流主要为黄河及其支流黑河、白河。较大的湖泊主要有哈丘湖和错拉坚湖。地下水类型主要为第四系松散堆积层孔隙水。植被类型主要为沼泽、草甸和森林,其中沼泽植被主要包括木里苔草(Carexmuliensis)群落、乌拉苔草(C.meyeriana)群落和西藏嵩草(Korbresiatibetica)群落及各种群落复合体。土壤类型主要有沼泽土、泥炭土和亚高山草甸土[18-19]。
1.2 研究地点设置
根据以往在四川省红原县瓦切乡日干乔湿地保护区开展的人为排水和放牧干扰下沼泽植被退化过程与机理研究,本研究选择在同一区域的瓦切乡北部的沙化沼泽区进行。该沙化区为白河(黄河一级支流)下游东侧沙丘向红原县境内的延伸部分,区内原生的木里苔草沼泽已完全消亡,仅谷地中部季节性小河河床残存华扁穗草(Blysmussinocompressus)沼泽化草甸,谷地北缘分布有东西走向流动沙丘。按照从沼泽化草甸至流动沙丘方向建立研究样带,样带宽200—350 m。依据生境类型、土壤类型和植物群落种类构成变化,沿地势逐渐升高方向,进一步划分出6个沙化梯度,相应植被类型为华扁穗草群落、高山嵩草(Kobresiapygmaea)群落、四川嵩草(K.setchwanensis)群落、四川嵩草-黑褐穗苔草(Carexatrofuscasubsp.minor)群落、黑褐穗苔草群落和粗壮嵩草(Kobresiarobusta)群落(表1)。

表1 若尔盖高原沙化沼泽区不同沙化阶段植被及其生境特征
1.3 野外调查
在2010年7—8月开展野外植物群落调查。采用随机+主观取样的原则进行取样,每个沙化阶段内随机设置3个20 m×20 m的样地,样地间隔30—50 m不等,样地内再主观设置5个1 m2样方。样方设置时避开群落边缘和单优无性系斑块,以调查到更多的种类。调查分种盖度、密度和高度后,齐地面割下样方内所有植物,室内105℃烘干称重,计算群落的地上生物量。每个沙化阶段15个样方,总计90个。
土壤取样保证在雨后至少连续3个晴天后进行。每个沙化阶段设置3个土壤采样点,挖土壤探槽,现场采用铝盒和环刀按剖面深度0—20 cm和20—40 cm取样,室内测定土壤湿度、孔隙度和容重等指标。每个沙化梯度6个土壤样品,总计36个。
1.4 数据分析
物种多样性指标分别计算了总种数(样点的总物种数,SRt)、物种丰富度(单位样方内物种数,SRs)和β多样性(Whittaker指数,βw)[27]。各沙化梯度的总种数为3个样点总种数(5个样方)的均值。物种丰富度、Whittaker指数和生物量均为15个样方的均值,在检验方差齐性前提下,用Duncan多重比较分析各多样性指标、生物量在不同沙化梯度间的差异显著性,显著水平P= 0.05。
土壤物理指标计算了土壤湿度(Soil moisture,缩写Soil m.)、容重(Soil volume weight,缩写Soil v.w.)、孔隙度(Soil porosity,缩写Soil p.)。每个沙化阶段的土壤指标均为两个土层的均值。
应用法瑞学派的植被分析方法,依据物种的重要值和存在度,对所有植物进行种类成分划分[28]。将仅在特定群落类型中出现的或仅在该群落类型中重要值最高、而在其他群落中重要值极低的种划为特征种,并联合为特征种组,用以指示特定群落类型和生境条件;将出现于各种群落、存在度较高而重要值相对较低的划为恒有伴生种,这些种对群落无明显选择性,具有较强的生态适应性;将重要值和存在度均很低的划为偶见种,可能为演替残余成分或偶然侵入种。存在度=某种植物出现的样方数/90,划分出5级,I级:0—20%;II级:21%—40%;III级:41%—60%;IV级:61%—80%;V级:81%—100%。
以重要值> 0.1的前40种植物进行RDA排序(典范冗余分析,Canonical Redundancy Analysis),以土壤湿度、容重和孔隙度为环境因子,分析植物群落种类组成与土壤环境条件的相互关系,所有数据均经对数转换(y= log(x+1))以满足线性要求。
文中涉及的公式如下:
重要值=100×(相对盖度+相对高度+相对密度)/3
(1)
Whittaker指数(βw):
(2)
式中SRt和SR分别为总种数和物种丰富度均值。
多重比较应用SPSS 16.0软件完成,RDA排序应用Canoco 4.5 for Windows完成。
2 结果与分析
2.1 植物群落种类组成特征
总计调查到63种维管植物,隶属26科47属,以菊科(Asteraceae)双子叶杂类草最多,为9种,其次禾本科(Poaceae)8种,莎草科(Cyperaceae)6种。
根据重要值和存在度的变化,以及物种对生态条件要求的一致性,将63种植物划分成特征种、恒有伴生种和偶见种。不同沙化阶段植物群落特征种组分述如下:(1)无沙化阶段为华扁穗草种组,包括华扁穗草、木里苔草、溪木贼(Equisetumfluviatile)、竹叶眼子菜(Potamogetonmalaianus)、长茎毛茛(Ranunculuslongicaulis)、西伯利亚蓼(Polygonumsibiricum)6种;(2)极轻度沙化阶段为高山嵩草种组,包括高山嵩草、发草(Deschampsiacaespitosa)、珠芽蓼(Polygonumviviparum)、高山紫菀(Asteralpinus)、条叶银莲花(Anemonetrullifoliavar.linearis)、湿生扁蕾(Gentianopsispaludosa)、条叶垂头菊(Cremanthodiumlineare)、高原毛茛(Ranunculustanguticus)和甘青大戟(Euphorbiamicractina)9种;(3)轻度沙化和中度沙化阶段均为四川嵩草种组,仅四川嵩草和冷地早熟禾(Poacrymophila)2种;(4)重度沙化阶段为黑褐穗苔草种组,包括黑褐穗苔草、二裂委陵菜(Potentillabifurca)、密花香薷(Elsholtziadensa)和垂穗披碱草(Elymusnutans)4种;(5)极度沙化阶段为粗壮嵩草特征种组,包括粗壮嵩草、野胡萝卜(Daucuscarota)、硬秆仲彬草(Kengyiliarigidula)、镰荚棘豆(Oxytropisfalcata)和密毛微孔草(Microulahispidissima)5种。
余下种类中,恒有伴生种10种,在各沙化阶段中均有分布,具有较广的生态适应范围,但在群落中优势度相对较低,主要有湿地繁缕(Stellariauda)、蒲公英属(Taraxacumsp.)、蕨麻(Potentillaanseriana)、草玉梅(Anemonerivularis)、平车前(Plantagodepressa)等。余下27种均为偶见种。
2.2 植物物种多样性变化
物种丰富度和总种数在不同沙化阶段间差异显著(P<0.05),并表现为一致的单峰变化格局,峰值均出现在极轻度沙化阶段,分别为20.47和25.60,最低值均出现在无沙化阶段,分别为5.73和8.60。最高值分别为最低值的3.86倍和2.97倍。Whittaker指数表现为“U”型变化格局,以无沙化和极度沙化阶段的最高,其值分别为0.53和0.54,极轻度沙化和中度沙化阶段的最低,分别为0.25和0.28(图1)。

图1 若尔盖高原沙化沼泽区植物群落物种多样性变化Fig.1 Variation of species diversity of plant community in the desertified swamp area of Zoigê plateau图中字母表示各指数在不同沙化梯度间的Duncan多重比较结果,不同字母代表差异显著,显著水平0.05;ND:无沙化Non-desertification;EMD:极轻度沙化 Extremely mild desertification;SLD:轻度沙化 Slight desertification;MD:中度沙化 Moderate desertification;SD:重度沙化 Severe desertification;ED:极度沙化 Extreme desertification
2.3 植物群落地上部生物量的变化

图2 若尔盖高原沙化沼泽区植物群落地上部生物量(干重)变化Fig.2 Changes of above-ground biomass (dry weight) of plant community in the desertified swamp area of Zoigê plateau图中不同字母代表差异显著, 显著水平0.05;星号表示极端值,圆圈表示异常值,数字代表样本序号
沿土壤沙化梯度,植物群落地上部生物量呈单峰变化格局,最高值出现在中度沙化阶段,均值为165 g/m2,最低值出现在极度沙化阶段,平均值不足50 g/m2,两者相差达3倍以上(图2)。
2.4 植物群落种类组成与土壤因子的RDA排序分析
RDA排序结果较好地体现了植物群落种类构成与土壤因子的关系。与第一轴相关性最高的是土壤湿度,解释物种多度变异总方差的24.8%;与第二轴相关性最高的是土壤容重,方差解释量1.4%。沿第一排序轴,各特征种组对环境表现出明显的选择性,粗壮嵩草种组和黑褐穗苔草种组分布于第一轴最左侧,共同指示沙生生境。其次为四川嵩草种组,指示中生生境;高山嵩草种组与华扁穗草种组分布于第一轴右侧,分别指示湿中生生境和湿生生境,表明土壤水分是影响群落特征种多度变化的主要因素之一。恒有伴生种和偶见种多度随环境梯度未表现出明显的变化,在整个沙化梯度上均有分布,对生境无特殊偏好(图3)。
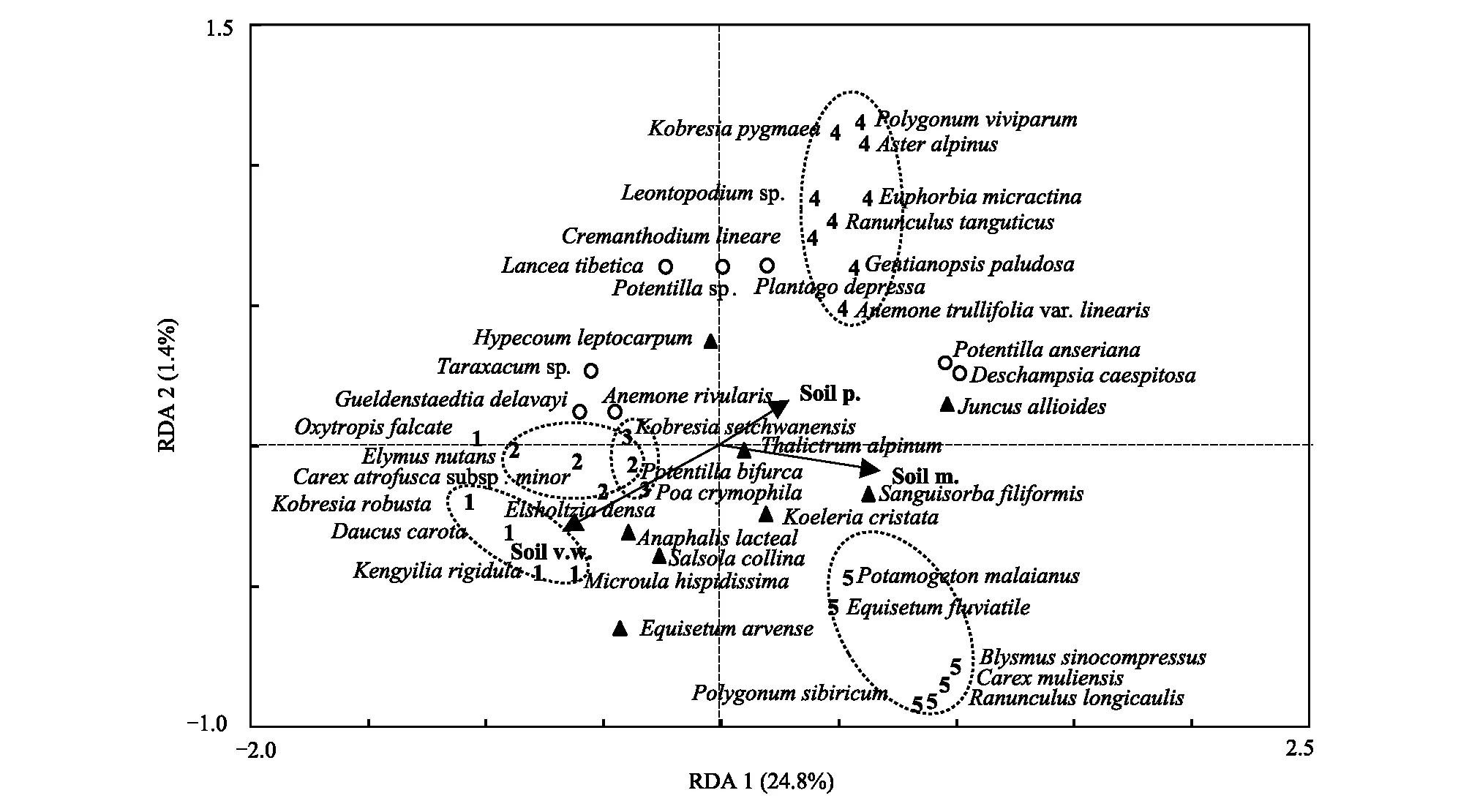
图3 若尔盖高原沙化沼泽区植物群落物种组成与土壤因子RDA分析Fig.3 RDA analysis on species composition of plant community in the desertified swamp area of Zoigê plateau数字和符号代表特征种组,1:粗壮嵩草(Kobresia robusta)种组;2:黑褐穗苔草(Carex atrofusca subsp. minor)种组;3:四川嵩草(Kobresia setchwanensis)种组;4:高山嵩草(Kobresia pygmaea)种组;5:华扁穗草(Blysmus sinocompressus)种组;○:恒有伴生种组;▲:偶见种组;箭头代表环境因子,Soil v.w.:土壤容重 Soil volume weight;Soil m.:土壤湿度 Soil moisture;Soil p.:土壤孔隙度 Soil porosity
3 讨论
3.1 植物群落种类组成变化
植物群落往往具有特定的种类组成和分布范围,并指示不同的生境条件[28]。常学礼和邬建国[13]在科尔沁沙地的一项研究中,提出沙化首先导致群落中特有种的局域灭绝,文中将特有种界定为仅出现于一种生境的种类,本文引入法瑞学派中特征种的概念,界定为出现于一种生境或在该生境中优势度最高的种,并联合为特征种组,尽管两个概念本质不同,但是均可用于指示生境的变化。如本研究中,随着土壤沙化程度增加,特征种组依次出现更替,其顺序为:长根茎型华扁穗草种组→密丛型高山嵩草种组→密丛型四川嵩草种组→长根茎型黑褐穗苔草种组→疏丛型粗壮嵩草种组。表明各群落的特征种组对环境条件变化更为敏感,暗示着这些种类易随环境变化而发生局域灭绝,在这个意义上,与常学礼和邬建国[13]研究结论是一致的,这对于理解沙化过程中植物群落的变化规律以及沙地植被恢复具有重要指导意义。但需要指出群落种组划分具有一定的区域性,因为种的分布区与群落的分布区并不一致,在外推时需谨慎。
3.2 物种多样性变化
植物多样性与生物量是植物群落种类组成变化的结果,同时也反映环境条件的差异[29]。以往本区的同类研究表明在沙化过程中,群落物种多样性和地上生物量均呈单调减少的变化格局[14-16]。但本次研究得到了不同结果,在由沼泽化草甸向流动沙地逆转过程中,物种多样性和地上生物量均呈先增加、后减少的单峰格局。这是生态梯度不同所致,以往研究中沙化梯度是从草甸至流动沙丘,而本研究的沙化梯度是从退化沼泽至半流动沙丘。在疏勒河上游进行的与本研究的生态梯度类似的一项研究发现[30],在湿地向沙地退化过程中,植物群落物种多样性和地上生物量也均表现为单峰格局,可为本文提供佐证。
具体到本研究看,物种丰富度峰值出现在高山嵩草草甸,这与该群落发育于季节性小河高河漫滩生境有关,处于水陆过渡界面,环境相对较好,因此物种多样性较高。但是,高山嵩草植株矮小,群落高度仅10—15 cm,地上生物量并不高,生物量峰值出现在中度沙化的四川嵩草-黑褐穗苔草群落,其原因在于该群落的上层优势种四川嵩草植株较高,呈密丛型生长,丛高20—40 cm,生殖枝则高达60—80 cm,而下层优势种黑褐穗苔草为长根茎型多年生植物,易形成单优群落,盖度较大,由此导致该群落地上生物量出现最大值。β多样性呈“U”型变化格局,最高值出现在粗壮嵩草群落和华扁穗草群落的两个极端生境中,这可能与这两个群落发育的微地貌有关。在华扁穗草群落中,受水位波动和家畜放牧影响,常形成草丘,丘上植物与丘间积水中植物种类差异明显,而粗壮嵩草在流动沙丘上呈丛状分布,草丛内部和外部的种类盖度和组成差异均较大,这两种微地貌类型均增加了群落生境异质性,导致较高的β多样性。
3.3 若尔盖高原沼泽土壤沙化过程分析
来自其他沙化区的研究结果表明,在沙化过程中,土壤湿度是影响植物群落物种组成的关键因素[6,31-33]。本研究也得到类似结果,土壤水分是影响群落分布和物种组成的主要驱动因素之一。在若尔盖高原,沙化地为沼泽极度退化的结果,主要有以下几个方面的证据:(1)若尔盖高原生沼泽多为泥炭沼泽和潜育沼泽,其中发育薄层泥炭土、潜育沼泽土的沼泽分布区容易发生退化;(2)受新构造运动上升影响强烈,野外可见白河河谷下切1 m以上,二者构成沼泽沙化的地质运动背景因素;(3)人为的大规模排水是沼泽沙化的直接诱因[18-19,21]。以往在同一区域的研究发现,在排水沼泽区,相同海拔坡麓分布西藏嵩草和华扁穗草沼泽化草甸,土壤类型为潜育沼泽土[21],而本研究的沙化区同海拔处则退化为四川嵩草中生草甸,土壤类型相应退化为贫瘠的草甸风沙土。上述说明沼泽土壤沙化是从排水干扰进一步退化而来,二者之间具有内在的因果关系,从而形成沼泽—沼泽化草甸—草甸—沙化地的连续退化过程,在此过程中,土壤水分状况经历了强烈的由湿到干的转变,土壤水分状况成为影响植物群落生态分布和种类构成的关键因素之一。为此,退化沼泽生态恢复需要按照沼泽形成、发育、演化规律进行,因土壤沙化导致的若尔盖高原沼泽退化,不能直接从沙地恢复到沼泽,需要经历沙地—沙化地—草甸—沼泽化草甸—沼泽的顺序完成最终的生态恢复。
另外,限于高原恶劣的野外工作条件和环境特殊性,本研究仅现场测定了土壤湿度、容重和孔隙度3个常规指标,其中土壤水分的方差解释量为24.8%,仅作为关键驱动因素之一,还有一些重要的生态因子未纳入观测,诸如土壤有机质含量和粒度组成等,这些指标对土壤持水性能具有重要影响[34-35],需要在后续工作中开展。
3.4 若尔盖高原沼泽区沙化地植被恢复的思考
土壤沙化已成为若尔盖高原的重大环境问题之一。根据野外考查发现,若尔盖地区土壤沙化按照起源可分为3种类型:(1)山坡上沙化为原地起沙;(2)谷地边缘(洼地)无泥炭或有薄层泥炭发育的沼泽是山坡沙化经风力搬运到谷地边缘,掩埋沼泽和沼泽化草甸,属于盛行风向作用下异地起沙过程,前提是陡峻山坡先发生沙化;(3)在没有明显陡峻山坡的区域沼泽化草甸区沙化是原地起沙。在本研究地点,属于第2种情况,其为经风力搬运、堆积而成[18-19],特征为沙粒的分选性较强,粒径小,极易起沙扬尘。近几年采取栽植灌木、撒播多年生牧草、设置生物格栅等多种手段固沙,但恢复效果不理想。硬秆仲彬草、紫羊茅(Festucarubra)等人工撒播的草种盖度很低,仅作为群落伴生种,尽管这些种类适应高原环境,但不适合作为沙地恢复的先锋种。尤其硬秆仲彬草,为高大上繁牧草,不能形成高盖度的群落层片,从而达到减小地表热辐射和水分蒸发的目的。相反,流动沙丘顶部的优势草本植物为粗壮嵩草,该种为疏丛型根茎植物,耐沙埋,常形成基径10—20 cm的分蘖丛,形成突起微地貌,与高原柳镶嵌分布,本研究调查样方即设置在高原柳个体死亡后形成的林隙。大量研究已经证明丛生型灌木或草本植物在荒漠化生态系统中具有特殊生态意义:首先,减少养分随风力水力等外界条件转移流失,积聚枯落物和促进养分归还,改变生物地化循环过程;其次,改变地表物理格局,增加环境异质性,为种子萌发提供安全岛[36];最后,为演替中后期种类侵入创造条件,为人工草地建植奠定基础。本研究中,粗壮嵩草更合适作为沙地恢复的先锋物种,与野胡萝卜、镰荚棘豆、密毛微孔草等一起构成有机的群落种组,这既是植物对流沙环境的一种适应,也是沙地生态系统的一种自我恢复。在沙化地治理中不但要考虑物种生物生态学特性及其与环境的相互作用,还要考虑植物种类组合规律等基本的群落生态学原理,并充分预期在群落演替发育过程中可能出现的问题,诸如群落功能衰退、与野生种的竞争关系等。在科尔沁沙地,建于上世纪50年代的沙地樟子松(Pinussylvestrisvar.mongolica)人工林如今出现大面积衰退[37],就充分说明建立长期、动态群落学思想的重要性。
4 结论
在若尔盖高原沙化沼泽区土壤沙化过程中,植物群落物种丰富度、地上生物量随着沙化程度增加表现出先升高、后降低的单峰变化格局,植物群落表现出明显的退化特征,土壤含水量是这一逆向演替过程的关键驱动因素之一。
沼泽生态系统有其自身的形成、发育、演化规律,由土壤沙化诱发的若尔盖高原沼泽退化,不能直接从沙地恢复到沼泽,需要经历沙地—沙化地—草甸—沼泽化草甸—沼泽的顺序完成最终的生态恢复。
从沙地植被恢复的角度,粗壮嵩草可考虑作为沙地恢复的先锋物种。本研究仅基于植物群落学视角,提出一些恢复对策,沙化地治理是一项系统工程,科学决策更应该综合考虑地质、地貌、气候、水文以及土壤动物、微生物等其他生物类群等多个方面。
