水稻蛋白激酶OsMPK7 负调节水稻的耐旱性
2020-09-15兰金苹郭亚璐王田幸子李莉云窦世娟刘国振
徐 珊, 兰金苹, 郭亚璐, 王田幸子,李莉云, 窦世娟, 刘国振
(1. 河北农业大学 生命科学学院, 河北 保定 071001; 2. 河北北方学院 生命科学研究中心,河北 张家口 075000; 3. 中国农科院 农业基因组研究所,广东 深圳 518116)
促分裂原活化蛋白激酶(Mitogen-activated protein kinase, MAPK)在酵母、真核生物中广泛存在[1]。植物中MAPK 的序列结构高度保守,其作用模式主要是将胞外的刺激信号通过膜受体传递给MAPKKK,然后激活并磷酸化下游的MAPKK,MAPKK 蛋 白 质 中 有 保 守 的S/T-X3~5-S/T 序 列(S 为丝氨酸,T 为苏氨酸,X 为任意氨基酸)。MAPKK 进而激活下游的MAPK[2-3],这样MAPK系统将信号逐级放大并传递给下游的转录因子,如WRKY、bHLH 等,使植物对外界的刺激信号做出应答。
植物MAPK 级联系统在生长发育和逆境应答过程中都发挥重要作用,对生物和非生物逆境胁迫的应答直接决定着作物的产量,其重要性在多种真核细胞信号转导过程中已经被普遍发现和证明[4]。水稻基因组分析表明共有75 个MAPKKK、8 个MAPKK 以及17 个MAPK 基因[5-6]。Ning 等人鉴定到对干旱敏感的DSM1 基因突变,该基因是水稻的MAPKKK,过量表达DSM1 降低水稻的失水速度,提升了苗期水稻对干旱的耐受性,DSM1 的表达受盐胁迫、干旱胁迫和ABA 处理的诱导,但低温处理不影响其表达[7]。水稻OsMPK4 基因的表达受机械损伤和二化螟侵染的诱发,超表达OsMPK4的水稻受二化螟的危害减轻且植株生长迟缓,提示OsMPK4 是二化螟抗性的正调控子和植株生长的负调控子[8]。淹胁迫也能激活MAPK 级联途径,通过磷酸化耐淹基因SUB1A1 提高水稻对淹胁迫的耐受性[9]。本实验室曾报道超表达水稻OsMPK17 蛋白质增强了水稻的耐旱性[10]。水稻MAPK 家族有近百个成员,逐个鉴定水稻MAPK 基因在生物和非生物逆境胁迫应答过程中的功能对水稻基础和应用研究都有重要的意义。
本研究拟开展OsMPK7 的功能研究,首先在前期建立的水稻蛋白质样品资源库(RiceS-A300)的基础上[11],通过免疫印迹(Western blot, WB)分析,发现OsMPK7 蛋白质在旱胁迫下表达下调,具备了开展进一步功能调查的初步数据,为此制备了超表达的载体,转化水稻后鉴定获得了超表达OsMPK7蛋白质的转基因材料,鉴定发现超表达转基因材料在萌发期和幼苗期都降低了对旱胁迫的耐受性。本研究增进了对水稻OsMPK7 基因功能的了解。
1 材料与方法
1.1 材料
1.1.1 植物材料 所用水稻品种为粳稻TP309。
1.1.2 菌株与质粒 克隆用大肠杆菌菌株为DH5a,原核表达载体为pET30a 和pGST,该载体由pGEM改造获得,具体步骤参见文献[12]。超表达质粒构建载体有pUC57-3HA、pEASY-T1(购自上海生工)和pUBI-C4300(由美国UC Davis 植病系Pamela Ronald 教授惠赠)。包含目的基因的质粒为水稻OsMPK7 基因cDNA(AK099472),购自日本农业生物资源研究所水稻基因组资源中心。
1.2 方法
1.2.1 水稻样品总蛋白质提取及WB 分析 水稻样品称重后用锡箔纸包裹,液氮速冻,用研磨仪打碎,按3∶8 样品和蛋白质提取液比例加入相应体积的缓冲液并置于冰上,保持低温。将蛋白质提取缓冲液与水稻样品混合物涡旋4 ~5 次,每次30 s,间隔 2 min。完成涡旋的样品用冷冻离心机离心(4 ℃, 12 000 r/min)20 min,离心回收上清后加入1/4 体积的上样缓冲液(250 mmol/L Tris-HCl pH 6.8、10% SDS、0.5%溴酚蓝、50%甘油和5% β-巯基乙醇)煮沸即得到水稻的总蛋白质[11]。WB 分析方法参见文献[13],主要步骤为:用SDS-PAGE 将总蛋白质样品进行分离,电泳条件为恒压160 V。电泳完成后采用湿转法将蛋白质转移到PVDF膜上,电压100 V,电泳时间1 ~1.5 h。转膜后用30 mL TTBS 缓冲液(0.2 mol/L Tris、1.36 mol/L NaCl、1.5% HCl 和1% Tween-20)配制的 5%的脱脂奶粉封闭,封闭时间为1 ~1.5 h。封闭结束后准备一抗孵育液,在20 mL 的牛奶中加入特定效价比例的一抗(anti-OsMPK7 抗体),室温孵育3 h 或4 ℃过夜。二抗孵育在20 mL 的脱脂奶粉中加入辣根过氧化物酶标记的羊抗鼠二抗。信号检测用MiniChemi610 化学发光成像仪(北京赛智创业),用Image J 软件[14]进行信号采集。以水稻HSP82 的表达丰度作为上样内参[15]。
1.2.2 水稻旱胁迫处理 水稻非生物逆境胁迫处理样品参见文献[11]报道,主要步骤为:水稻种子浸种3 d 露白后在培养箱进行培养(30 ℃,12 h 光/12 h 暗),将水稻种子在纱网上培养,培养5 d 时进行PEG-6000 旱胁迫处理,在0 h、4 h、8 h、12 h、 1 d、2 d、3 d、5 d 和7 d 等时间点取材,对照组用水处理。
1.2.3 超表达转基因载体的构建 目的基因的PCR 引 物 通 过Primer CE[16]软 件 设 计,由 北 京六合华大公司合成。上游引物序列为5’-GC GGTACCAT G C C T G A G G C A A AT G C G-3’,斜体表示Kpn I 识别位点, 下游引物序列为5’-GCGAGCTCGTACATCCTTGAAACAC-3’,斜体表示Sac I 识别位点。PCR 扩增模板为OsMPK7基因的cDNA,将 PCR 产物和 pET30a 载体双酶切,连接产物转化感受态细菌,重组质粒经酶切和测序确认,具体构建过程可参见文献[10]。
1.2.4 农杆菌介导的水稻遗传转化 将pUBIC4300-OsMPK7 质粒DNA 转入水稻TP309,以甘露糖作为筛选压力,遗传转化由武汉伯远公司完 成[17-18]。
1.2.5 水稻种子萌发期旱胁迫处理 参照文献[19]进行PEG-6000(PEG)拟旱处理,取野生型和转基因材料种子各30 粒,用15 ~20 mL 70%的乙醇洗5 min,无菌水洗3 遍,每次5 min,25%次氯酸钠处理30 min,无菌水洗3 遍,每次5 min,进行种子的消毒处理。培养皿里放2 层灭菌滤纸,待萌发的种子用无菌水浸种3 d,至种子露白后进行PEG处理。野生型和转基因材料的种子分为对照组和试验组,对照组用无菌水处理,试验组用20% PEG 处理,在培养箱中培养7 d 后拍照、测量10 株以上水稻的根长和芽长,计算平均值并进行统计分析。
1.2.6 苗期土培干旱胁迫处理 取野生型和转基因材料的种子5 ~8 粒,培养箱里浸种3 d,选取露白且长势均匀的种子进行试验,每盆播种野生型和转基因种子各5 粒,正常培养至苗期并开始停止浇水,让花盆自然干燥。经一定时间的干旱胁迫后,水稻叶片开始干枯、萎蔫,然后再适时恢复浇水,拍照观察记录。
2 结果与分析
2.1 旱胁迫对OsMPK7 蛋白质的丰度的影响
利用本实验室张剑硕等构建的水稻RiceA-S300蛋白质样品资源库[11]。采集水稻苗期经PEG-6000旱胁迫处理0 h、4 h、8 h、12 h、1 d、2 d、3 d、5 d、7 d 等时间点的样品,提取总蛋白质,经SDS-PAGE胶分离后转膜,用OsMPK7 特异抗体进行WB 检测。由图1 可见,对照水处理样品中OsMPK7 的表达丰度基本保持恒定,而旱胁迫处理的样品中OsMPK7 的丰度在4 h 时表现提高,随后持续下降,在1 d 时丰度开始低于0 h 对照时间点,1 d 后样品中OsMPK7 丰度均下降明显。该结果表明,旱胁迫使OsMPK7 蛋白质的表达水平下调,丰度与旱胁迫的负相关提示OsMPK7 基因在旱胁迫应答过程中发挥负调控作用。
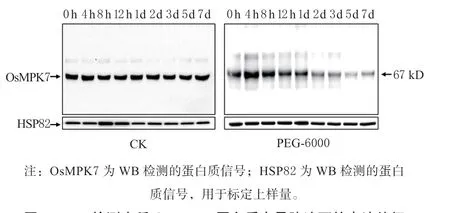
图1 WB 检测水稻OsMPK7 蛋白质在旱胁迫下的表达特征Fig. 1 WB detection of OsMPK7 protein in seedlings under drought stress
2.2 水稻OsMPK7 超表达转基因株系的构建和鉴定
以含有OsMPK7 基因全长cDNA 的质粒为模板进行PCR 扩增,OsMPK7 基因扩增产物大小与理论值相符。将扩增的OsMPK7 片段插入到超表达中间载 体pEASYT1-3HA 中,用Hind III 和Xba I 双 酶切重组质粒,得到4 000 bp 左右的载体片段和1 779 bp 的OsMPK7 片段,酶切产生片段与理论值一致。将重组质粒进行双酶切,回收插入片段,再与转化载体pUBI-C4300 连接,用Kpn I 和Sac I 双酶切验证,确认获得了预期的超表达载体质粒,构建的pUBI-C4300-OsMPK7 超表达载体用于后续的水稻遗传转化。通过农杆菌介导法将构建的OsMPK7 基因超表达质粒转入水稻TP309 中,T0 代获得了22个阳性株系,将阳性植株种子继续播种,在后代筛选鉴定。在T4 代获得了4 个超表达纯合株系(编号依次为A101、A103、A105 和A107)。4 个株系的PCR 和WB 检测结果见图2。由此可见,转基因植株PCR 结果都表现为阳性,而野生型对照植株表现为阴性,WB 检测结果表明转基因株系中OsMPK7蛋白质的丰度明显高于野生型。

图2 超表达OsMPK7 转基因水稻的筛选与鉴定Fig. 2 Screening and identification of OsMPK7 over-express transgenic lines of rice
2.3 OsMPK7 超表达转基因水稻在萌发期旱胁迫下的形态特征
如前所述,苗期样品WB 结果提示OsMPK7 蛋白质在旱胁迫应答中发挥负调控作用。为了进一步验证其功能,用20% PEG 对野生型和超表达转基因材料进行拟旱处理,以水处理的种子作为对照,培养7 d 的结果见图3。在培养过程中发现,拟旱胁迫条件下,超表达转基因种子的萌发较晚,生长缓慢,芽长与根长都低于野生型。其中,A103 转基因株系的生长受旱胁迫影响最大,根与芽的生长受到了严重的抑制。水稻萌发期超表达转基因株系的耐旱能力不强,说明超表达OsMPK7 降低了水稻的耐旱性,而在对照水处理培养条件下,野生型与转基因的种子长势没有明显区别。

图3 超表达OsMPK7 水稻植株萌发期耐旱性鉴定Fig. 3 Drought tolerance assay of OsMPK7 over-expressed rice plants at germination stage
2.4 OsMPK7 超表达转基因水稻苗期耐旱性鉴定
采用同盆栽培的方法对OsMPK7 超表达转基因植株的耐旱性进行分析,每盆栽培野生型与转基因植株共10 棵,正常培养15 d 后停止浇水,7 ~10 d左右水稻叶片表现萎蔫,拍照记录比较,然后再恢复浇水,旱胁迫处理条件下水稻的生长情况见图4。由图可见,在干旱胁迫下,转基因植株的生长均弱于野生型,在离盆拍摄的照片中可清楚地看到明显的萎蔫。恢复浇水后也可看到超表达水稻的生长弱,说明超表达OsMPK7 蛋白质使水稻对旱胁迫更为敏感。
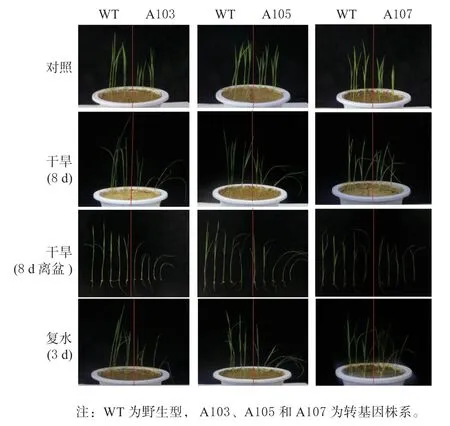
图4 超表达OsMPK7 蛋白质水稻苗期耐旱性鉴定Fig. 4 Characterization of drought tolerance for rice seedlings with over expressed OsMPK7 protein
3 结论与讨论
水稻是重要的粮食作物,水稻的正常生长和产量的获得是抵抗各种逆境胁迫的结果,了解水稻与逆境的互作机制具有重要的理论意义和应用价值。本研究采用基于抗体的水稻蛋白质组学策略[20],通过对逆境胁迫过程中目标蛋白质的表达特征调查获得与功能相关的线索,进而开展转基因验证。
MAPK 级联途径在水稻的正常生长和各种胁迫应答过程中的功能已经有许多报道。敲除水稻的OsMPK15 基因导致病程相关基因(Pathogenesisrelated proteins, PR)的组成型表达、活性氧(ROS)积累的提高以及显著提高对稻瘟病和白叶枯病的抗性,而在OsMPK15 的超表达转基因植株中,PR 基因的表达上调,ROS 的积累提高,对病原物的抗性下降,说明OsMPK15 是抗性的负调控因子[21]。超表达水稻MPKK10.2 通过磷酸化下游的MPK6和MPK3 增强了对细菌条斑病及干旱胁迫的抗性,RNA 干扰植株则降低了抗性,说明MPKK10.2 是一个节点,连同其下游的MPK6 和MPK3 正调控对病原物和干旱胁迫的抗性[22]。水稻OsMPK5 受低温、高盐、干旱和机械损伤等诱导,通过转基因干扰OsMPK5 的表达降低了对干旱、高盐和低温的耐性,支持OsMPK5 在这些逆境胁迫过程中发挥正调控作用[23]。
本研究对OsMPK7 蛋白质在逆境胁迫下的表达特征进行分析,鉴定到OsMPK7 在旱胁迫下表达下调,丰度的负相关提示其发挥负调控作用。为此,构建了OsMPK7 超表达转基因质粒,通过农杆菌介导的策略转化水稻,获得超表达纯合转基因植株。种子萌发期的拟旱试验和苗期的缺水试验结果表明,超表达OsMPK7 降低了水稻的萌发期和苗期对旱胁迫的耐受性。水稻中有近百个MAPK 家族成员,精细地平衡与调控下游基因的表达,保证水稻在面临逆境时做出适当的反应。所以需要对每个MAPK的功能进行多方面的调查,才能逐步描绘出完整的MAPK 调控网络。
