苦豆子种子害虫的斑块边缘效应
2020-09-14张大治吴光胜陈良越马志芳
张大治,吴光胜,陈良越,马志芳
(宁夏大学生命科学学院,银川 750021)
边缘效应最初指生态交错带内的物种数与相邻群落之间的差异,后来不同的学者又根据不同的研究对象、目的和角度,赋予边缘效应不同的概念(田超等,2011)。边缘效应作为一种普遍存在的自然现象,是生态系统(斑块)边缘和生态过渡区所呈现出的生态效应,它在研究群落景观的能量流、物质流和有机体流等生态过程中起着重要的作用,是景观生态学理论探讨的热点之一,也是生态学和保护生物学中非常重要的概念(周宇峰,2007;Carusoetal.,2011)。但由于景观性质、研究目标和斑块形状的变化,边缘效应影响的范围和程度差异较大。核心斑块与周边景观之间的相似性既可以增强边缘效应,也可以减弱边缘效应(陈利顶等,2004),边缘效应是造成不同形状的斑块中生态学差异的最重要的原因(徐化成,1996;王巍巍等,2012),斑块内部和斑块之间的昆虫种群动态将因边缘效应的存在而变得非常复杂(Olson & Andow,2008),甚至景观斑块的边缘部分由于受到外界环境的影响而表现出与其中心部分不同的生态学特征(肖笃宁等,2003)。
在宁夏中东部荒漠沙化区,自然生长的苦豆子呈典型的斑块状分布(贺达汉,2004)。豆荚螟Etiellazinckenella是苦豆子种子的重要害虫,在斑块化的景观格局中,苦豆子豆荚螟趋于均匀分布,这种空间分布型主要是由环境因素引起的(郭晶静等,2012)。斑块面积和斑块隔离度对生物种群有一定的影响,但是斑块面积对苦豆子种子害虫的数量及其危害率的影响不显著(贺泽帅等,2018)。那么在一个相对独立的斑块中,苦豆子种子害虫在斑块内中的空间分布是否存在方位性?是否具有边缘效应?对这些问题的探讨,可以为苦豆子种子害虫野外调查取样、掌握危害发生规律,开展生态防控提供理论依据。
1 研究区域及研究方法
1.1 研究区域概况
调查样地位于宁夏灵武白芨滩国家级自然保护区中北部的宁东镇余家湖,该地域为沙漠低山丘陵地貌,大部分地段为固定半固定沙地,典型的沙生植物有苦豆子Sophoraalopecuroides、柠条Caraganakorshinskii、沙拐枣Calligonummongolicum、甘草Glycyrrhizauralensis、花棒Hedysarumscoparium等。天然野生苦豆子植物在该区域分布相对较多,呈典型的斑块状分布。本实验样地为一个形状较为规则(基本呈圆形)、面积相对较大(半径约30 m)、且较独立(与相邻斑块间隔至少50 m以上)的苦豆子斑块,地理坐标为38°08′13″N,106°60′51″E,斑块周长182 m,面积2 964.23 m2,海拔1 273.2 m,斑块内苦豆子的盖度超过90%,其他植物很少。
1.2 调查方法
于2017年6月中下旬至7月苦豆子结荚中期到豆荚成熟前期进行调查,此时种子害虫处于幼虫期且在豆荚内,便于统计计数。以样地中心为原点,设置半径间隔5 m的五个同心圆,按照东、西、南、北、东北、西北、东南、西南8个方位与同心圆的交叉点为取样样点(1 m×1 m),在每个取样点随机选取5株苦豆子,剪取其全部豆荚分别装入塑封袋带回室内进行测量、剥检。室内测量记录每枚豆荚的长度,在体视显微镜(OLYMPUS SZX7)下剥检每一豆荚,记录荚内豆粒总数、被害豆粒数、种子害虫数等。重复调查2次。
1.3 数据计算方法
采用直接计数法统计荚内的豆荚螟幼虫数,由于个别豆荚螟幼虫啃食完荚内豆粒后会转荚或由于野外采集豆荚在塑封袋带回实验室的途中挤压受热,荚内部分豆荚螟幼虫会咬破豆荚皮而逃离,因此豆荚上的蛀孔数在计数时也视为豆荚螟的幼虫数。凡是荚内存在豆荚螟幼虫或有受害豆粒(通常豆荚上有孔)的就记为受害豆荚(1),否则为不受害豆荚(0)。豆粒受害与否按照豆荚螟幼虫是否啃食进行统计。按以下公式计算:
每荚豆荚螟数=剥检出的总豆荚螟数/总豆荚数;豆荚受害率(%)=受害豆荚数/总豆荚数×100;豆粒受害率(%)=受害豆粒数/总豆粒数×100。
利用EXCEL 2007、DPS(v16.05)统计软件对所得数据进行分析处理。
2 结果与分析
2.1 斑块不同方位和不同距离的豆荚生长情况
一般情况下,景观斑块的边缘部分由于受到外界环境的影响,可能会表现出与其中心部分不同的生态学特征,从而给实验结果带来误差。因此本实验对由斑块中心到边缘不同距离、不同方位苦豆子的豆荚长度、荚内豆粒数进行了统计(表1、表2),豆荚长度为42.53 mm~59.83 mm,平均长度52.45 mm;荚内豆粒数为3~5粒/荚,平均为4.04粒/荚。单因素方差分析(ANOVAs)显示,由中心到边缘不同距离的豆荚长度(F0.05(4,35)=0.7500<2.64,P=0.5647)、每荚豆粒数(F0.05(4,35)=1.3974 < 2.64,P=0.2551)均无显著性差异;8个方位的豆荚平均长度 (F0.05(7,32)=1.0777< 2.31,P=0.4001)、每荚豆粒数(F0.05(7,32)=0.8885 < 2.31,P=0.5268)均无显著差异。因此可以排除苦豆子的自然生长对本实验结果的影响。

表1 各样点苦豆子豆荚长度(mm/荚)Table 1 Pods length of Sophora alopecuroides at different plots(mm/per pod)

表2 各样点苦豆子豆粒数(粒/荚)Table 2 Number of beans of Sophora alopecuroides in per pod (number of beans/per pod)
2.2 斑块不同距离和不同方位的种子害虫数量
苦豆子种子害虫的数量在样地中心区(0 m)平均为0.2602个/荚,然后由5 m时的0.4323个/荚逐渐增加到25 m时0.7328个/荚,呈现出由斑块中心向边缘逐渐增加的趋势(图1),数量差异显著(F0.05(4,35)=1.9748,P<0.05),具有明显的边缘正效应(R2=0.9008)(表3)。不同方位的种子害虫数差异显著性主要体现在北(F0.05(4,20)=3.1171,P<0.05)、南(F0.05(4,20)=2.9249,P<0.05)两个方向,但在东北(F0.05(4,20)=2.6634,P>0.05)、西北(F0.05(4,20)=0.8101,P>0.05)、西(F0.05(4,20)=1.0033,P>0.05)、西南(F0.05(4,20)=1.4137,P>0.05)、东南(F0.05(4,20)=1.8774,P>0.05)、东(F0.05(4,20)=2.8099,P>0.05)方向上的数量并无显著差异(表3)。

表3 各样点苦豆子种子害虫的数量(头/荚)Table 3 Number of seed pest of Sophora alopecuroides in per pod (individual/pod)
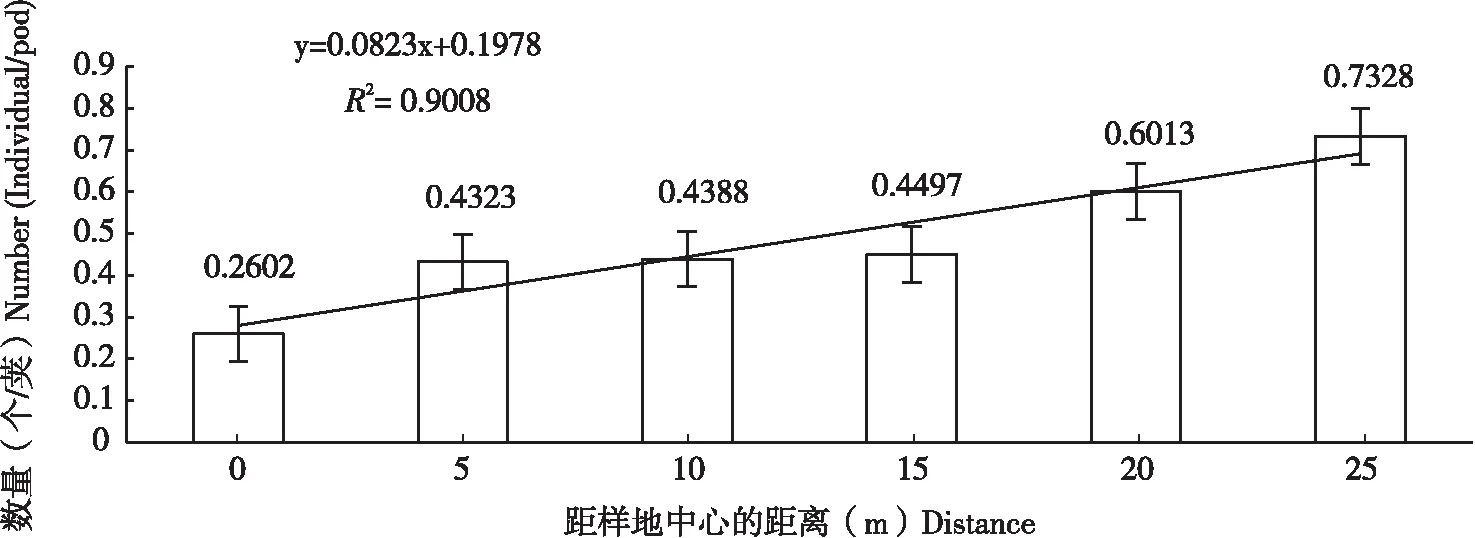
图1 不同距离梯度下的豆荚螟数Fig.1 Number of seed pest of Sophora alopecuroides at different distances
斑块内豆荚螟在空间上主要沿着东南-西北方向呈梭形分布(图2A),且在东南、西北方向趋于边缘分布。

图2 不同方位苦豆子种子害虫的数量(A)及豆荚、豆粒受害率(B)Fig.2 Number and damage ratio of seed pest of Sophora alopecuroides at different directions
2.3 斑块不同距离和不同方位的豆荚、豆粒的受害率
总豆荚受害率、总豆粒受害率由斑块中心向边缘呈逐渐增加的趋势,具有较为明显的边缘正效应(图3)。总体上斑块内不同方位间的总豆荚受害率(F0.05(7,32)=3.3858>2.31,P<0.05)、总豆粒受害率(F0.05(7,32)=2.5088>2.31,P<0.05)差异显著(表4、表5)。豆荚、豆粒的受害率在西北-东南方向明显高于其他方位,呈纺锤状(图2B),这与豆荚螟在空间上的分布完全一致。

图3 距中心不同距离梯度下豆荚受害率及豆粒受害率情况Fig.3 Damage rate of pods and beans at different distances

表5 各样点苦豆子豆粒受害率(%)Table 5 Beans damage rate in different directions
豆荚受害率在东南方向上危害最为严重(63.83%),其次是西北方向(47.87%),受害率最低的为东北方向(23.27%)。差异性检验结果显示东北(F0.05(4,20)=7.4519,P<0.05)、东(F0.05(4,20)=3.1145,P< 0.05)、南(F0.05(4,20)=3.2469,P<0.05)三个方位不同样点间的豆荚受害率有显著差异,而北(F0.05(4,20)=2.1988,P>0.05)、西北(F0.05(4,20)=0.9927,P>0.05)、西(F0.05(4,20)=0.8611,P>0.05)、西南(F0.05(4,20)=1.3666,P>0.05)、东南(F0.05(4,20)=1.6819,P>0.05)方位不同样点间的豆荚受害率差异不显著(表4)。

表4 各样点苦豆子豆荚受害率(%)Table 4 Damage rate of pods in different directions
豆粒受害率在东南方向最高(41.82%),其次是西北方向(30.30%),受害率最低的是东北方向(16.24%)。差异性检验结果显示豆粒受害率在东北(F0.05(4,20)=6.5319,P<0.05)、东(F0.05(4,20)=3.4089,P<0.05)两个方向上具有显著差异,而在北(F0.05(4,20)=1.3099,P>0.05)、西北(F0.05(4,20)=0.7378,P>0.05)、西(F0.05(4,20)=1.4424,P>0.05)、西南(F0.05(4,20)=1.1178,P>0.05)、南(F0.05(4,20)=2.2154,P>0.05)、东南(F0.05(4,20)=0.9472,P>0.05)方向上的受害率并无显著差异(表5)。
3 结论与讨论
边缘效应是推动生物在景观中分布和丰富的最普遍的力量之一(Ewers & Didham,2006;Ribeiroetal.,2016)。物种对边缘的反应通常是高度可变的,并且与环境有关(Ewers & Didham,2006;Hurstetal.,2013)。一般来说,边缘效应是由于生境质量沿着斑块从边缘到内部梯度在资源和条件的质量和数量方面存在差异而形成的(Villasenoretal.,2014)。边缘效应的形成和维持有一定的空间尺度,其中斑块之间的交错形成的边缘效应属于小尺度水平(周婷和彭少麟,2008)。斑块的地形地貌等因素影响斑块内昆虫的空间分布,其中斑块形状对边缘效应的影响十分明显,一般认为圆形或正方形斑块的边缘效应较小(陈利顶等,2004),但是如果核心斑块与周边景观类型有一定的差异,那么斑块的边缘效应会有较大差异(Klinketal.,2015;杨贵军等,2016)。有研究认为在斑块尺度下,斑块周边景观类型的差异使斑块边缘害虫群落多样性高于中间,而天敌群落的多样性低于中间(戈峰等,2004),斑块边缘人为的干扰会使沙生植物害虫在空间分布上受影响较大,虫口密度变小(王文娟,2010),也有研究认为以植物依赖性为介导的边缘效应是非常有限的,植食性昆虫的自身生物学特性起决定性的作用(Altamirano,2016),对一些非专性寄生的植食性昆虫而言,景观周边的资源补偿机制可能会变得比较重要(Summerville & Crist,2010),具有较为明显的边缘效应(张大治,2011;Gonzalezetal.,2015,2017)。本实验苦豆子豆荚长度、每荚豆粒数调查结果显示从斑块中心到边缘各样点也无显著差异,苦豆子生长状况基本一致,盖度也几无变化,加之样地地处自然保护区内,基本不存在人为干扰因素,因此可以排除由于苦豆子斑块边缘区与其内部的生境差异、人为干扰、寄主植物的生长情况等对种子害虫空间分布及边缘效应的影响。豆荚螟的分布及其对种子的危害呈现显著的边缘正效应,与种子害虫的生物学特性、周边景观类型有一定的关系。我们在研究荒漠沙生植物种子害虫的过程中,发现豆荚螟是危害柠条、苦豆子、沙冬青等荒漠沙生豆科植物的种子害虫,成虫具有一定的迁飞能力,可以转移寄主。周边景观基质中的许多豆科植物都可以作为其桥梁寄主,使斑块边缘地带的种群数量高于斑块中心地带,而呈现边缘正效应。但是本实验选择的苦豆子斑块周边只有极少量的柠条,没有其他豆科植物,因此这种边缘效应的出现可能与豆荚螟成虫在苦豆子斑块间飞行产卵的习性有关。
另外,本研究的苦豆子斑块地处流动半流动荒漠丘陵区,南北方向通透,在其东至东南方向、西至西北方向均有稍隆起的小沙丘,豆荚螟成虫在飞翔移动、寻找合适产卵场所(豆荚)的过程中,风的干扰使其向东南、西北方向扩散而择株产卵,致使斑块东南、西北方向苦豆子危害率升高。当然在景观尺度上,一个空间单元是否属于边缘效应影响区,关键在于看它是否受异类景观的影响及其程度,如果受异类景观的影响较大,那么该地区属于边缘效应影响区,否则属于核心地区(田超等,2011),从景观尺度如何来定量评价斑块边缘效应的范围和大小需要进一步探讨。
