不同沙棘品种种子中α-亚麻酸含量差异及相关基因分析
2020-09-09吕中睿魏继华张国昀罗红梅何彩云
张 彤,吕中睿,魏继华,张国昀,罗红梅,何彩云*
(1. 中国林业科学研究院林业研究所,国家林业和草原局林木培育重点实验室,北京 100091;2. 中国林业科学研究院沙漠林业实验中心,内蒙古 磴口 015200)
沙棘(Hippophae rhamnoidesL.)是胡颓子科沙棘属的一种雌雄异株的灌木或小乔木,耐旱性和耐寒性良好。沙棘果实具有独特的药用和营养价值,在中国和俄罗斯有上百年的药用历史[1]。沙棘果实富含维生素、有机酸、氨基酸、脂肪酸、抗氧化剂和类黄酮[2-4],在炎症性疾病、肝脏性疾病、动脉粥样硬化等疾病的治疗以及保护心脏等方面具有显著功效[5]。在沙棘种子含有的众多脂肪酸中,α-亚麻酸(α-Linolenic acid,C18:3N3)的含量十分丰富,且α-亚麻酸对大脑发育、心血管健康、炎症治疗等都具有重要作用[6-9],但α-亚麻酸是人体必需脂肪酸,不能在人体内合成,只能通过食物摄取。豆油、花生油等常见植物食用油中的主要脂肪酸为油酸、亚油酸、棕榈酸和硬脂酸[10-11],α-亚麻酸含量极低,而沙棘中高积累20.3%~36.3%的α-亚麻酸[12],并且沙棘作为耐旱木本植物可以在沙地、荒山、盐碱地等边际土地生长,可避免与大田油料作物争夺土地,是理想的α-亚麻酸供给对象。
目前,油料作物脂肪酸合成途径及调控基因的研究较多,在沙棘的相关研究中多围绕脂肪酸组分及脂肪酸测定方法展开,少有针对沙棘中α-亚麻酸合成机制的研究。本研究选取α-亚麻酸含量存在差异的2个沙棘品种种子,对其含量差异及相关基因进行分析,在基因表达水平上研究沙棘中α-亚麻酸合成及代谢规律,为培育高α-亚麻酸含量的沙棘良种奠定理论基础。
1 实验材料
沙棘种子采自中国林业科学研究院沙漠林业实验中心(内蒙古磴口县),2个沙棘品种——蒙古大果沙棘向阳(Hippophae rhamnoides‘Mongolia’)(XY)和中国沙棘丰宁(H. rhamnoides‘Sinensis’)(FN)的种子分别采自3个不同的发育时期,依次为未成熟期(T1)、半成熟期(T2)和成熟期(T3),在这3个时期沙棘种子的种皮颜色依次呈现为白色、黑白相间斑点色、黑色。因沙棘种皮与果皮的转色期基本一致,为保证时期界定更加精准,同时利用色差计测量沙棘果皮的红绿色指标 a 值和黄蓝指标 b 值,计算色泽比(h=a/b),种皮呈白色且果皮h值介于-0.18~-0.10的界定为T1时期,种皮呈黑白相间斑点色且果皮h值介于0.20~0.28的界定为T2时期,种皮呈黑色且果皮h值介于0.52~0.60的界定为T3时期。沙棘果实采集后快速将种子从果实中剥离,放入液氮中进行速冻,然后转移至-80℃冰箱长久保存,用于下一步分析。
2 实验方法
2.1 脂肪酸提取
将样品冰上解冻,取50 mg样品于2 mL离心管中。加入1 mL的氯仿甲醇(2:1 v/v)溶液,超声30 min。取上清液,加入1%硫酸甲醇溶液2 mL,80℃水浴,甲酯化30 min,随后加入1 mL的正己烷萃取,5 mL的纯水洗涤。吸取上清液500 μL,加入25 μL的水杨酸甲酯(500 mg·L-1)作为内标,混匀加入进样瓶,进样量1 μL,分流比 10∶1,分流进样,进行GC-MS 检测。
2.2 色谱-质谱分析
样品采用 AgilentDB-WAX 毛细管柱(30 m×0.25 mmID×0.25 μm)气相色谱系统进行分离。程序升温:初始温度为 50℃,保持 3 min,随后以10℃·min-1的速度升温至220℃并维持 20 min。载气为氦气,载气流速1.0 mL·min-1。样本队列中每间隔6个实验样本设置1个QC样本,用于检测和评价系统的稳定性及重复性。采用 Agilent7890A/5975C 气-质联用仪进行质谱分析。进样口温度280℃,离子源温度230℃,传输线温度250℃。电子轰击电离(EI)源,SIM 扫描方式,电子能量70 eV。采用 MSDChemStation 软件提取色谱峰面积及保留时间。绘制标准曲线,计算样品中中长链脂肪酸的含量。
2.3 转录组测序分析
按照Qingen(Qingen,DE)试剂盒[13]操作说明对样品中的总RNA进行提取,并去除总RNA中DNA片段的残留。分别使用NanoPhotometer分光光度仪、Qbuit 2.0 荧光计和Qubit RNA 检测试剂盒以及Agilent 2100和RNA Nano 6000试剂盒检测RNA纯度、RNA浓度以及RNA完整性。RNA检测合格后进行文库制备。使用Illumina HiSeq 2000 对RNA文库进行测序,获得原始测序数据。为了保证测序数据质量,对原始数据进行过滤,从而获得高质量数据(Clean data),用于后续生物信息分析。首先使用DESeq R(1.18.0)[14]进行基因表达量分析,筛选差异表达基因,然后通过GOseq R数据包[15]和KOBAS(2.0)[16]分别对差异表达基因进行GO功能注释和KEGG通路分析。
3 结果与分析
3.1 两个沙棘品种种子中脂肪酸组分差异分析
利用气相色谱分析,在2个品种沙棘的种子中共检测到27种脂肪酸(表1),其中,主要脂肪酸为C18:3N3(α-亚麻酸)、C18:2(亚油酸)、C16:0(棕榈酸)、C18:1(油酸)、C18:0(硬脂酸)。随着果实不断成熟,2个品种沙棘种子中,5种主要脂肪酸的含量均逐渐增加,其中,α-亚麻酸和亚油酸含量上升幅度较大。
从图1可以看出:XY与FN的5种主要脂肪酸的含量均在T2时期表现出显著差异。按照脂肪酸含量从大到小排序,在XY中,依次为α-亚麻酸、亚油酸、油酸、棕榈酸、硬脂酸,含量最高的脂肪酸为α-亚麻酸,亚油酸次之;而在FN中,脂肪酸从大到小排序依次为亚油酸、α-亚麻酸、棕榈酸、油酸、硬脂酸,亚油酸大于α-亚麻酸含量(图1)。
在T2时期,2个沙棘品种种子中α-亚麻酸与亚油酸的比值(α-亚麻酸/亚油酸)存在显著差异(P<0.05)。XY中,T2时期3个样本的α-亚麻酸/亚油酸比值分别为1.21、1.17、1.16,而FN分别为0.92、0.75、0.92(表2)。
3.2 转录组数据分析探究沙棘种子α-亚麻酸的合成规律
以XY与FN种子的3个发育时期、每个发育时期3个生物学重复的18个样品为材料,通过RNAseq测序分析共产生140 G clean data,933 483 428条高质量序列(Clean reads)(Q20>97%;Q30>93%),其中,81.5%的序列唯一比对到参考基因组上。对2个品种3个不同发育时期及相同时期不同品种间的表达基因进行差异分析,共鉴定出24 917个差异表达基因(DEGs)。从图2A样本DEGs的主成分分析(PCA)可以看出:2个沙棘品种各个时期生物学重复聚类效果良好,不同时期之间与不同品种之间数据离散效果良好。通过DEGs的分布图(图2B)可以看出:在T1-T3的发育过程中,在T2时期XY与FN之间的DEGs最多。

表1 XY与FN 2个沙棘品种种子在不同发育时期脂肪酸含量的动态变化Table 1 Dynamic changes of fatty acid composition in the seeds of XY and FN at different developmental stages μg·g-1
依据序列同源性将24 917个差异表达基因进行GO功能注释,2 605个基因注释为生物过程(Biological process),554个基因注释为细胞组成(Cellular component),1 354个基因注释为分子功能(Molecular function)。为了进一步了解基因的生物学功能和相互作用,使用KEGG数据库对基因进行通路富集分析。24 917个DEGs注释到122个KEGG通路,其中,有94个DEGs的功能富集于脂肪酸合成(Fatty acid biosynthesis)通路、脂肪酸代谢(Fatty acid metabolism)通路、α-亚麻酸代谢(α-Linolenicacid metabolism)通路和亚油酸代谢(Linoleicacid metabolism)通路。

图1 沙棘XY与FN种子中主要脂肪酸组分的含量变化Fig. 1 Distribution of main fatty acid components in XY and FN seeds of sea buckthorn
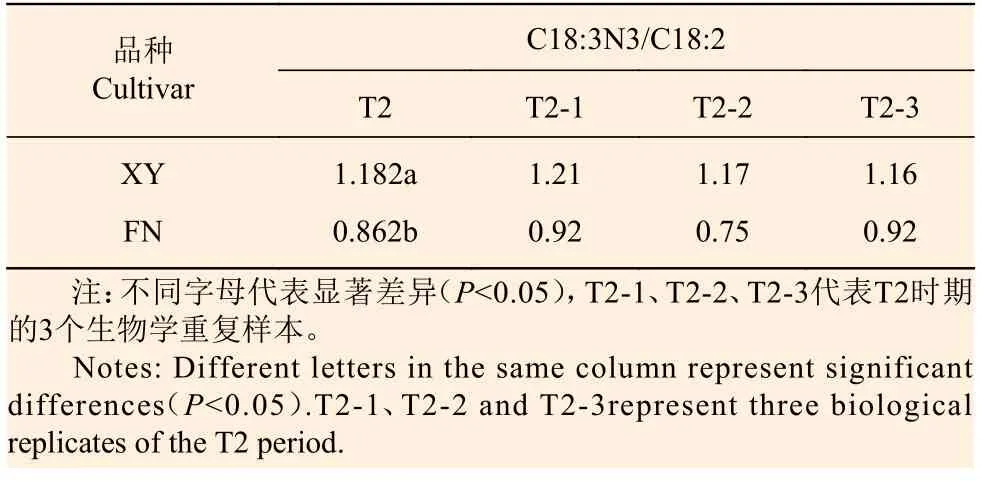
表2 T2时期XY与FN种子中的C18:3N3/C18:2Table 2 C18:3n3 / C18:2 in XY and FN seeds of sea buckthorn at T2
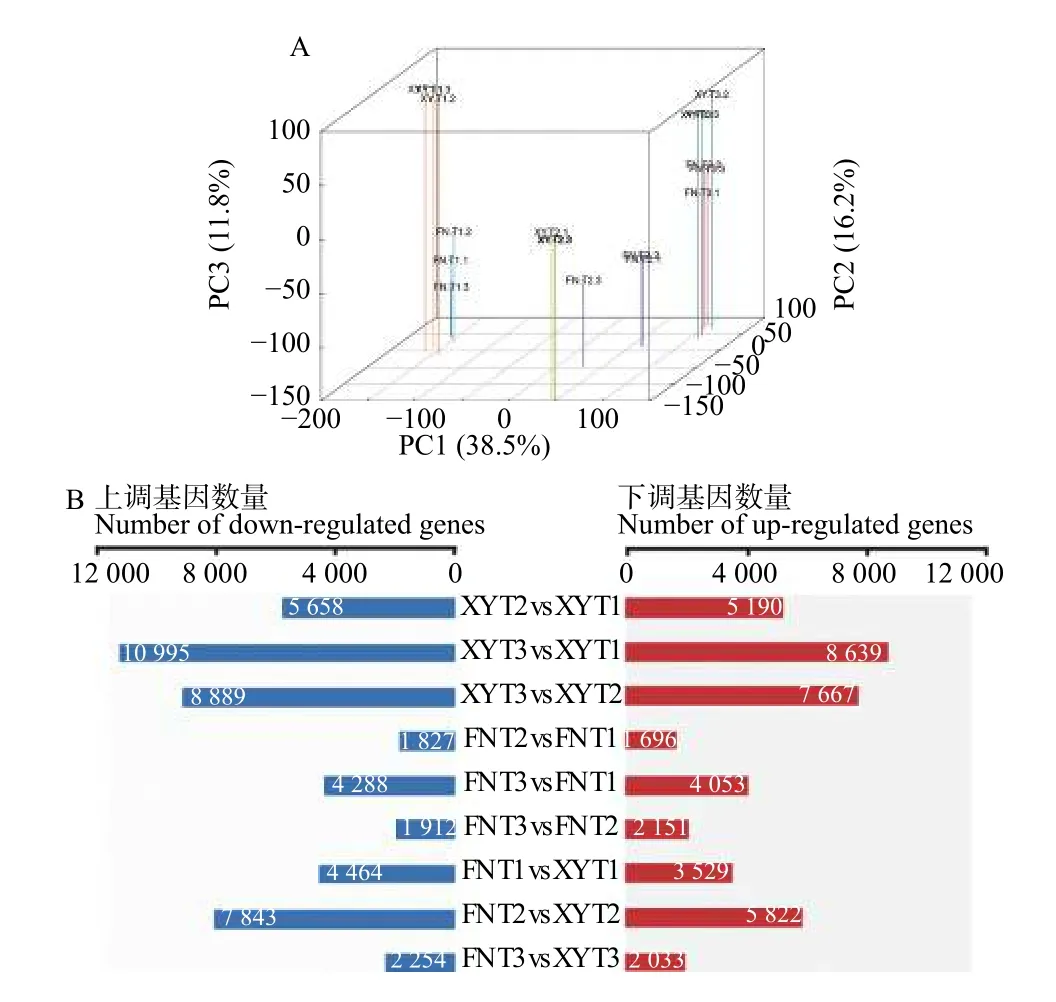
图2 主成分分析与差异表达基因上调和下调分布情况Fig. 2 PCA and up-regulated and down-regulated distribution of DEGs
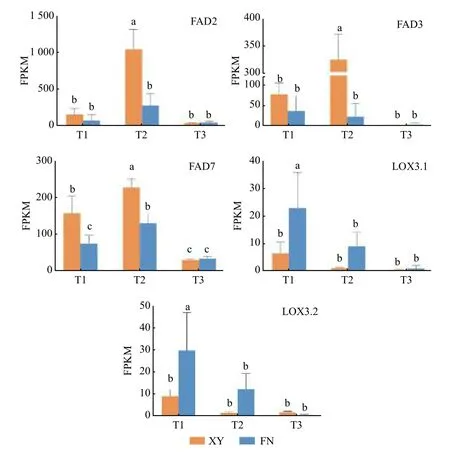
图3 沙棘种子发育过程中与α-亚麻酸合成相关的基因表达模式Fig. 3 Gene expression profiles of Alpha linolenic acid biosynthetic genes during seed development of sea buckthorn
3.3 α-亚麻酸合成与代谢相关基因表达分析
T2时期,基因FAD2在2个沙棘品种中的表达量差异显著(图3),在XY中的表达量显著高于FN,是FN的3.83倍,为α-亚麻酸(C18:3N3)的合成提供充足底物。FAD2基因在XY和FN中表达量均为先上升后下降,在第2时期表达量急剧上升达到峰值,与XY和FN中的亚油酸(C18:2)含量在第二时期显著增加(图1)相一致。
基因FAD3和基因FAD7调控亚麻酸生成α-亚麻酸(图4),在FN中的整体表达水平远低于XY,与XY中α-亚麻酸含量高于FN(图1)的结果相一致。基因FAD3在FN种子中的表达呈下调趋势,在XY种子中的表达水平先急剧上升,在T2时期达到峰值,后在T3时期表达量下调;T1时期,基因FAD3在XY中的表达量是FN 中的2.02倍,T2时期基因FAD3在XY和FN中的表达差异显著,在XY中的表达量是FN中的13.63倍(图3)。基因FAD7在XY和FN中的表达趋势均为先上升后下降,T1和T2时期在XY中的表达量显著高于在FN中的表达量,T1时期在XY中的表达量是FN 中的2.09倍,T2时期在XY中的表达量是FN 中的1.72倍(图3)。
基因LOX调控α-亚麻酸氧化(图4),生成脂肪酸氢过氧化物,是调控α-亚麻酸转化为其他物质的第一步,基因LOX3.1在XY和FN种子的生长发育过程中表达趋势均为逐渐下降,在FN中的表达量高于XY,T1时期基因LOX3.1在FN中的表达量是XY中的3.58倍,T2时期基因LOX3.1在FN中的表达量是XY中的8.02倍(图3)。基因LOX3.2在FN中的表达量同样高于XY,T1时期在FN中的表达量是XY中的3.24倍,T2时期在FN中的表达量是XY中的7.12倍(图3)。基因LOX的高表达不利于α-亚麻酸的积累,与图1中所显示的XY中α-亚麻酸含量高于FN的结果相一致。
4 讨论
沙棘种子中富含不饱和脂肪酸,其中,α-亚麻酸含量一般较高。α-亚麻酸作为人体必需脂肪酸,具有多种有益的生理功能,因此,为了进一步提高沙棘种子中α-亚麻酸含量,培育沙棘新品种和良种,在分子水平上研究沙棘中α-亚麻酸的合成代谢规律至关重要。亚油酸是α-亚麻酸合成的底物,脱饱和生成亚麻酸[17-18]。本研究选取的2个沙棘品种中,α-亚麻酸/亚麻酸的比值存在显著差异,XY种子中α-亚麻酸是含量最高的脂肪酸,亚油酸含量次之,而在FN中正相反,含量最高的脂肪酸是亚油酸,α-亚麻酸含量次之。因此,基于这2个品种存在的差异挖掘沙棘种子中调控亚油酸向α-亚麻酸转化的关键基因以及抑制α-亚麻酸分解的关键基因,探究α-亚麻酸合成代谢规律。
FAD2催化油酸转化为亚油酸[19-21],FAD2基因表达量下降使亚油酸合成受阻,进而影响α-亚麻酸的生成,而FAD2基因的高表达可以为α-亚麻酸的合成提供充足的底物。2个沙棘品种种子中的亚油酸含量在T2时期快速积累,与T2时期基因FAD2的表达水平急剧上升相一致。在XY和FN种子中,T1和T2时期之间的亚油酸含量差异显著。T3时期基因FAD2的表达量降低,亚油酸含量积累速度明显下降。
α-亚麻酸在FAD3和FAD7的催化下由亚油酸脱饱和产生[17-18]。FAD3与FAD7同属于ω-6 型脂肪酸去饱和酶,已有的研究表明,基因FAD3在植物种子α-亚麻酸的合成过程中起到关键作用[22-24]。实验表明,拟南芥的FAD3基因在大豆中异位表达可使大豆中α-亚麻酸含量增加,大豆的FAD3C基因在芝麻中特异性表达同样也可使芝麻中α-亚麻酸的含量显著提高[25]。此外,Wu等将麻风树的JcFAD3基因在拟南芥的种子中特异性表达使拟南芥种子中α-亚麻酸的含量提高了20.50%~24.94%;同时亚油酸的含量相比野生型降低了11.4%~23.5%[26]。在烟草中过表达拟南芥FAD7基因,同样使烟草中α-亚麻酸的含量升高且亚油酸含量下降[23]。
在T1和T2时期,XY中基因FAD3和基因FAD7的表达量远高于FN,尤其在T2时期,基因FAD3在XY和FN中的表达差异显著,在XY中的表达量是FN中的13.63倍,调控大量亚油酸脱饱和生成α-亚麻酸,因此,XY种子中α-亚麻酸的含量大于亚油酸的含量,而FN中亚油酸含量比α-亚麻酸高。这解释了为何基因FAD2在XY种子中的表达量高于FN,但亚油酸含量却是FN种子大于XY种子。
基因LOX是α-亚麻酸代谢过程中的关键基因,是调控α-亚麻酸转化为其他物质的第一步,调控α-亚麻酸氧化,生成脂肪酸氢过氧化物[27-29]。基因LOX3.1和基因LOX3.2在XY和FN发育的T1时期表达量处于3个时期的最大值,抑制α-亚麻酸的积累,协同基因FAD2、FAD3、FAD7在T1时期的低表达水平,导致T1时期α-亚麻酸含量处于低水平,T2时期基因LOX3.1和LOX3.2表达量下调,基因FAD2、FAD3、FAD7表达量急剧上升,因而α-亚麻酸含量在T2时期显著增长。同时,基因LOX3.1和LOX3.2在FN种子中的表达量高于XY种子,抑制FN种子中α-亚麻酸的积累,使FN种子中α-亚麻酸积累量低于XY。
沙棘种子中α-亚麻酸的高积累源于多个基因的协同调控作用,基因FAD2、FAD3、FAD7的高表达协同基因LOX3.1和LOX3.2的低表达使α-亚麻酸的高积累得以实现。
本研究在基因和代谢水平上研究沙棘中α-亚麻酸合成代谢规律,对进一步提高沙棘种子中α-亚麻酸含量、培育沙棘良种具有重要意义。
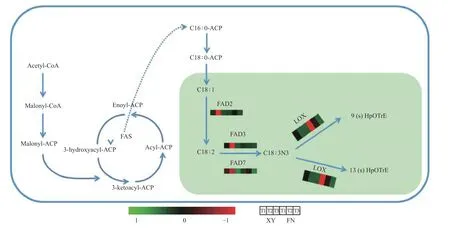
图4 与α-亚麻酸生物合成与代谢相关的途径Fig. 4 Pathway related to the biosynthesis and metabolism of α-linolenic acid
5 结论
通过对2个不同沙棘品种种子中中长链脂肪酸和转录组数据联合分析得出:沙棘种子中α-亚麻酸的高积累源于多个基因的协同调控作用。基因FAD2调控油酸去饱和生成亚油酸,其高表达为α-亚麻酸的合成提供了充足的底物。α-亚麻酸在基因FAD3和基因FAD7的调控下由亚油酸去饱和产生,基因FAD3和基因FAD7的高表达促进亚油酸向α-亚麻酸转化,使α-亚麻酸积累,同时使亚油酸含量下降。沙棘种子中基因LOX3.1和LOX3.2是α-亚麻酸代谢过程中的关键基因,其表达水平下降有利于种子中α-亚麻酸的积累。
