泥鳅选育品系、大鳞副泥鳅选育品系及其杂交子代的生长和形态差异分析
2020-09-07程林慧吴华东王自蕊孙艺文钟建贵彭建华吴春林李福贵
程林慧 ,吴华东,王自蕊,周 颖,孙艺文,钟建贵,彭建华,吴春林,李福贵*
(1.江西农业大学 动物科学技术学院,江西 南昌 330045;2.赣源生态泥鳅养殖场,江西 吉安 331317;3.江西省吉安市渔业局,江西 吉安 343099)
【研究意义】泥鳅(Misgurnus anguillicaudatus)和大鳞副泥鳅(Paramisgurnus dabryanus)分别隶属于鳅科(Cobitidae)、花鳅亚科(Cobitinae)的泥鳅属(Misgurnus)、副泥鳅属(Paramisgurnus),广泛分布于东南亚及我国各大水系江河及湖泊[1-2],是我国重要的小型特种养殖经济鱼类。据FAO 统计,2018 年泥鳅养殖产量多达35 万t[3]。泥鳅具有环境适应能力强,病害少,蛋白质含量高、氨基酸种类丰富、脂肪含量适中,副产物应用价值高等特点[4]。大鳞副泥鳅具有高蛋白、低脂肪、肉质鲜美等特点,其肌肉必需氨基酸含量丰富、比例合理,是理想的动物蛋白源[5]。两种泥鳅外部形态相似度高,主要差异集中体现在颌须、皮褶长和体高3 项指标上[6]。泥鳅生长速度较慢,养殖一年半才达到上市规格,3 年达性成熟,而大鳞副泥鳅生长速度较快,养殖半年可达上市规格,两年达到性成熟[2,7]。近年来,随着市场需求量急剧增加和生态条件的恶化,野生泥鳅和大鳞副泥鳅资源急剧减少。因此,培育养殖性能更加优异的泥鳅品种有着极为重要的生产意义。【本研究切入点】杂交是动植物界普遍存在的自然现象,也是鱼类新品种培育的重要途径之一[8-9]。研究表明,通过种间杂交可以集合双亲的优良性状,丰富杂交后代的遗传结构,产生新的遗传变异,从而获得杂种优势,有效提高子代的生长速率、抗病性、抗逆性等,对培育优良新品种具有积极作用[10]。杂交育种具有周期短、效率高等特点,在生产中应用广泛,一般在子一代即可表现出杂种优势。但不同物种间的远缘杂交,常因生殖隔离而操作复杂、成功率低[9]。杂交育种技术已经广泛应用在水产动物的品种改良及生产实践中,并发挥了巨大的作用。【前人研究进展】顾志敏等[11]通过翘嘴红鲌(Erythroculter ilishaeformis)(♀)和团头鲂(Megalobrama amblycephala)(♂)间的属间杂交成功获得了杂种F1并探讨了杂种F1的性状变异,结果表明杂种F1的框架体型与父母本差异较大,接近于中间型,但受母本影响较多。郭洪洪等[12]获得了三角鲂(Megalobrama terminalis)(♀)×翘嘴鲌(Culter alburnusBasilewsky)(♂)新型鲂鲌杂交F1优良组合,比较分析了杂交F1的生长性能及形态差异,结果显示其具有超双亲生长优势,且可数和可量性状多数表现为中间型。蒋大宁等[7]通过黄河泥鳅(♀)×辽河泥鳅(♂)杂交所获得的F1代在生长性状上表现出长期稳定的生长优势。王林娜等[13]通过云纹石斑鱼(Epinehelus moara)(♀)×鞍带石斑鱼(Epinephelus lanceolatus)(♂)杂交获得的后代“云龙斑”在一些营养指标中明显高于其父母本,显现出一定的杂交优势。关于泥鳅及大鳞副泥鳅杂交,You 等[14]运用微卫星从长江流域多个水系中检测到了泥鳅及大鳞副泥鳅的自然杂交后代。Cui 等[15]对泥鳅和大鳞副泥鳅杂交进行了受精分析研究,表明两者杂交成功率较高。杨兴丽等[16]通过四倍体泥鳅(♂)×二倍体大鳞副泥鳅(♀)杂交所获得的三倍体F1代泥鳅多项营养指标明显高于亲本,表明泥鳅与大鳞副泥鳅杂交种。马来艳等[2]进行了泥鳅与大鳞副泥鳅杂交后代形态学分析,表明正反杂交后代形态均与母本相近,且通过形态参数可以进行初步判别。【拟解决的关键问题】笔者对泥鳅及大鳞副泥鳅分别进行了两代群体选育,均获得了生长性能的提升,并在此基础上构建4 个群体,对两个选育群体间杂种与亲本间的形态差异关系与判别及其生长差异特征进行了分析,以期为泥鳅种质资源保护和新品种培育提供基础资料。
1 材料与方法
1.1 试验材料
本试验所用亲本泥鳅均取自于江西省吉安市新干县赣源生态泥鳅养殖场(北纬27°47′12.96″,东经115°27′35.67″),且所有养殖试验均在该场进行。本试验所用泥鳅(M)与大鳞副泥鳅(P)亲本均取自于经2 代群体选育的亲本群体,分别通过M(♀)×P(♂)与P(♀)×M(♂)获得正交(MP)与反交(PM)2 个杂交F1代群体,通过M(♀)×M(♂)与P(♀)×P(♂)分别获得2个自交F1代群体。
1.2 测定方法
1.2.1 泥鳅的养殖与生长性能的测定 取经2代群体选育的泥鳅(M)与大鳞副泥鳅(P)亲本,随机分成4组,人工繁殖获得M(♀)×P(♂)正交、P(♀)×M(♂)反交F1代群体以及2 个自交F1代群体,每天投喂2 次,从投放苗种开始计算整个饲养时间为期1年。养殖试验期间,每种泥鳅随即抽取60尾,对2、3、4、5、12月龄的4种泥鳅进行体长和体质量的测量。测定前擦干泥鳅体表的水,用电子天平称、直尺分别测量体质量、体长,并记录数据。
1.2.2 形态学数据测量 用电子称、圆规加直尺测量4个群体泥鳅各30尾的传统形态学可量性状(7项)及框架数据(20项),用肉眼观察传统形态学可数性状(4项)。
传统形态学可量形状包括:体质量、体长、全长、体高、体宽、眼径、头长。框架数据包括:D1-2、D1-3、D3-5、D5-7、D7-9、D2-4、D4-6、D6-8、D8-10、D4-1、D4-3、D4-5、D6-3、D6-5、D6-7、D8-5、D8-7、D8-9、D10-7、D10-9,具体测量方法如下图所示(图1)。
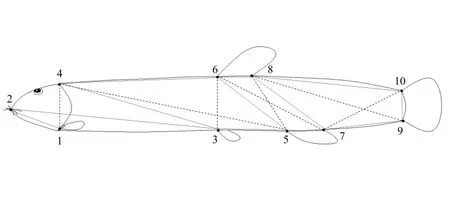
图1 泥鳅框架结构数据测量示意图Fig.1 Sketch map of truss network measures of loach
传统形态学可数性状包括:背鳍鳍条数、胸鳍鳍条数、腹鳍鳍条数、臀鳍鳍条数。
计算出25项形态比例参数:全长/体长、体长/体高、体长/头长、体长/尾柄长、尾柄长/尾柄高、D1-2/全长、D1-3/全长、D3-5/全长、D5-7/全长、D7-9/全长、D2-4/全长、D4-6/全长、D6-8/全长、D8-10/全长、D4-1/全长、D4-3/全长、D4-5/全长、D6-3/全长、D6-5/全长、D6-7/全长、D8-5/全长、D8-7/全长、D8-9/全长、D10-7/全长、D10-9/全长。
计算4个泥鳅群体可量可数性状的杂种指数(hybrid index,HI)[12,17]:HI=100×(Hi-Mi1)/(Mi2-Mi1)。(式中:Hi为杂种平均值;Mi1为母本平均值;Mi2为父本平均值。45<HI<55 属中间性状,HI>100 或HI<0为超亲偏离性状,HI≥55偏父本性状,HI≤45偏母本性状。)
1.3 实验数据处理
生长性能数据采用SPSS 21.0 软件One-way ANOVA 进行方差分析,用LSD 法进行多重比较分析,显著水平为0.05。结果以平均值±标准差的形式表示。形态学数据用Excel 计算各性状比值,分别求出各群体25 项形态比例参数的平均值,对其作聚类分析、判别分析,所采用的聚类方法为Ward 法(度量区间为平方Euclidean距离)。
2 结果与分析
2.1 4个泥鳅群体的生长性能的比较
4个泥鳅群体的不同生长阶段的平均生长量如表1所示。4个泥鳅群体的平均月增质量(g/M)、平均月增长量(cm/M)如表2所示。结合表1表2,结果显示MP 与PM 在生长性能上差异显著(P<0.05),MP 在生长性能上显著高于PM、M,显著低于P;PM在生长性能上显著高于M,显著低于P。
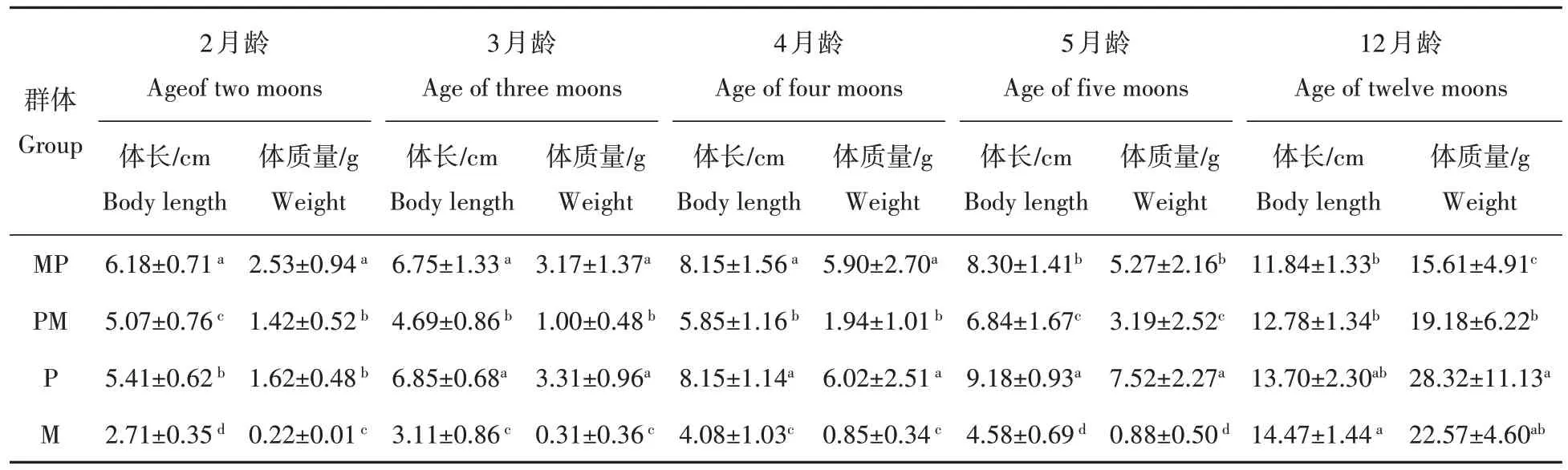
表1 4个泥鳅群体不同生长阶段的平均生长量Tab.1 Average growth of four loach populations at different growth stages
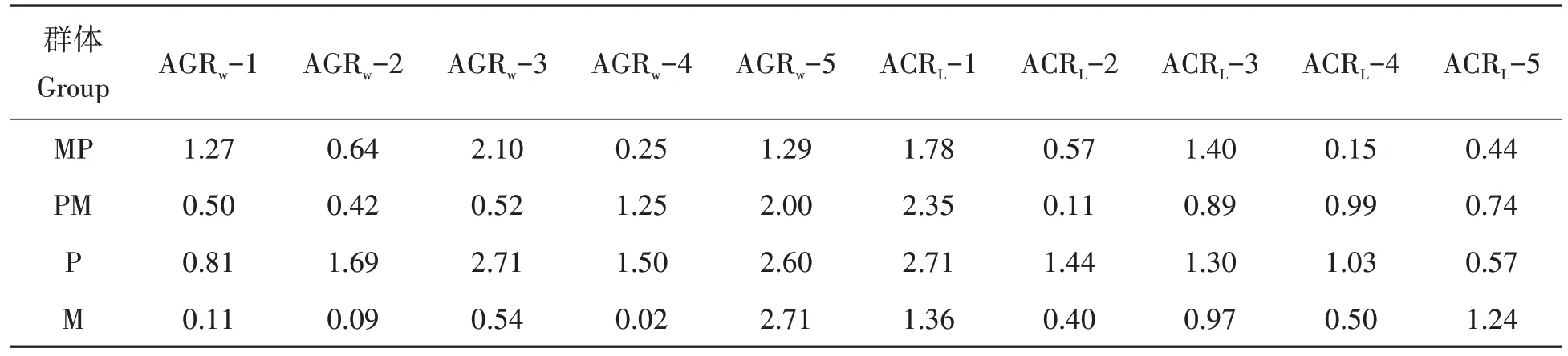
表2 4个泥鳅群体的平均月增质量(g/M)、平均月增长量(cm/M)Tab.2 Average monthly weight gain(g/M)and average monthly growth amount(cm/M)of the four loach populations
2.2 4种泥鳅的形态学数据比较分析
2.2.1 4 个泥鳅群体可量性状形态比例参数 4 个泥鳅群体的外形如图2 所示。通过对4 个泥鳅群体的形态观察,可以发现MP群体同母本M一致,其尾鳍前端皆有一个黑色斑点且体色偏淡;PM群体同母本P一致,尾鳍前端无黑色斑点且体色偏深。

图2 4个泥鳅群体外形图Fig.2 Body shapes of four loach populations
2.2.2 4个泥鳅群体可量性状形态比例参数 4个泥鳅群体的可量性状形态比例参数如表3所示。结果显示,MP 群体的全长/体长、体长/头长、尾柄长/尾柄高、体长/尾柄长、D1-3/全长、D4-3/全长、D4-5/全长7 个比例参数的HI<0,头长/眼间距、D4-6/全长、D10-9/全长3 个比例参数的HI>100,表明这10 个比例参数为超亲偏离性状;D6-5/全长的HI>55,为偏父本P性状;D6-8/全长的45<HI<55,属中间性状;体长/体高、全长/体高、D7-9/全长、D8-10/全长、D4-1/全长、D6-3/全长、D6-7/全长、D8-5/全长、D8-7/全长、D8-9/全长、D10-7/全长11个比例参数的0≤HI<45,为偏母本M性状。PM群体的体长/头长的HI<0,全长/体长、体长/尾柄长、D1-3/全长、D4-3/全长、D4-5/全长、D6-5/全长、D8-7/全长7个比例参数的HI>100,表明这8 个比例参数为超亲偏离性状;体长/体高、全长/体高、尾柄长/尾柄高、D6-8/全长、D8-10/全长、D4-1/全长、D6-3/全长、D6-7/全长、D8-5/全长、D8-9/全长、D10-9/全长11个比例参数的55<HI≤100,为偏父本M 性状;D10-7/全长的45<HI<55,属中间性状;头长/眼间距、D7-9/全长、D4-6/全长3 个比例参数的0≤HI<45,为偏母本P 性状。D1-2/全长、D3-5/全长、D5-7/全长、D2-4/全长4 个比例参数由于P、M 两个亲本数值相同,HI值无效。
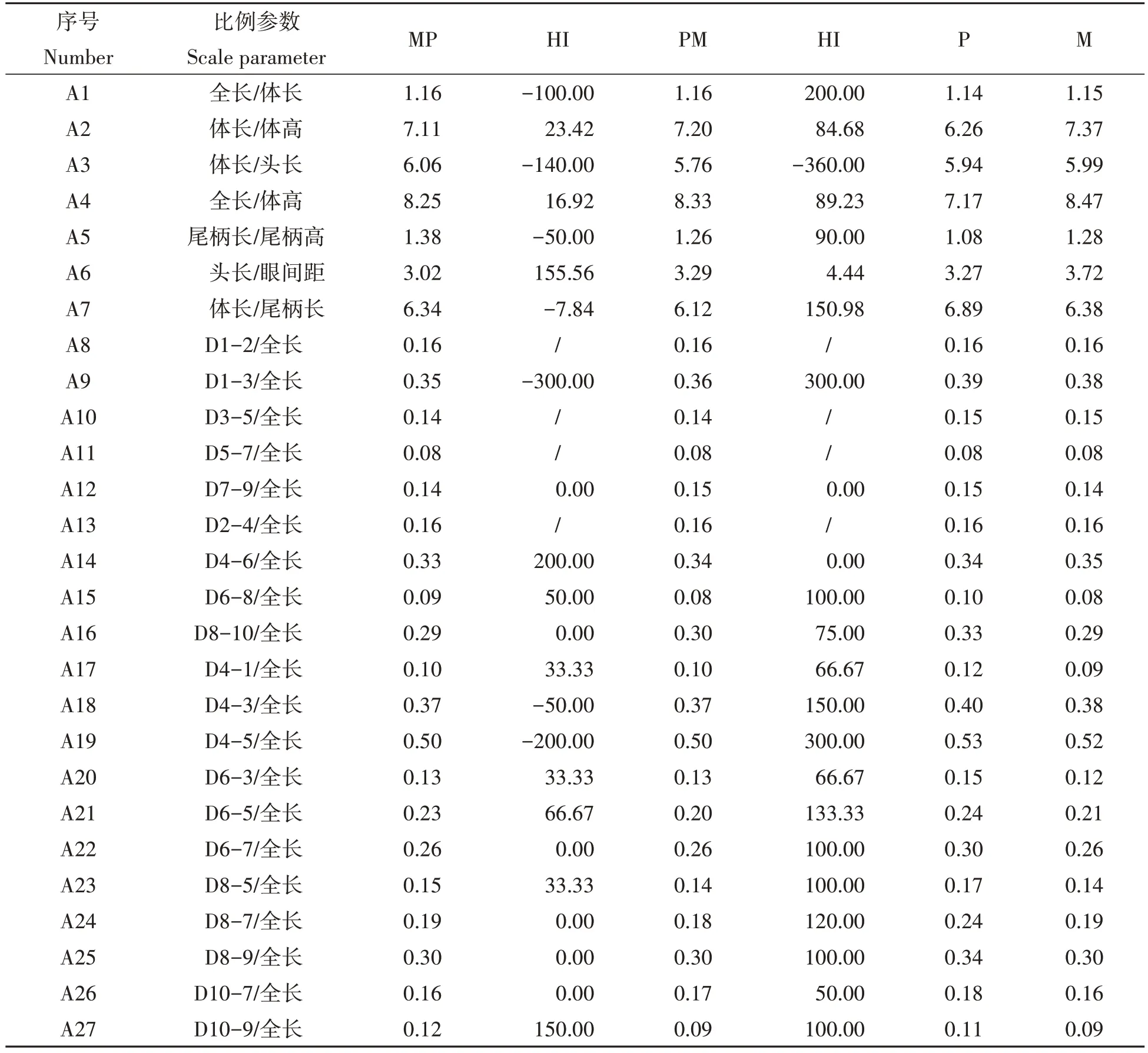
表3 4个泥鳅群体的可量性状形态比例参数Tab.3 Quantitative morphological parameters of four loach populations
2.2.3 可数性状 4个泥鳅群体的可数性状形态比例参数如表4所示。结果显示,MP 群体背鳍、腹鳍鳍条数偏向母本M 群体,胸鳍鳍条数为超亲偏离性状;PM 群体背鳍、腹鳍鳍条数偏向父本M 群体,胸鳍鳍条数偏向母本P群体;由于P、M两个亲本臀鳍条数相同,故其臀鳍条数HI值无效。
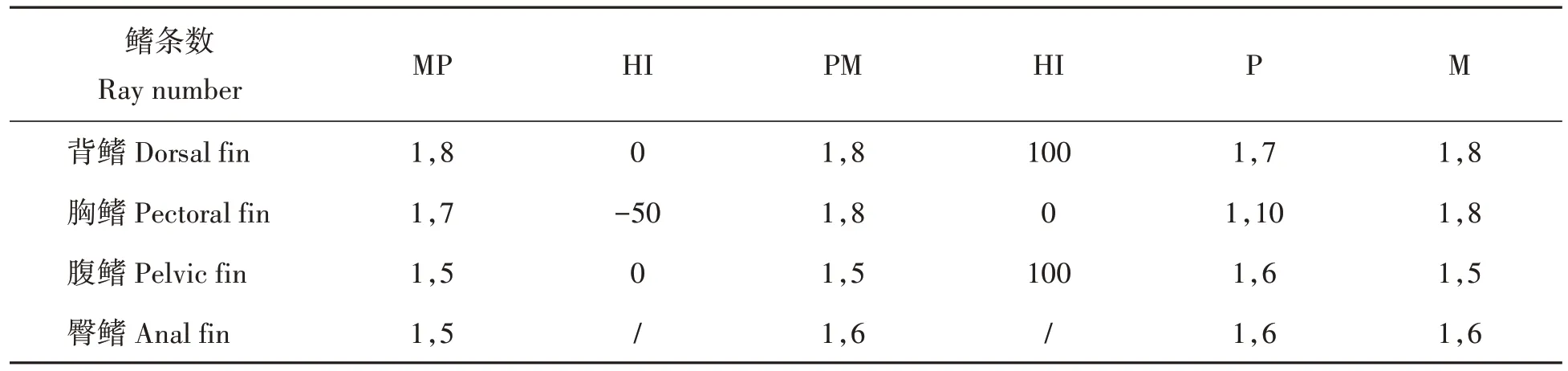
表4 4个泥鳅群体的可数性状形态比例参数Tab.4 Morphological ratio parameters of four loach populations with countable traits
2.2.4 聚类分析 4 个泥鳅群体形态学参数的聚类结果如图3 所示。结果表明,可分为2 个主要分支:MP、PM 和M 聚为一支;P 为另一支。说明MP、PM 两个杂交群体和M 的亲缘关系较近,与P 的亲缘关系较远。
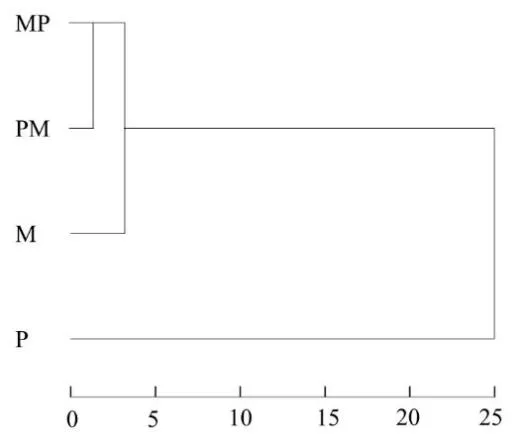
图3 4个泥鳅群体形态聚类图Fig.3 Morphological cluster plot of the four loach groups
2.2.5 主成分分析 4个泥鳅群体形态学参数的主成分分析如表5所示。
结果表明,总方差的贡献率分别是:主成分1 为43.000%,主成分2为23.935%,主成分3为11.871%,累积贡献率为78.806%。即它们包含了总变异的很大一部分,说明可以用几个相互独立的因子来概括4 个泥鳅群体的形态差异。进一步研究分析发现,主成分1对总变异的贡献率高达43.000%,且框架结构特征及体长等21 个指标均可以作为4 个群体形态学差异的判别因子。
判别分析获得全长/体长、D1-2/全长、D1-3/全长、D3-5/全长、D5-7/全长、D7-9/全长、D2-4/全长、D4-6/全长、D6-8/全长、D8-10/全长、D4-1/全长、D4-3/全长、D4-5/全长、D6-3/全长、D6-5/全长、D6-7/全长、D8-5/全长、D8-7/全长、D8-9/全长、D10-7/全长、D10-9/全长等21 个指标贡献最大,分别以A1、A8-A27表示。
当判断一尾未知泥鳅的群体归属时,将所测数据代入方程,函数值最大的可视为其所属群体。构建的判别方程如下所示。
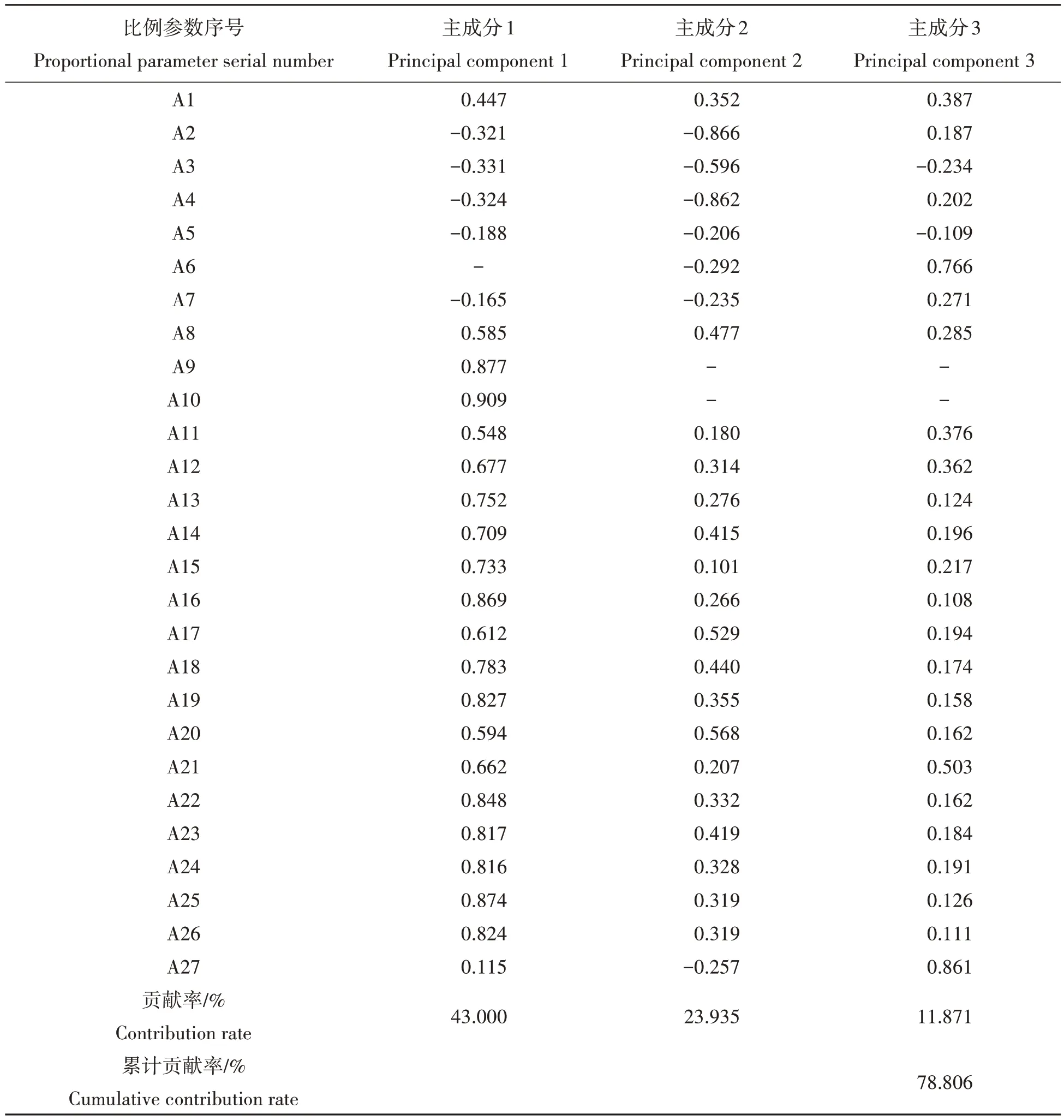
表5 4个泥鳅群体主成分分析中的因子负荷矩阵及主成分对总变异的贡献率Tab.5 Components matrix and contribution of principal components to total variancein PCA analysis for four loach populations
MP:
Y=41 319.124A1-2 510.264A8-341.976A9+573.425A10-1 179.177A11-204.566A12-356.149A13-2 768.239A14-4 164.249A15+163.825A16+2 147.579A17+824.755A18+51.408A19+132.292A20-402.547A21-3 379.714A22-1 231.989A23-528.276A24-1 937.148A25-1 879.491A26+260.810A27-2 2845.430
PM:
Y=41 242.568A1-2 497.251A8-337.371A9+617.835A10-1 266.988A11-178.990A12-421.183A13-2 755.862A14-4 177.782A15+177.691A16+2 222.487A17+800.033A18+51.358A19+103.266A20-422.658A21-3 301.668A22-1 281.460A23-565.836A24-1 941.029A25-1 862.366A26+261.461A27-22 739.406
P:
Y=41 107.434A1-2 572.865A8-335.179A9+511.681A10-1 220.642A11-232.809A12-370.395A13-2 788.333A14-4 133.131A15+175.925A16+2 296.434A17+798.910A18+60.460A19+195.125A20-434.949A21-3 309.319A22-1 225.841A23-486.062A24-1 937.816A25-1 812.345A26+250.248A27-22 621.181
M:
Y=41 312.721A1-2 490.246A8-322.962A9+588.097A10-1 233.939A11-202.314A12-357.556A13-2 738.985A14-4 168.299A15+147.563A16+2 108.515A17+801.962A18+67.930A19+119.406A20-430.780A21-3310.503A22-1 262.934A23-567.895A24-1 935.786A25-1 881.208A26+260.536A27-22 849.493
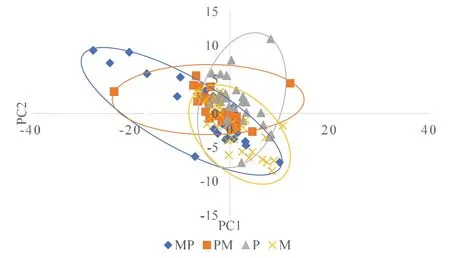
图4 4个泥鳅群体主成分1对主成分2散点图Fig.4 Scatterdiagram for PC1 and PC2 of the four populations
为了验证上述公式的实用性,对所测量的样本按判别公式进行预测分类,将判别结果列于表6。由表6可知,构建方程对各个群体实验鱼的归属进行判别,对MP、PM、P、M 4个泥鳅群体的判别率分别达到83.33%、86.67%、93.33%、83.33%,综合判别率达86.67%。
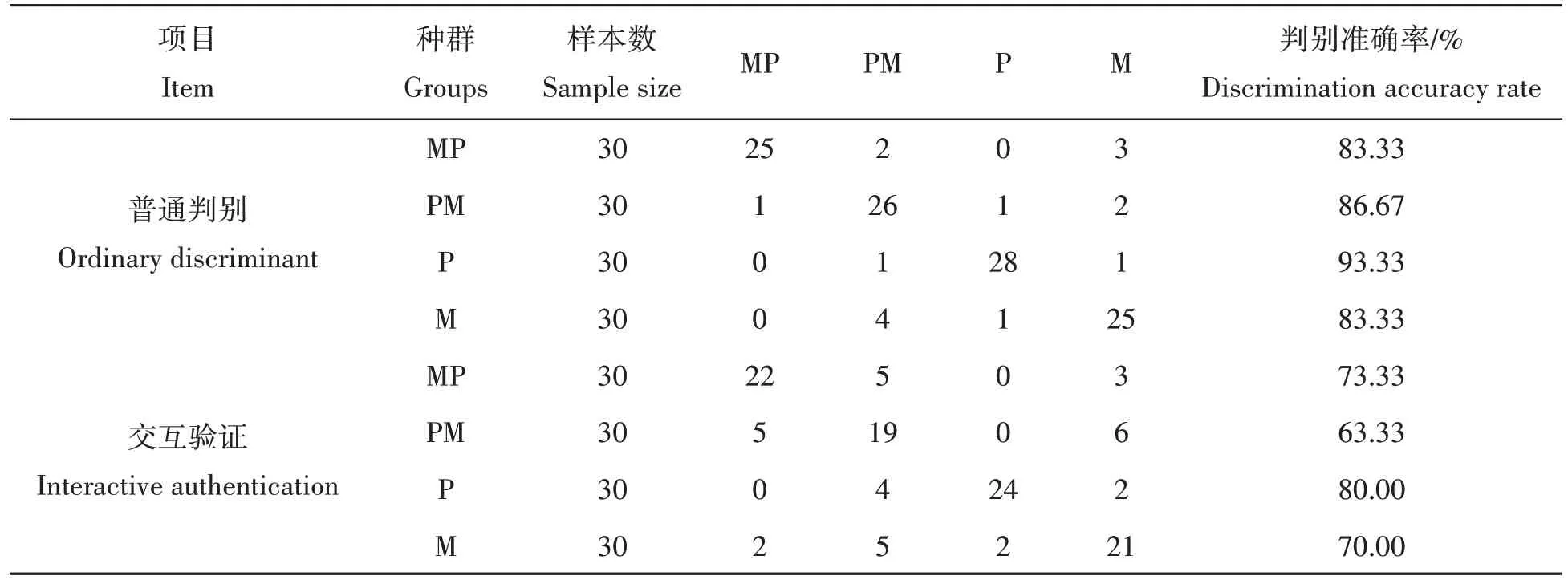
表6 4个泥鳅群体判别分析归类结果Tab.6 Discriminant analysis results of the four loach populations
3 结论与讨论
3.1 生长性能对比
体质量和体长为鱼类最直观的2个生物学体征,是判断鱼类种质质量和养殖效果的标准之一[18]。本研究生长对比试验时间为1 年,泥鳅、大鳞副泥鳅及两杂交子代的2、3、4、5、12 月龄的平均体长、体质量结果见表1,结果显示单位平均体质量由大到小依次是P、MP、PM 和M,MP与PM 在生长性能上差异显著(P<0.05),MP 在生长性能上显著高于PM、M(P<0.05),显著低于P(P<0.05);PM 在生长性能上显著高于M(P<0.05),显著低于P(P<0.05)。4 个泥鳅群体的平均月增质量和平均月增长量,结果表明MP 的0~4月龄的生长速度快于PM、P 和M 3 个群体,5~12 月龄的生长速度慢于其他3 个群体,但差异不显著。为期12月的生长对比试验结果表明杂交后代生长性状上体现出了明显的超单亲(泥鳅)优势,与大鳞副泥鳅存在一定的差距。
顾志敏等[11]、郭洪洪[12]等、李炎璐等[18]、赵博文等[19]、蔡磊等[20]进行了属间、种间、亚种间杂交,杂交获得子代在生长、耐低氧等性状方面有明显的超亲优势,且已经在生产中进行推广应用。蒋大宁等[7]进行了辽河、黄河两个水系的泥鳅品系间杂交,获得体质量性状超双亲的子代。本研究证明泥鳅、大鳞副泥鳅杂交后代MP与PM 在生长1龄阶段前期生长速度均较泥鳅快,体质量增质量比泥鳅高,为越冬准备了更多的能量储备,大大提高了越冬存活率。MP、PM 杂交组合于9~12 龄可以捕获上市,且在该期间错开了泥鳅集中上市高峰期,售卖价格相对较高,养殖效益提升较大。因此,后续研究中继续推进亲本生长性状选育,再进行选育品种间杂交,有望培育有更突出的生长优势性状的杂种泥鳅。
3.2 形态差异比较分析
鱼类的形态学特征与遗传选育[21]、生态环境[22]、投喂方式[23]等都有着一定的联系。当具有不同性状的物种杂交时,杂交种的性状通常表现为两亲本的中间态[24]。据研究报道,大多数鱼类杂交后代的形态性状一般处于中间态且偏向母本的,如唇䱻(Hemibarbus labeo)和花䱻(Hemibarbus maculatus)杂交F1代[25]、青龙斑(斜带石斑鱼Epinephelus coioides♀×鞍带石斑鱼Epinepheluslanceolatus♂)和虎龙斑(棕点石斑鱼Epinephelus fuscoguttatus♀×鞍带石斑鱼♂)杂交后代[26]、鳜(Siniperca chuatsi)(♀)×斑鳜(Siniperca scherzeri)(♂)杂种F1[27]等;但也有一些杂交种形态则偏向父本,如斑鳢(Channa maculata)(♀)×乌鳢(Ophiocephalusargus cantor)(♂)杂种F1[28]、云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交F1等[29];或者不同指标有不同的偏向性,如草鱼(Ctenopharyngodon idellus)、赤眼鳟(Squaliobarbus curriculus)正反杂交F1代[11,30-31]、翘嘴红鲌(Eryth roculter ilishaeformis)(♀)×团头鲂(♂)杂种F1
[11]、赤眼鳟(♀)×鳙(Aristichthys nobilis)(♂)杂种F1[32]等。本研究对4 个群体进行了形态差异分析,在传统可量性状比值方面,HI杂种指数表明,MP 群体有10个超亲偏离性状、1个偏向P、11个偏向M、1个中间性状;PM群体有8个超亲偏离性状、11个偏向M、3个偏向P、1个中间性状,即表明MP群体与PM群体可量性状比值皆偏向于M亲本。在鳍条数目方面,MP群体背鳍、腹鳍数量偏向M,胸鳍数量为超亲偏离性状;PM 群体背鳍、腹鳍数量偏向M,胸鳍数量偏向P;即表明MP 群体与PM 群体鳍条数目皆偏向于M 亲本。在聚类分析方面,聚类图(图3)结果表明MP、PM 2 个杂交群体和M的亲缘关系较近,与P 的亲缘关系较远。因此,MP、PM 杂交种的性状确实表现为两亲本的中间态,但更偏向其中一个亲本M。马来艳等[2]的研究结果显示2种杂交泥鳅形态相近,均与大鳞副泥鳅更相似,而本研究结果显示2 种杂交泥鳅形态相近,均与泥鳅更相似。鉴于地方品种或地理群体间存在形态差异[33-34]、以及家系与群体定向选育导致形态差异[21,35],笔者考虑为不同地方品种的潜在差异以及亲本定向群体选育导致杂交后代形态差异特征结果不同,确切原因有待进一步探究与验证。
判别分析是鱼类鉴定中比较常见的一种方法[36],根据建立的判别方程能进行杂交种与亲本群体间的鉴别。本研究采用了逐步判别分析的方法,按照入选性状的差异大小逐步进行判别,即一边判别,一边引进判别能力最强的变量,最后选取了21 个变量,综合判别率达86.67%,判别效果良好,但存在一定比例的误判。判别公式中贡献较大的变量主要集中在泥鳅背鳍至尾鳍的中间距离,即泥鳅的后半段的长度可作为4个泥鳅群体形态判别的重要指标。
因此,在鱼类形态学研究时可通过可数性状和可量性状的测定与比较的传统方法进行物种鉴定,但对种群或品系鉴定效果有限[37]。本研究涉及4 个群体可以通过外部形态特征的检测及判别方程的建立快速进行区分,但为了提高判别准确度,还需要结合分子标记等手段对杂交种及亲本遗传关系做全面系统的评估。
