艾纳香MYB转录因子家族生物信息学分析
2020-09-02官玲亮查英白琳于福来陈振夏陈晓鹭谢小丽王凯黄梅
官玲亮 查英 白琳 于福来 陈振夏 陈晓鹭 谢小丽 王凯 黄梅

摘 要:MYB轉录因子是植物中庞大的转录因子家族之一,在植物体内有着广泛的生物学功能。基于艾纳香全长转录组数据库,利用DNAMAN 6.0、MEGA 5.05软件以及ProtParam、WebLogo和SOPMA在线网站分析艾纳香MYB转录因子家族蛋白的分类、理化性质、氨基酸序列高级结构、保守结构域、系统进化等。结果表明,挖掘到127个BbMYB基因,预测47条具有MYB转录因子保守结构域的蛋白序列;根据结构域可分为1R-BbMYB和R2R3-MYB 2个亚类,其中R2-BbMYB与R3-BbMYB基序中都含有3个保守的色氨酸,而R1-BbMYB基序只有2个保守的色氨酸;艾纳香MYB家族蛋白均为亲水性蛋白,热稳定性较高且富含碱性氨基酸,大部分蛋白以无规则卷曲为主,其中R2R3-BbMYB3、R2R3-BbMYB4与1R-BbMYB15蛋白是以α-螺旋为主;与拟南芥MYB转录因子家族共同构建的进化树发现,艾纳香MYB家族在进化上包括2个大类,6个亚类,通过艾纳香与拟南芥MYB蛋白序列相邻或是进化关系近可以预测艾纳香MYB基因功能,为进一步对艾纳香MYB家族基因研究及其功能鉴定提供科学依据。
关键词:艾纳香;MYB转录因子;生物信息学中图分类号:S31 文献标识码:A
Bioinformatics Analysis of MYB Transcription Factor Family inBlumea balsamifera
GUAN Lingliang1, ZHA Ying1,2, BAI Lin1,2, YU Fulai1*, CHEN Zhenxia1, CHEN Xiaolu1, XIE Xiaoli1, WANG Kai1, HUANG Mei1
1. Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences / Hainan Provincial Engineering Research Center forBlumea balsamifera, Haikou, Hainan 571101, China; 2. College of Agricultural, Heilongjiang Bayi Agricultural University, Daqing, Heilongjiang 163319, China
Abstract: MYB is one of the largest transcription factors family and has a wide range of biological functions in plants. Based on the full-length transcriptome database ofBlumea balsamifera, DNAMAN 6.0, MEGA 5.05 software and ProtParam, WebLogo and SOPMA online sites were used to analyze and predict the types, physicochemical properties, advanced structure of amino acid sequence, conserved domain and phylogenetic evolution of MYB transcription factor inB. balsamifera, The results showed that 127BbMYBgenes were extracted, and 47 protein sequences with MYB conserved domains were predicted. According to the domain, they could be divided into 1R-BbMYB and R2R3-MYB. Meanwhile, the R2-BbMYB and R3-BbMYB motifs contained three conserved tryptophans, while R1-BbMYB contained only two. Morever, all proteins of MYB family inB. balsamiferawere hydrophilic with high thermal stability and rich in basic amino acids. And most of the proteins were irregular curls except the R2R3-BbMYB3, R2R3-BbMYB4 and 1R-BbMYB15. The MYB family ofB. balsamiferaevolved into two major categories and six subcategories by comparing with the evolutionary tree ofArabidopsis thaliana. It would provide a method for further research on the functional identification of the MYB family inB. balsamiferaby the adjacent or evolutionary relationship with the same function.
Keywords: Blumea balsamifera; MYB transcription factor; bioinformatics
DOI: 10.3969/j.issn.1000-2561.2020.07.004
自1987年转录因子基因首次被报道以来,参与调控植物生长发育、次生代谢以及抗逆等相关的数百种转录因子不断被分离[1]。其中,MYB转录因子是参与植物代谢和调控的一类重要的转录因子[2]。MYB蛋白N端具有由R基序(R1, R2和R3)组成的保守的MYB结构域,且每个R基序含约52个氨基酸,折叠形成螺旋-转角-螺旋(HTH)结构[3]。根据结构域特征将MYB家族分为1R-MYB/MYB-related、R2R3-MYB、R1R2R3- MYB和4R-MYB 4个亚类[4]。其中,1R-MYB/ MYB-related在维持染色体结构的完整性和调节基因转录上起重要作用[5];而R2R3-MYB蛋白参与调节植物的生长发育[6]以及次级代谢[7-8]等。
艾纳香[Blumea balsamifera(L.) DC.]为菊科艾纳香属植物,是我国一种重要的民族药材,主要分布在我国华南、西南地区,在黎族、苗族、壮族等少数民族中有着悠久的药用历史[9]。艾纳香以根、嫩枝、叶入药,用于治疗伤风感冒以及病毒性感染等[10]。研究表明艾纳香还具有抗肿瘤、保肝护肝、抗氧化、抗酪氨酸激酶等药理作用,其中起生物活性的化学成分是黄酮、萜类、噻吩、苯丙素、挥发油等[11]。艾纳香素是艾纳香中一种重要的黄酮类化合物[12],能修复肝细胞药源性损伤而具有保肝作用[13]。夏奇峰等[14]预测了艾纳香中黄酮类物质的代谢途径,表明查耳酮合成酶(CHS)与查耳酮异构酶(HCT)是该路径中的关键酶,可通过调节这些关键酶基因的表达量来提高艾纳香黄酮类成分的含量,从而提高艾纳香的品质。大量研究表明,MYB转录因子参与黄酮类次生代谢产物生物合成的调控,如黄芩[15](Scutellaria baicalensisGeorgi)、拟南芥[16](Ar a b i dopsis thaliana)、苦荞[17] [Fagopyrum tataricum(L.) Gaertn.],但在艾纳香中未见相关报道。
本研究在前期艾纳香全长转录组测序的基础上,利用生物信息学挖掘艾纳香MYB家族转录调控因子,并对其结构域、系统进化、蛋白质理化性质及氨基酸高级结构进行分析,为进一步对艾纳香MYB家族基因的研究及其功能鉴定提供科学依据。
1 材料与方法
1.1序列来源
从艾纳香全长转录组数据库中挖掘到127条BbMYB序列,通过SMART(http://smart.embl.de/)和NCBI BLAST(https://blast.ncbi. nlm.nih.gov/ Blast. cgi)预测得到47条具有MYB转录因子保守结构域的蛋白序列,并通过NCBI的CCD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrp sb.cgi)数据库对蛋白保守结构域进行鉴定。根据牛义岭等[2]對MYB基因家族的结构分类,将47条艾纳香MYB蛋白序列归类到1R-MYB、R2R3-MYB类转录因子中。拟南芥(Arabidopsis thaliana,At)的MYB蛋白序列从TAIR(https:// www.arabidopsis.org/)中下载。
1.2 序列分析
利用ExPASy提供的在线ProtParam软件(http://web.expasy.org/protparam/)对艾纳香MYB转录因子编码蛋白的氨基酸数目、分子量、等电点、脂肪族氨基酸数和蛋白质疏水性等理化性质进行分析。
通过WebLogo网站(http://weblogo.berkeley. edu/)对艾纳香MYB转录因子蛋白进行保守氨基酸基序分析。
利用DNAMAN 6.0软件对47条艾纳香MYB转录因子蛋白和与其结构相似的26条拟南芥MYB转录因子的氨基酸序列进行比对。然后利用MEGA 5.05软件绘制候选艾纳香MYB基因与26个拟南芥MYB基因的进化树,采用邻接法(neighbor-joining method),参数选择P-distance、pairwise deletion和Bootstrap method=1000,其他参数选择默认值。
利用SOPMA(https://npsa-prabi.ibcp.fr/NPSA/ npsa_sopma.html)在线软件进行艾纳香MYB转录因子蛋白二级结构分析,并利用Swiss-Model(https://swissmodel.expasy.org/interactive/)程序对艾纳香MYB转录因子编码蛋白进行三级结构同源建模。
2 结果与分析
2.1 艾纳香MYB转录因子家族基因的挖掘以及蛋白序列的分类
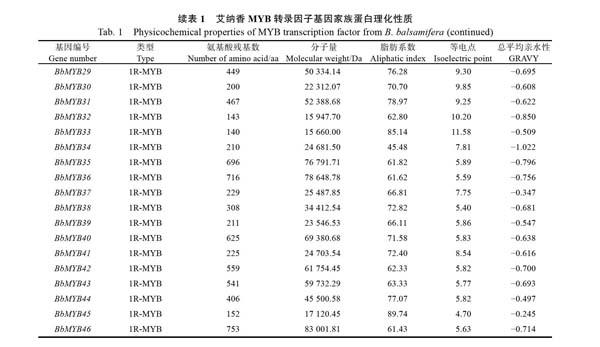
本研究从艾纳香全长转录组数据库中挖掘到127条BbMYB碱基序列,通过SMART和NCBI BLAST预测,去除重复序列及冗余转录本后,最终得到47条具有完整MYB保守结构域的序列,命名为BbMYB1~BbMYB47,其中属于1R-MYB转录因子的碱基序列有32条,长度分布在916~ 3185 bp,属于R2R3-MYB类的碱基序列有15条,长度分布在1615~5173 bp。
2.2 艾纳香MYB转录因子家族蛋白理化性质分析
通过ProtParam在线软件对艾纳香MYB转录因子家族蛋白的理化性质(表1)进行分析可知,艾纳香R2R3-MYB转录因子的氨基酸残基数为158~1307,1R-MYB转录因子的氨基酸残基数为140~799;蛋白等电点在碱性范围内的数量多于酸性范围的数量,表明该家族蛋白富含碱性氨基酸;整个家族蛋白的总平均亲水性GRAVY均为负值,表明艾纳香MYB转录因子家族均为亲水性蛋白,且热稳定性较高。
2.3 艾纳香MYB转录因子家族蛋白保守基序分析
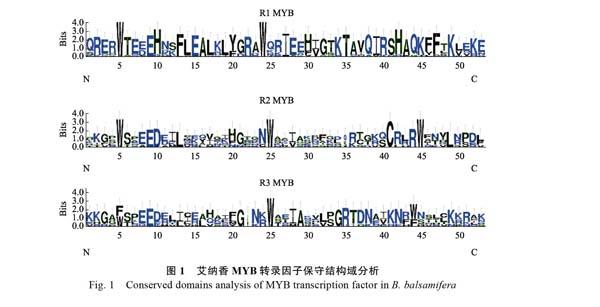
利用DNAMAN 6.0和WebLogo网站对47个艾纳香R2R3-MYB以及1R-MYB类转录因子分别进行多序列比对,保守区域以及2个亚类的基序特征进行分析(图1)。结果表明,R1结构在第5位和第24位有一个保守的色氨酸(W),而
第3个色氨酸被组氨酸(H)、赖氨酸(K)以及精氨酸(R)取代。通过分析R2R3-MYB类蛋白的保守结构域,发现R2、R3结构都包括3个极度保守的色氨酸,且色氨酸之间间隔19~20个氨基酸,符合MYB结构域特征;其次R3基序除第2个色氨酸高度保守外,第1个和第3个色氨酸处还包括苯丙氨酸(F);此外R2、R3结构域中
还包括其他保守的氨基酸残基,如R2结构中的谷氨酸(E)、天冬氨酸(D)、天冬酰胺(N)、半胱氨酸(C)、精氨酸(R)和亮氨酸(L);R3结构中的谷氨酸(E)、脯氨酸(P)、甘氨酸(G)、天冬酰胺(N)、精氨酸(R)和赖氨酸(K),这些氨基酸残基对维持MYB转录因子功能域的HTH结构有关。
2.4 艾纳香MYB转录因子家族系统进化分析
利用MEGA 5.05软件构建艾纳香和拟南芥MYB蛋白家族系统进化树(图2),结果显示艾纳香和拟南芥MYB基因被聚为2个大类(Ⅰ、Ⅱ),其中1R-MYB主要是分布在Ⅰ类,占艾纳香总1R-MYB类转录因子的76.9%,R2R3-MYB主要被聚在第Ⅱ类中,占艾纳香总R2R3-MYB类转录因子的58.3%。第Ⅰ大类又被分为5个亚类,1R-BbMYB24与1R-BbMYB15是最先被分化的,其中1R-BbMYB24与1RMYB-At2G470210相邻,而在1R-BbMYB15基础上进化出相邻的1R-Bb MYB40与1R-BbMYB19,在第5个亚类基础上衍生出R2R3-MYB类。由此可以看出,第Ⅱ类是在第Ⅰ类基础上进化来的,其中1R-BbMYB2与R2R3-BbMYB13相邻且与R2R3MYB-At1 G1 4350、R2R3MYB-At2G02820位于同一进化亚类不同分支;R2R3-BbMYB14与R2R3-BbMYB16相邻且是在R2R3-BbMYB17基础上进化来的;R2R3-BbMYB3、R2R3MYB-At1G09770与R2R3- Bb MYB8、R2R3-AtMYB44分别相邻,表明其进化关系更近。在该进化树中存在着不同结构域但同源性较高的情况,如R2R3-BbMYB1与1R-BbMYB41相邻;同时存在着相同结构域同源性高的情况,如1R-BbMYB42与1R-BbMYB35,1R-BbMYB26与1RMYB-At3G09600分別相邻。总体表明,艾纳香MYB家族蛋白与拟南芥MYB家族蛋白同源性高,在相邻或较近进化关系上的
2.5艾纳香MYB转录因子家族蛋白高级结构分析
2.5.1 蛋白质二级结构分析 利用SOPMA对艾纳香MYB蛋白质进行二级结构预测(图3),结果显示大部分蛋白都以无规则卷曲为主,α-螺旋、β-转角、β-折叠散布于整个蛋白序列;其中R2R3- BbMYB3、R2R3-BbMYB4与1R-BbMYB15蛋白是以α-螺旋为主,β-转角、β-折叠以及无规则卷曲分布在蛋白序列中。值得注意的是,α-螺旋恰
蓝色:α-螺旋;红色:β-折叠;绿色:β-转角;紫色:无规则卷曲。
Blue: α-helix; Red: β-sheet; Green: β-turn;Purple: Random coil.
好是MYB基因保守结构域的位置。因此,推测α-螺旋参与维持MYB转录因子家族的HTH结构。
2.5.2 蛋白质三级结构分析 分别选取R2R3- BbMYB9、R2R3-BbMYB4、1R-BbMYB2、1R- BbMYB15序列进行三级结构同源建模(图4),结果表明,艾纳香MYB家族所有蛋白均具有α-螺旋、无规则卷曲、β-折叠和β-转角,这与二级结构预测相符合。R2R3-BbMYB亚族蛋白三级结构相似性较高,R2、R3结构分别有3个α-螺旋,能明显看到HTH结构,主要是在无规则卷曲长度有所不同。对于1R-BbMYB亚族蛋白,主要在α-螺旋数量上存在差异。
3 讨论
本研究鉴定出32个1R-BbMYB和15个R2R3- BbMYB基因,相对于拟南芥基因组中的118个2R-MYB基因[18]偏少。通过对其翻译的氨基酸序列中保守结构域基序分析,表明R2R3-BbMYB结构域比1R-BbMYB结构域保守。其中,R2-BbMYB與R3-BbMYB基序中都含有3个保守的色氨酸,色氨酸之间的间隔氨基酸数目分别为20、20与20、19。R3-BbMYB基序第1个和第3个色氨酸处还包括疏水氨基酸苯丙氨酸,这与黑果枸杞R3-MYB基序一致[19],表明MYB基因家族N端在不同植物间高度保守,对于维持HTH的构型有着重要的作用[20];其中R2-BbMYB基序中2个色氨酸之间的间隔与杜海[21]描述大豆植物R2R3- MYB一致,说明艾纳香R2-BbMYB基序的色氨酸非常保守;而R3-BbMYB基序中色氨酸比大豆中少1个氨基酸残基,表明艾纳香R2R3-BbMYB类转录因子中的R3-Bb MYB基序发生氨基酸缺失,该区域可能是其植物进化和功能分化的热点区;而R1-BbMYB基序只有2个保守的色氨酸且间隔19个氨基酸,第3个色氨酸被组氨酸(H)、赖氨酸(K)以及精氨酸(R)取代。蛋白理化性质表明艾纳香蛋白富含碱性氨基酸,均属于亲水性蛋白以及热稳定性好。分别对1R-BbMYB和R2R3-BbMYB蛋白进行二级结构预测,分析表明所有蛋白都具α-螺旋、β-转角、β-折叠及无规则卷曲结构,且大部分为无规则卷曲为主,只R2R3- BbMYB3、R2R3-BbMYB4与1R- BbMYB15蛋白是以α-螺旋为主;从2个亚族中选取具有代表性的序列R2R3-BbMYB9、R2R3-BbMYB4、1R- BbMYB2、1R-BbMYB15进行三级结构同源建模,发现R2R3-BbMYB亚族中的蛋白质三级结构相似性较高,在无规则卷曲长度上略有不同;而1R-BbMYB亚族蛋白,主要是在α-螺旋数量上差异较大。通过对艾纳香MYB转录因子的保守结构域基序分析以及氨基酸高级结构的预测,为研究艾纳香属及菊科其他植物的MYB转录因子的结构和功能进化奠定基础。
拟南芥作为一种模式植物,其MYB转录因子的功能研究已经比较清楚,如AtMYB123/ TT2、AtMYB4、AtMYB7和AtMYB24参与调控拟南芥花青素的合成[22-23];研究者认为可以根据进化树中位于同一分枝中拟南芥蛋白功能来预测研究物种蛋白的功能,如在构建的进化树中,与拟南芥R2R3-MYB处于相邻位置上的葡萄、水稻的R2R3-MYB转录因子被认为具有相同或相似功能[19];本研究用所得47条艾纳香MYB类转录因子蛋白与22条拟南芥MYB蛋白作系统进化树,结果显示艾纳香和拟南芥MYB基因被聚为2个大类,其中1R-MYB分布广泛,在Ⅰ类中占艾纳香总1R-MYB类转录因子的76.9%,R2R3-MYB主要被聚在第Ⅱ类中,占艾纳香总R2R3-MYB类转录因子的58.3%;在该进化树中存在着不同结构域但与相同结构域相邻的情况。总体表明,艾纳香MYB家族蛋白与拟南芥MYB家族蛋白同源性高。其中,R2R3-BbMYB8与R2R3-AtMYB44相邻;R2R3-AtMYB11与R2R3- AtMYB12相邻且在R2R3-BbMYB9基础上进化来的。研究表明拟南芥中黄酮类物质主要受AtMYB11、AtMYB12和AtMYB111以及AtMYB75/ PAP1、AtMYB90、AtMYB114调控[24-25]。因此可以推测R2R3-BbMYB9可能具有相似的功能,可能与艾纳香中黄酮类次生代谢产物的生物合成有关,这将为艾纳香MYB转录因子的功能研究提供参考。
本研究参考拟南芥MYB转录因子家族,通过多种生物信息学软件分析艾纳香MYB基因家族的结构特征,为进一步研究艾纳香MYB转录因子的结构和功能奠定基础。
参考文献
[1] 杨致荣, 王兴春, 李西明, 等. 高等植物转录因子的研究进展[J]. 遗传, 2004, 26(3): 403-408.
[2] 牛义岭, 姜秀明, 许向阳. 植物转录因子MYB基因家族的研究进展[J]. 分子植物育种, 2016, 14(8): 2050-2059.
[3] Lipsick J S. One billion years of Myb[J]. Oncogene, 1996, 13(2): 223-235.
[4] 祝志欣, 鲁迎青. 花青素代谢途径与植物颜色变异[J]. 植物学报, 2016, 51(1): 107-119.
- 陈 俊, 王宗阳. 植物MYB类转录因子研究进展[J]. 植物生理与分子生物学学报, 2002, 28(2): 81-88.
- Ambawat S, Sharma P, Yadav N R, et al. MYB transcription factor genes as regulators for plant responses: an overview[J]. Physiology and Molecular Biology of Plants, 2013, 19(3): 307-321.
- Dubos C, Stracke R, Grotewold E, et al. MYB transcription factors in Arabidopsis[J]. Trends in Plant Science, 2010, 15(10): 573-581.
- 沈忠伟, 许 昱, 夏 犇, 等. 植物类黄酮次生代谢生物合成相关转录因子及其在基因工程中的应用[J]. 分子植物育种, 2008, 6(3): 542-548.
- 庞玉新, 谢小丽, 陈振夏, 等. 艾纳香本草考证[J]. 贵州农业科学, 2014, 42(6): 10-13.
- 官玲亮, 庞玉新, 王 丹, 等. 中国民族特色药材艾纳香研究进展[J]. 植物遗传资源学报, 2012, 13(4): 695-698.
- 谢月英, 谢朋飞, 黄宝优, 等. 艾纳香属植物化学成分及药理活性研究概况[J]. 中国现代中药, 2016, 18(8): 1071-1076, 1083.
- 段 震, 周 英. 艾纳香化学成分及药理研究进展[J]. 中华现代临床医学杂志, 2006, 4(21): 1941.
- 许实波, 胡 莹, 林永成, 等. 艾纳香素对护肝及血小板聚集的作用[J]. 中山大学学报论丛, 1994(6): 48-53.
[14] 夏奇峰, 赵 致, 刘红昌, 等. 艾纳香黄酮类物质生物合成途径分析[J]. 中国中药杂志, 2016, 41(19): 3630-3636.
[15] 伍 翀. 黃芩MYB转录因子功能初步研究[D]. 武汉: 武汉工业学院, 2012.
[16] Borevitz J O, Xia Y J, Blount J,et al. Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis[J]. The Plant Cell, 2000, 12(12): 2383-2394.
[17] 赵海霞, 吴小峰, 白悦辰, 等. 苦荞芽期黄酮合成关键酶和MYB转录因子基因的表达分析[J]. 农业生物技术学报, 2012, 20(2): 121-128.
[18] Stracke R, Werber M, Weisshaar B. The R2R3-MYB gene family inArabidopsis thaliana[J]. Current Opinion in Plant Biology, 2001, 4(5): 447-456.
[19] 严 莉, 王翠平, 陈建伟, 等. 基于转录组信息的黑果枸杞MYB转录因子家族分析[J]. 中国农业科学, 2017, 50(20): 3991-4002.
[20] Jin H, Martin C. Multifunctionality and diversity within the plant MYB-gene family[J]. Plant Molecular Biology, 1999, 41(5): 577-585.
[21] 杜 海. 植物MYB转录因子家族的分子进化机制及调控类黄酮生物合成MYB基因的鉴定[D]. 雅安: 四川农业大学, 2013.
[22] Lepiniec L, Debeaujon I, Routaboul J M,et al. Genetics and biochemistry of seed flavonoids[J]. Annual Review of Plant Biology, 2006, 57(1): 405-430.
[23] Song S S, Qi T C, Huang H,et al. The Jasmonate-ZIM domain proteins interact with the R2R3-MYB transcription factors MYB21 and MYB24 to affect Jasmonate-regulated stamen development inArabidopsis[J]. The Plant Cell, 2011, 23(3): 1000-1013.
[24] Stracke R, Ishihara H, Huep G,et al. Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of theArabidopsisthalianaseedling[J]. The Plant Journal, 2007, 50(4): 660- 677.
[25] Gonzalez A, Zhao M, Leavitt J M,et al. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex inArabidopsisseedlings[J]. The Plant Journal, 2008, 53(5): 814-827.
