安徽萧县金寨遗址(2017)植物遗存分析
2020-09-02段绮梦张小雷靳桂云
杨 凡 段绮梦 张小雷 靳桂云
(1.山东大学历史文化学院 山东济南 250100;2、4.山东大学文化遗产研究院 山东青岛 266237;3.安徽省文物考古研究所 安徽合肥 230601)
内容提要:金寨遗址位于淮河中游的皖北地区,是大汶口文化中晚期到龙山文化时期的一处中心性聚落。该遗址2017年的浮选和植硅体样品分析结果显示,在大汶口文化晚期到龙山文化时期,粟是金寨聚落主要的农作物,其次是水稻,这与淮河中游其他遗址的情况一致。淮河流域从东南向西北稻的比重逐渐降低,可能是受到气候条件限制或稻作农业传播速率的影响。植物遗存的空间分布显示金寨聚落的水稻在东区加工、北区消费,粟黍的加工和消费都集中在东区。造成这种差异的原因可能是不同区域的居民具有不同的食物偏好或等级差别。此次发现的龙山文化时期的植物遗存较少,可能和该时期聚落居住区的迁移有关。
淮河中游不仅是中国东部南北地理、气候过渡带,同时还是史前中国南北、东西文化交流、碰撞与融合的核心区域,中原、海岱、江汉、环太湖四大文化区之间的传播、扩散和交汇都是以淮河流域为中介而实现的[1]。淮河流域的新石器时代和青铜时代文化在中国古代文明的发展进程中独具特色,自成体系,“淮系文化”在某种意义上与黄河和长江流域文化并列,共同创造了中国古代文明[2]。“苏鲁豫皖文化区”可能存在一个或多个重要的原始文化[3],而农业是文明发生与发展的主要动因之一[4]。淮河中游处于中国南方稻作农业区与北方旱作农业区的过渡地带。根据现有研究资料,淮河上游地区在裴李岗文化中期阶段就出现了旱稻混作的农业模式[5];淮河中游在裴李岗时代晚期也出现了旱稻混作农业[6],旱稻混作是该地区农业的主要特征,但不同遗址间稻作和旱作的比例差别较大,影响农作物选择的因素也需要进一步探讨。金寨遗址是淮河中游一处中心性的聚落遗址,其年代从大汶口文化中晚期延续到龙山文化中期,这个时期社会分层持续发展,社会基本结构从家族转变为个体家庭[7]。对该遗址的植物遗存进行分析为我们探讨淮河中游地区农作物结构的变迁以及农业和文化交流、社会分层等问题的互动关系等提供了科学依据。
一、遗址简介
金寨遗址位于安徽省宿州市萧县庄里乡尠沟行政村金寨自然村旁,现代村落位于遗址中间。向南延伸到宿州市埇桥区夹沟镇草厂村,西北距萧县30千米,东北距徐州35千米。遗址坐落于皖东北丘陵之中的山前平原,西、南、东三面环山,西、北、东南临小河;文化遗存的年代从大汶口文化中晚期到龙山文化中期,距今约5200—4300年;总面积约50万平方米,是徐淮地区面积最大的新石器时代遗址。2016年9月—2017年1月和2017年10月—2018年10月,安徽省文物考古研究所对该遗址进行了两次发掘。遗址东区主体为一集中墓地,也发现少量房址和灰坑;北区主体为一片排房基址,堆积丰富,从大汶口文化中期延续到晚期。大汶口文化中期时,遗址西区和北区为居住区,东区为墓葬区;大汶口文化晚期时,西区和北区主体是居住区,并有少量墓葬,东区有少量墓葬;到了龙山文化时期,整个聚落向东北方向延伸[8]。
二、材料与方法
本次分析的浮选土样和植硅体土样都是在2017年发掘过程中采用针对性采样法采集[9],性质明确的遗迹作为主要采样单位,分层的单位则分层采集。浮选土样包括大汶口文化中期土样1份、大汶口文化晚期土样31份(30份取自灰坑、1份取自灰沟)、龙山文化时期土样18份(17份取自灰坑、1份取自地层),合计50份,土量729升,平均每份约14.58升。植硅体土样有大汶口文化晚期土样20份(都取自灰坑)和龙山文化时期土样13份(12份取自灰坑、1份取自地层)。
浮选使用小水桶浮选法在发掘现场完成。轻浮和重浮分别使用80目和20目的分样筛进行收集。样品阴干之后,在山东大学植物考古实验室进行分类、鉴定和拍照。样品鉴定使用尼康(SMZ-645)体视显微镜,拍照使用尼康数字相机系统(DS-5M-L1),炭化植物遗存的鉴定参考了实验室所收集的现代植物标本、炭化植物标本、发表的文章中的种子照片和相关植物图谱[10]。鉴定过程中,对于破损严重、丢失了可鉴定部位或者已经成为碎块的植物遗存归入不可鉴定类,在分析讨论时不参与计数。
所有植硅体土样在山东大学植物考古实验室进行处理和分析。植硅体提取采用重液提取法[11],把土壤样品烘干后称取2克进行提取,具体操作为:(1)将土样放入试管中加入30%双氧水(H2O2),充分反应,去除有机质;(2)加入浓度为10%的稀盐酸,用水浴锅加热30分钟,去除样品中的有机质和钙、铁等矿物质;(3)用蒸馏水离心清洗3—5次,将盐酸清洗干净;(4)加入碘化镉重液,使植硅体浮于重液表层并收集,加蒸馏水洗去重液并烘干;(5)用中性树胶制成固定玻片。
植硅体鉴定和统计所用的显微镜为Nikon公司生产的型号为eclipse LV100P0L的显微镜(200×和400×),每一个样品均随机选择400粒左右的植硅体进行统计分析,不足400粒的样品全部统计,并用NikonBF53进行拍照。植硅体形态鉴定和分类按照国际植硅体命名法规[12]、现代植物植硅体分析结果和已经发表的文章中的植硅体照片进行鉴定和统计。水稻[13]、粟、黍[14]农作物的植硅体鉴定参考吕厚远等人的鉴定标准。黍、粟稃壳植硅体单片内包含的特殊纹饰多于2个的植硅体个体记入统计数量。方型、长方型、尖型、棒型等由于形态简单、缺乏表面特征,不具备植物种鉴别意义,但这些植硅体类型特别是其组合,对指示古环境具有重要意义[15],所以也进行统计。其中棒型、刺棒型植硅体单体长度大于l0μm的个体计入统计。植硅体的统计是计算每个植硅体类型的百分含量(单个类型数与总统计数的比值乘以100%),出土概率根据发现某种植物种类的样品数在样品总数中所占的比例计算。
三、鉴定结果
(一)炭化植物遗存鉴定结果
此次浮选出的炭化植物遗存有炭屑和植物种子(或果类)两大类(表一)。
1.炭屑
此次浮选出的炭屑总体较为细碎。大于1毫米的炭屑共计56.746g,每个单位的炭屑重量在0~9.842g不等,平均为0.079g/L,与淮河中游同时期的安徽宿州杨堡遗址(0.115g/L)[16]、蒙城尉迟寺遗址(0.27g/L)[17]相比相对较少,但与金寨遗址2016年的浮选结果(0.01g/L)[18]相比,炭屑的数量显著上升。这可能和2017年清理了北部生活区有关,生活区由于房屋倒塌和木材燃烧等原因,炭屑也会相应较多。
2.植物种子/果实
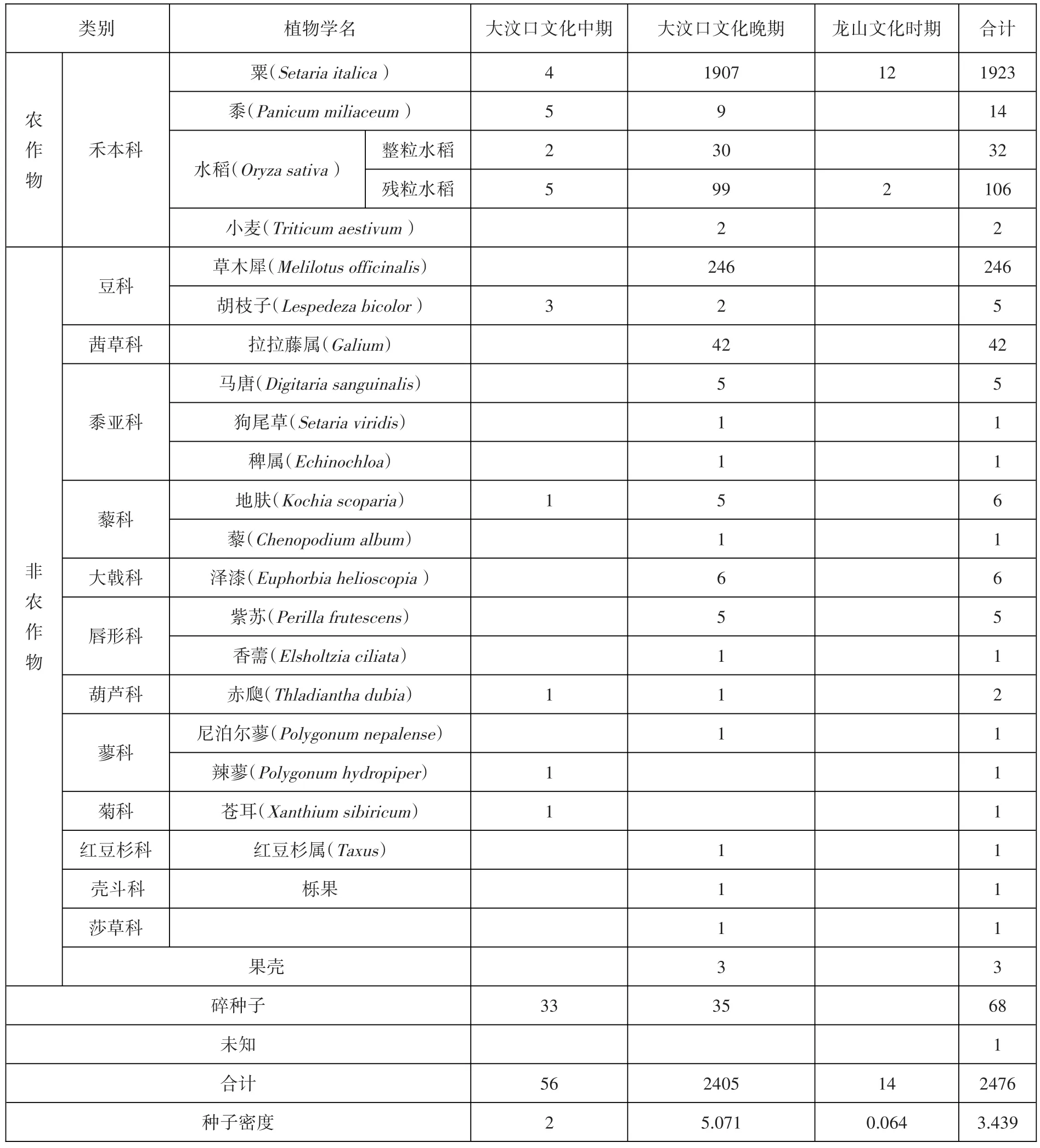
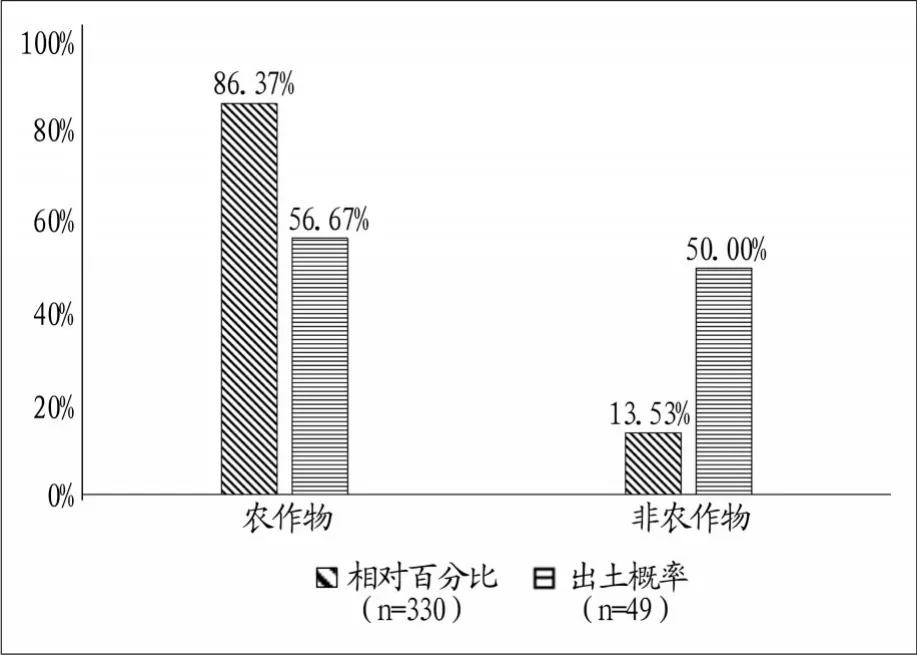
此次浮选发现的炭化植物种子/果类总计2476粒,平均密度为3.44粒/升。除去68粒特征不明显或由于炭化严重而失去特征部位无法鉴定的炭化植物种子,以及1粒未能鉴定出种属的种子外,剩下的2407粒可分为农作物和非农作物两大类。农作物包括粟、黍、水稻、小麦四种,共计2077粒,占比为86.29%,出土概率为55.10%。非农作物有草木犀、胡枝子、拉拉藤属、马唐、狗尾草、稗属、地肤、藜、泽漆、紫苏、香薷、赤瓟、尼泊尔蓼、辣蓼、苍耳、红豆杉属、栎果、莎草科和果壳,共计330粒,占比为13.71%,出土概率为36.73%(图一)。
(1)农作物
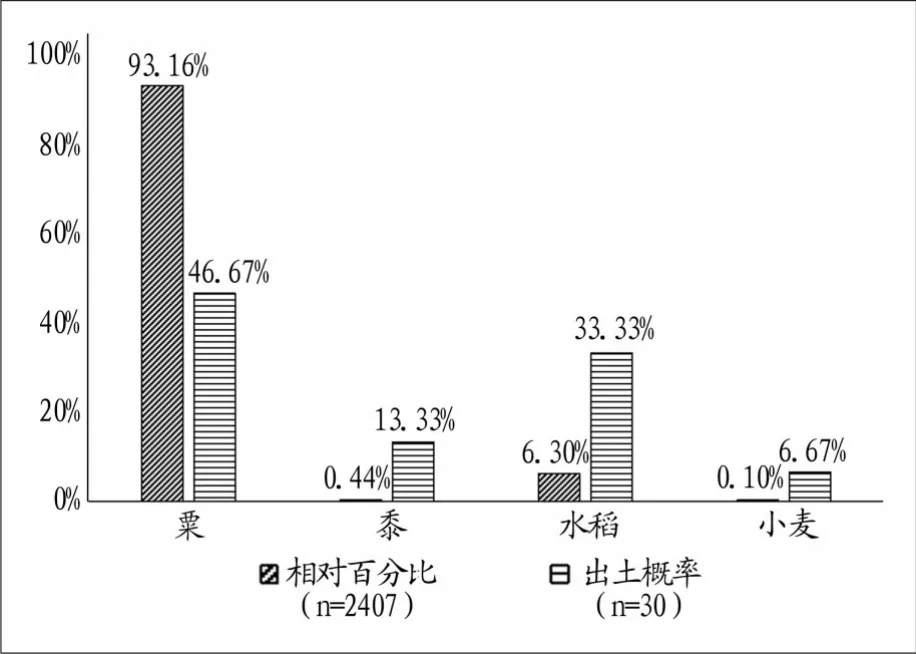
本次共浮选出粟、黍、水稻、小麦四种农作物,从绝对数量和出土概率两方面看,粟是金寨遗址最主要的农作物(图二)。
表一// 金寨遗址2017年出土植物遗存绝对数量统计表
粟:1923粒,其中粟1377粒,粟秕546粒。占农作物总数的92.59%,出土概率为46.93%,密度为2.67粒/升,是遗址主要的农作物类型之一。炭化粟粒大多数保存完整,近似圆形或椭圆形,表面光滑且有光泽,胚区较短,呈深V形。平均长约1.22、宽约1.17毫米(图三︰1)。
黍:14粒,占农作物总数的0.67%,出土概率为10.20%,密度为0.02粒/升。炭化黍粒保存状况一般,部分残破,胚区呈短U形。平均长约1.74、宽约1.70毫米(图三︰2)。
图一// 金寨遗址农作物、非农作物百分比和出土概率
图二// 金寨遗址农作物百分比及出土概率

图三// 金寨遗址出土的农作物种子
水稻:138粒,其中完整稻粒32粒,残块106粒。占农作物总数的6.64%,出土概率为26.53%,密度为19.17粒/升。炭化稻粒保存较差,大多残破,我们对完整的32粒进行测量,其平均长约4.44、宽约2.68毫米,长宽比约为1.657。按照目前主要的判别指标,一般长/宽<2.3为粳型稻,长/宽在2.31—2.5间为籼粳中间型,长/宽在2.51—3.5间为籼型,其中3—3.5的部分可能含有不典型的野生稻型,长/宽>3.51的为典型普通野生稻[19],金寨遗址此次发现的水稻均属粳型稻(图三︰3)。
小麦:2粒,占农作物总数的0.1%,出土概率为4.08%,密度为0.003粒/升。其中一粒保存较差,另一粒较好,长3.91~4.50、宽2.80~3.01毫米,长宽比为1.4—1.5(图三︰4)。
(2)非农作物
非农作物包括植物种子和果类遗存两类,共计330粒,属于12科18个种/属,其中以豆科的草木犀和胡枝子数量最多,茜草科次之,其余数量都较少。果壳3粒,因破碎失去主要特征而无法鉴定种属(图四)。
豆科:251粒,包括草木犀246、胡枝子5粒。占非农作物遗存的76.06%,出土概率为6.12%,密度为0.35粒/升。主要出土于H140②,共243粒,占豆科遗存总数的96.81%。草木犀保存较好,为椭圆肾形,表面光滑,平均长约1.28、宽约0.86毫米(图五︰1);胡枝子体积较小,斜结下有脐环,平均长约1.07、宽约0.84毫米。
茜草科:42粒,全部为拉拉藤属,占非农作物遗存的12.73%,出土概率为4.08%,密度为0.06粒/升。41粒出土于H74①,拉拉藤属为椭圆形或近卵形,稍扁平,平均长约1.11、宽约1毫米(图五︰2)。
黍亚科:7粒,包括马唐5、稗属1、狗尾草1粒(图五︰4)。占非农作物遗存的2.12%,出土概率为12.24%,密度为0.01粒/升。马唐为近椭圆形,表面较为光滑,胚区位于一端,平均长约2.11、宽约0.73毫米(图五︰3)。
藜科:7粒,包括地肤6、藜1粒。占非农作物遗存的2.12%,出土概率为6.12%,密度为0.01粒/升。地肤保存较好,呈卵形,中部扁凹,平均长约1.453、宽约1.021毫米(图五︰7)。
大戟科:6粒,只有泽漆一种,占非农作物遗存的1.82%,出土概率为10.2%,密度为0.01粒/升。泽漆种子呈卵形,表面有网状凹陷,平均长约2.03、宽约1.49毫米(图五︰8)。
唇形科:6粒,包括紫苏5,香薷1粒(图五︰5)。占非农作物遗存的1.82%,出土概率为8.16%,密度为0.01粒/升。紫苏表面较为光滑,有六边形的褶皱凸起,平均长约1.68、宽约1.51毫米(图五︰6)。

图四// 金寨遗址非农作物百分比和出土概率

图五// 金寨遗址出土的非农作物种子
葫芦科:2粒,只有赤瓟一种,占非农作物遗存的0.61%,出土概率为4.08%,密度为0.002粒/升。种子呈倒卵形,表面有褶皱,平均长约1.59、宽约1.38毫米。
蓼科:2粒,包括尼泊尔蓼1、辣蓼1粒。占非农作物遗存的0.61%,出土概率为4.08%,密度为0.002粒/升。辣蓼保存完好,呈三棱状,表面较为光滑(图五︰9)。
莎草科:1粒,占非农作物遗存的0.30%,出土概率为2.04%,密度为0.001粒/升。
菊科:苍耳1粒,占非农作物遗存的0.30%,出土概率为2.04%,密度为0.001粒/升。
红豆杉科:红豆杉属1粒,占非农作物遗存的0.30%,出土概率为2.04%,密度为0.001粒/升(图五︰10)。
栎果:1粒,占非农作物遗存的0.30%,出土概率为2.04%,密度为0.001粒/升。
果壳:3粒,占非农作物遗存的0.91%,出土概率为4.08%,密度为0.004粒/升。
(二)植硅体鉴定结果
除了H93,其余32份样品全部发现了植硅体,鉴定出22种植硅体类型,共计10816粒个体。农作物植硅体发现的数量和出土概率都很少。大汶口文化晚期的样品里仅在6份样品中发现了来自水稻茎叶的扇型植硅体和来自水稻稃壳的双峰乳突型植硅体,1份样品中发现了黍稃壳η型。龙山文化时期的7份样品中发现了水稻扇型(出土概率53.8%),不见其他农作物类型。其他可鉴定的植硅体类型主要有长方型、方型、平滑棒型、刺棒型、扇型、哑铃型、帽型、短鞍型、短尖型、中鞍型、齿型、导管型、芦苇扇型等,此外还有海绵骨针和硅藻等生物化石。
四、分析与讨论
(一)农作物结构和生计方式

图六// 金寨遗址主要植物遗存与探方的对应分析
此次所取土样的年代分别为大汶口文化中期、大汶口文化晚期和龙山文化时期,但大汶口文化中期只有1份来自探沟的样品,21L土样中发现了粟4粒、黍5粒以及水稻7粒(2粒完整、5粒残块),由于样品较少暂不进行探讨。下面我们分别讨论大汶口文化晚期和龙山文化时期的农作物结构和生计方式。
1.大汶口文化晚期
大汶口文化晚期发现的农作物有粟、黍、水稻和小麦,共计2047粒。粟共发现1907粒,占农作物总数的93.16%,出土概率为46.67%,说明粟是金寨聚落主要的农作物。水稻发现129粒,仅占农作物总数的6.30%,出土概率为33.33%,其地位仅次于粟。黍和小麦分别发现9粒和2粒,出土概率也很低(13.33%和6.67%),在金寨聚落农作物结构中所占比重很低。金寨遗址虽然没有发现水稻基盘,但发现了稻田常见杂草稗,来自稻茎叶的扇型和来自稻壳的双峰型植硅体也在多个遗迹中发现,说明金寨的水稻是本地种植的。水稻作为大粒作物,与粟、黍小粒作物相比,相同数量的粟、黍和水稻对先民的食物贡献率差异较大,经过千粒重[20]的换算后,水稻的比例明显上升,虽然粟仍然是对金寨先民食物结构贡献最大的作物,但水稻应该也是金寨聚落中非常重要的农作物,金寨遗址的农业模式是一种旱稻混作的结构,稻作农业种植规模可能稍小于旱作农业。2016年的浮选样品中未发现炭化水稻,仅发现水稻的植硅体[21],所以未能比较旱作农业和稻作农业在金寨遗址的重要性程度,2017年的浮选结果弥补了这一不足。
此外,金寨遗址也出土了较多的非农作物种子。藜全草可入药,嫩叶可食用[22],它在美洲和台湾等地是一种有着较长栽培历史的栽培作物[23],汉阳陵陪葬坑中藜属种子和粟、黍、小麦共出,可能是最早栽培藜属的证据[24]。紫苏是一种香料,可去除腥味,紫苏的叶、梗和籽粒均能食用,种子可榨油供食用[25]。紫苏和藜在史前到商周时期的遗址中普遍发现,因此,笔者推测金寨先民可能有意采摘它们用作时令蔬菜。遗址中还发现了栎果和破碎的果壳,金寨先民可能也采摘一些果类作为其食物资源的补充。动物骨骼鉴定结果显示,在金寨聚落出土的动物骨骼中猪骨占绝对主导地位,其次是少量的鹿[26]。笔者推测金寨聚落的生业经济模式是粟、黍、水稻种植和家猪饲养相结合,平时还会采集一些野菜和果类以及部分野生肉食动物作为食物资源的补充。
此次浮选发现的2粒小麦分别出土于大汶口文化晚期(3000—2600BC)的H74②和H81④中,当前有明确测年数据的年代最早的小麦发现于龙山文化时期山东胶州赵家庄遗址,测年数据为2500—2270BC[27],本次发现的2粒小麦年代较早,需要对样品进行测年再进一步讨论。
2.龙山文化时期
龙山文化时期的炭化种子仅有14粒农作物,但出土概率达到了50%。发现粟12、水稻2粒,出土概率分别为44.44%和11.11%。虽然这一时期的种子数量很少,但仅从占比和出土概率来看,粟的地位仍然高于水稻,经过千粒重的换算后,水稻的地位超过了粟。从植硅体的数据来看,龙山文化时期仅发现了来自水稻茎叶的扇型植硅体,出土概率达到了53.80%,所以笔者认为龙山文化时期水稻的比例上升,甚至可能超过了粟黍。淮河中游的宿州芦城孜[28]和尉迟寺遗址[29]到龙山文化时期水稻也超过了粟黍,这可能是淮河流域稻作农业发展的结果。
(二)植物遗存的空间分布反映的遗址功能分区
多元统计分析可以从不同角度整合数据,揭示变量之间是否存在联系或存在什么样的联系。笔者对六种出土数量较多的炭化植物遗存和出土位置进行对应分析(Correspondence Analysis)[30],并以此讨论植物遗存的空间分布以及不同种类植物遗存之间的关系(图六)。为了更加全面地探讨遗址的功能分区等问题,本文把2016年的浮选数据也纳入统计。此外,植硅体可以区分来自作物的茎秆或者稃壳,这恰好是炭化植物遗存不易保存的部分。把植硅体数据和炭化植物遗存的空间分布相结合可以更准确地探讨植物在遗址中的分布模式以及它可能代表的人类活动。
分析结果显示粟黍与水稻的出土不具有相关性,水稻和粟黍可能由于不同的人类活动造成了分布的差异。马唐和草木犀都是常见的田间杂草,金寨遗址的马唐、草木犀和粟黍都具有较强的相关性。马唐是常见的旱地杂草,草木犀虽然可以作为牧草或者燃料使用,但在金寨遗址中并没有体现出较为特殊的利用方式,可能仅是一般的农田杂草,并和马唐一起伴随着粟黍收割被带入遗址。稗是常见的稻田杂草,但此次浮选仅发现了1粒,可能是和水稻一起被收割并带入遗址的。
为了进一步探讨粟黍和水稻的分布规律以及它可能指示的人类活动,本文对植物遗存的空间分布进行了分析后可知,东区所出的农作物密度较北区更大。水稻和北区的探方关系更为密切,多集中出土于北区的探方,只有8粒出自东区,粟黍集中出土于东区。从植硅体的数据看,东区的5份样品中发现了来自水稻茎叶的扇型植硅体,其中在2份样品中还发现了水稻稃壳植硅体,而北区仅在1份样品中发现了1粒水稻扇型植硅体。2016年的样品分析结果显示东区没有发现炭化水稻,但较为普遍地发现了来自水稻茎叶和稃壳的植硅体,这和2017年的分析结果一致。干净的稻谷粒在北区集中出土,而水稻茎叶和稃壳却集中出土于东区,据此,笔者推测东区是水稻的加工区域(脱粒和脱壳),而北区是水稻的消费区域。东区和北区都有粟黍出土,但东区出土粟黍的密度更大,因此东区应该消费了更多的粟黍。
水稻在东区进行脱粒脱壳却集中在北区进行消费,粟黍在东区脱壳且多在东区进行消费,少量在北区消费,造成这种差异的原因可能有两种:第一,东区居民和北区居民有不同的食物偏好。金寨遗址处于南北文化交流通道上,大汶口文化晚期墓葬中也出土了良渚文化典型器物双鼻壶,以及屈家岭文化典型器物瓦形足盆形鼎、盂形杯,因此笔者推测北区居民对水稻的偏好可能受南方食稻人群的影响。这种影响是人口迁移还是文化因素扩散导致的,尚需要碳氮和锶同位素的证据。第二,大汶口文化晚期出现了等级分化,此时已形成大、中、小三级聚落的分层结构,社会内部也随着贫富分化的加剧和分层的发展,出现了统治和被统治阶级[31]。北区的居民可能是金寨聚落的特权阶层,谷物在东区脱粒脱壳后供他们食用;水稻作为金寨聚落较为稀有的农作物,集中在北区出土,可能代表了不同等级居民之间的食物差异,水稻可能更多的供高等级人群食用,这也需要进一步整理发掘材料进行探讨。
(三)淮河中游地区龙山时代的农业
淮河中游在裴李岗时代晚期出现了旱稻混作农业[32]。到了龙山时代,金寨、尉迟寺[33]、杨堡[34]、芦城孜[35]、禹会村[36]和平粮台遗址[37]的农业模式均为旱稻混作模式,仰韶时代(5000—3000BC)应该是淮河中游地区农业发展的关键期。我们此次虽然仅有1份仰韶时代(大汶口文化中期)的样品,但在该样品中发现了4粒粟和5粒黍,大汶口文化中期粟黍出现在金寨遗址中,为我们进一步探讨淮河中游地区稻作到旱稻混作农业模式的转变提供了新的证据。

图七// 淮河中游地区不同遗址农作物占比对比图
从大汶口文化晚期到龙山文化时期,也就是龙山时代(3000—2000BC),金寨遗址的农业结构一直是旱稻混作,且以旱作农业为主。本文将淮河中游发表的同时期的植物遗存数据进行统计,并对比了各遗址粟、黍、稻三者的比例(图七)[38]。从图上可以看出,靠东、靠南的遗址(宫庄、禹会村、芦城孜和杨堡遗址)稻作农业占比高,靠西、靠北的遗址(平粮台、金寨和尉迟寺遗址)旱作农业占比高。现代淮河流域水稻种植的主要限制因子是降水[39],靠东、南的遗址降水较多,更加适合水稻的种植。但目前淮河流域尚缺乏高分辨率的古气候环境记录,所以不同区域农业结构的差异与气候环境的关系有待进一步研究[40]。另一方面应考虑到稻作农业由南向北扩散的历程与周期,稻作农业传入海岱地区有两条路线[41],东线沿海,西线经由淮河中游向海岱地区中心区域传播,但都是自南向北传播过来的。由于西线是一条次要路线,所以稻作传播的速率没有东线那么快,因此西线北部的遗址对于稻作的接受度没有南部高。
五、结语
2017年的浮选结果显示,金寨聚落大汶口文化晚期的农业模式都是以种植粟为主,水稻为辅的旱稻混作模式,龙山文化时期稻作的比例略有提升。旱稻混作是淮河流域在龙山时代最显著的特征,但偏南、偏东的遗址(杨堡、芦城孜、宫庄和禹会村遗址)比偏北、偏西的遗址(金寨和平粮台遗址)的稻作比例高,这可能是受气候或稻作农业传播速率的影响。
对应分析结果显示,粟黍和水稻的加工可能都在东区,但水稻集中在北区消费,粟黍多在东区。这一现象说明金寨遗址东区与北区的居民可能有不同的食物偏好,北区居民可能受到更多南方食稻人群的影响;或者东区的居民有更高的社会地位,较为稀有的水稻脱粒脱壳后供他们食用。造成这种差异的原因还需要进一步整理发掘资料来进行探讨。
此次浮选出的龙山文化时期的种子较少,这可能与发掘区的位置有关——在龙山文化时期,整个聚落的范围向东北推移,而发掘区恰好位于这一时期聚落的边缘地带,人类活动较少,遗留的植物遗存自然也较少,这也从植物考古的角度印证了龙山文化时期金寨先民活动范围的变化。
[1]栾丰实:《淮河中游地区文明探源的新进展——读〈蚌埠禹会村〉》,《考古》2014年第10期。
[2]高广仁、邵望平:《析中国文明主源之一——淮系文化》,《东方考古》(第1集),科学出版社2004年,第36—64页。
[3]苏秉琦:《略谈我国东南沿海地区的新石器时代考古——在长江下游新石器时代文化考古学术讨论会上的一次发言提纲》,《文物》1978年第3期。
[4]严文明:《农业发生与文明起源》,科学出版社2000年,第338页。
[5]Jianping Zhang,Houyuan Lu,Wanfa Gu,et al.Early mixed farming of millet and rice 7800 years ago in the mid⁃dle Yellow River region,China.PLoS One,2012(7):1-8.
[6]Wuhong Luo,Chunguang Gu,Yuzhang Yang,et al.Phyto⁃liths reveal the earliest interplay of rice and broomcorn millet at the site of Shuangdun(ca.7.3-6.8 ka BP)in the middle Huai River valley,China.Journal of Archaeologi⁃cal Science.2019(102):26-34.
[7]孙波:《再论大汶口文化向龙山文化的过渡》,《古代文明》(第6卷),文物出版社2007年。
[8]a.张小雷:《安徽萧县金寨遗址发现大汶口至龙山文化遗迹》,《中国文物报》2018年12月14日第8版;b.安徽省文物考古研究所、萧县博物馆:《安徽萧县金寨新石器时代遗址北区2017年发掘简报》,《东南文化》2020年第3期。
[9]赵志军:《植物考古学的田野工作方法——浮选法》,《考古》2004年第3期。
[10]a.郭琼霞:《杂草种子鉴定图鉴》,中国农业出版社1998年;b.关广清等著:《杂草种子图鉴》,科学出版社2000年;c.国家林业局国有林场和林木种苗工作总站主编:《中国木本植物种子》,中国林业出版社2003年;d.刘长江、靳桂云、孔昭宸编著:《植物考古:种子、果实》,科学出版社2008年;e.赵志军:《植物考古学:理论、方法和实践》,科学出版社2010年;f.郭巧生等编著:《中国药用植物种子原色图鉴》,高等教育出版社2011年。
[11]王永吉、吕厚远:《植物硅酸体研究及应用》,海洋出版社1993年。
[12]李泉、吕厚远、王伟铭:《国际植硅体命名法规(Inter⁃national Code for Phytolith Nomenclature 1.0)的介绍与讨论》,《古生物学报》2009年第1期。
[13]a.Pearsall D.M.,Piperno D.R.,Dinan E.H.,et al.Dis⁃tinguishing rice(Oryza sativa Poaceae)from wild Oryza species through phytolith analysis:results of preliminary research.Economic Botany,1995,49(2):183-196.b.Fujiwara H.Research into the history of rice cultiva⁃tion using plant opal analysis.In:Pearsall D.M.&Piper⁃no D.R.(eds.),Current Research in Phytolith Analysis,Application in Archaeology and Paleoecology.MASCA,University of Pennsylvania,Philadelphia(PA)1993:147-158.c.Zhijun Zhao,Pearsall D.M.,Benfer R.A.,et al.Dis⁃tinguishing rice(Oryza sativa poaceae)from wild Oryza species throuth phytolith analysis,Ⅱ Finalized method.Economic Botany,1998,52(2):134-145.d.Gu Y.S.,Zhao Z.J.,Pearsall D.M.Phytolith morphol⁃ogy reearch on wild and domesticated rice species in East Asia.Quaternary International,2013(287):141-148.
[14]Houyuan Lu,Jianping Zhang,et al.Phytolith analysis for the discrimination of Foxtail millet(Setaria italica)and Common millet(Panicum miliaceum).PLoS ONE,2009(4):e4448.
[15]王永吉、吕厚远:《植物硅酸体研究及应用》,海洋出版社1993年,第164—181页。
[16]程至杰、杨玉璋、袁增箭等:《安徽宿州杨堡遗址炭化植物遗存研究》,《江汉考古》2016年第1期。
[17]赵志军:《安徽蒙城尉迟寺遗址浮选结果分析报告》,《植物考古学:理论、方法和实践》,科学出版社2010年,第109—119页。
[18]杨凡、张小雷、靳桂云:《安徽萧县金寨遗址(2016年)植物遗存分析》,《农业考古》2018年第4期。
[19]王象坤、孙传清主编:《中国栽培稻起源与演化研究专集》,中国农业大学出版社1996年。
[20]粟千粒重的平均值为3.1g,水稻千粒重的平均值为26g,数据来源于中华人民共和国国家标准粮食、油料检验千粒重测定法。
[21]同[18]。
[22]贺学礼主编:《植物学》,高等教育出版社2004年,第337页。
[23]Smith B.D.Chenopodium as a prehistoric domesticate in Eastern North America:Evidence from Russel Cave,Al⁃abama.Science,1984,226:165-167.
[24]杨晓燕、刘长江、张健平:《汉阳陵外藏坑农作物遗存分析及西汉早期农业》,《科学通报》2009年第54卷第13期。
[25]李扬汉主编:《中国杂草志》,中国农业出版社1998年,第571页。
[26]宋艳波、乙海琳、张小雷:《安徽萧县金寨遗址(2016、2017)动物遗存分析》,《东南文化》2020年第3期。
[27]靳桂云、王海玉、燕生东等:《山东胶州赵家庄遗址龙山文化炭化植物遗存研究》,《科技考古》(第三辑),科学出版社2011年,第36—53页。
[28]王育茜、陈松涛、贾庆元等:《安徽宿州芦城孜遗址2013年度浮选结果分析报告》,《海岱考古》(第九辑),科学出版社2016年,第365—380页。
[29]同[17]。
[30]对应分析(Correspondence Analysis)也称关联分析,是一种视觉化的数据分析方法,它可以揭示同一变量各个类别之间的差异,也可以体现不同变量各个类别之间的对应关系,在样本量较多的情况下有显著的实用性和有效性。本文使用SPSS软件进行数据分析。
[31]栾丰实:《大汶口文化的社会发展进程研究》,《古代文明》(第2卷),文物出版社2003年。
[32]杨玉璋、程至杰、李为亚等:《淮河上、中游地区史前稻—旱混作农业模式的形成、发展与区域差异》,《中国科学:地球科学》2016年第8期。
[33]同[17]。
[34]同[16]。
[35]同[28]。
[36]尹达:《禹会村遗址浮选结果分析报告》,中国社会科学院考古研究所、安徽省蚌埠市博物馆编《蚌埠禹会村》,科学出版社2013年。
[37]赵珍珍:《淮河中游龙山时代农业研究》,山东大学硕士学位论文,2018年。
[38]a.程至杰:《淮河上中游地区新石器时代植食性资源利用研究》,中国科学技术大学博士学位论文,2016年;b.罗武宏、顾纯光、杨玉璋:《安徽临泉宫庄遗址大汶口文化早期与龙山文化时期稻作农业发展的植硅体证据》,《微体古生物学报》2018年第4期;c.程至杰、杨玉璋、张东:《安徽临泉宫庄遗址炭化植物遗存分析》,《农业考古》2019年第3期。因不清楚原文中所指的大汶口文化具体属于哪一期,芦城孜遗址只取龙山文化的数据;其余遗址有大汶口文化晚期的与龙山文化合并计算,没有的只单独取龙山文化数据。
[39]俞芬、千怀遂、段海来:《淮河流域水稻的气候适宜度及其变化趋势分析》,《地理科学》2008年第4期。
[40]同[32]。
[41]栾丰实:《海岱地区史前时期稻作农业的产生、发展和扩散》,《文史哲》2005年第6期。
