薏苡苗期紫叶鞘性状的遗传分析
2020-08-28陆秀娟李祥栋石明潘虹魏心元陆平
陆秀娟 李祥栋 石明 潘虹 魏心元 陆平


摘要:为阐明薏苡苗期紫叶鞘性状的遗传方式和遗传规律,指导薏苡的品种选育,利用5份紫叶鞘材料为母本,1份绿色叶鞘材料为父本,分析不同杂交世代F1、F2的叶鞘色表型性状及其遗传规律。结果表明,紫叶鞘相对于绿叶鞘为显性,对于不同的F2群体,植株表现出紫叶鞘 ∶绿叶鞘=9 ∶7和紫叶鞘 ∶绿叶鞘=3 ∶1的分离比。因此,在不同的遗传背景中,薏苡苗期紫色叶鞘的遗传方式不同,表现为2对基因的显性互补遗传和单基因显性遗传2种类型。紫叶鞘可作为薏苡遗传改良的标记性状。
关键词:叶鞘色;花色素苷;杂交;性状;遗传规律;薏苡;单株提纯
中图分类号:S519.032 文献标志码: A
文章编号:1002-1302(2020)14-0146-04
花色素苷是类黄酮生物合成途径中的产物,也是植物的重要次生代谢产物之一,通常在植物的花、果实、种子、茎秆、叶片、叶鞘、柱头等部位积累,导致植物出现一系列的颜色变化。薏苡属于禾本科植物,是重要的药食兼用作物之一,具有抗肿瘤、降血糖、降血脂、镇痛和美化肌肤等多种生理功效[1-2]。薏苡具有多种资源类型,在种质的考察收集[3-5]以及表型[6-8]、品质[9]、抗逆性[10-13]等的分析评价方面也时有报道,但是由于长期处于野生或半野生状态,基本上没有被进行遗传改良。尽管也有学者尝试进行薏苡属不同种(变种)的杂交工作,但是有关薏苡质量性状和数量性状的遗传规律研究仍处于空白状态[14-17]。薏苡苗期的叶鞘色具有白色、绿色和紫色之分,并以绿色和紫色最为常见,可作为区分不同品种或种质的重要标记性状之一[8,18]。本研究分别以绿色和紫色叶鞘品种为亲本进行杂交并构建F2遗传群体,旨在阐明薏苡紫色叶鞘性状的遗传规律,并为薏苡的杂交育种实践提供重要指导。
1 材料与方法
1.1 试验材料
供试材料为6份经多年提纯的薏苡品种(系),其中兴仁白壳、黔薏2号、薏珠1号、安薏1号和安国薏苡为母本(♀)材料;CL91品系为共用父本(♂)材料。上述材料均经过笔者所在课题组多年单株提纯,可保证亲本的纯合性。试验材料性状表现及来源见表1。
1.2 试验方法
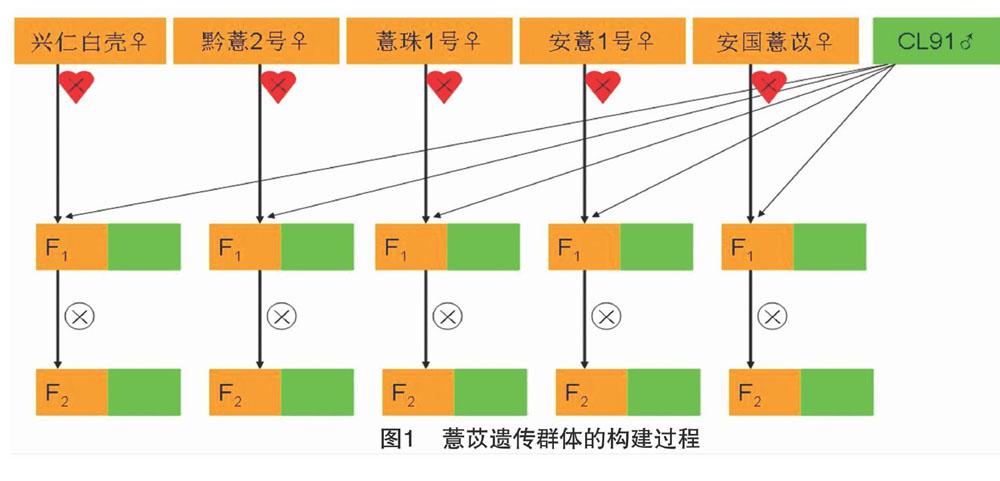
于2016年在贵州省兴义市黔西南布衣族苗族自治州农业科学研究所试验基地进行杂交试验,获得5组F1,第2年采用F1自交获得F2种子,第3年播种获得F2分离群体(图1)。通过目测鉴别各世代苗期叶鞘的颜色差异,并统计具有绿色和紫色叶鞘的株数。
1.3 数据统计
利用Excel 2003对统计的数据进行χ2测验,检验各性状的理论分离比例与实际分离比例的符合度,分析紫色叶鞘性状的遗传规律。
采用矫正后的χ2计算符合度:χ2=∑[SX(](|A-T|-0.5)2T[SX)],其中A为实际观测次数,T为理论次数。
2 结果与分析
2.1 F1代幼苗的叶鞘颜色表现
在5个杂交组合的F1植株中,4个紫色(♀)×绿色(♂)的杂交组合兴仁白壳(♀)×CL91(♂)、黔薏2号(♀)×CL91(♂)、薏珠1号(♀)×CL91(♂)和安薏1号(♀)×CL91(♂)的F1植株均表现为紫色叶鞘,而1个绿色(♀)×绿色(♂)叶鞘的杂交组合安国薏苡(♀)×CL91(♂)的F1植株全部为绿色叶鞘(表2、图2),说明薏苡的紫色叶鞘相对于绿色叶鞘为显性。
2.2 不同杂交组合F2代叶鞘颜色的分离
F2植株调查结果(表3)显示,在4个紫色(♀)×绿色(♂)的杂交组合中,叶鞘颜色在F2植株中出现了性状分离,而在绿色(♀)×绿色叶鞘的杂交组合安国薏苡(♀)×CL91(♂)中,F2植株全部表现为绿色叶鞘,并未表现出性状分离,证明绿色叶鞘是隐性基因控制的遗传性状。χ2检验发现,兴仁白壳(♀)×CL91(♂)和黔薏2 号(♀)×CL91(♂)杂交组合的紫色和绿色叶鞘植株分离比为9(紫色):7(绿色),而薏珠1号(♀)×CL91(♂)和安薏1号(♀)×CL91(♂)的紫色和绿色叶鞘植株符合 3 ∶1 的分离比。说明亲本兴仁白壳和黔薏2号的紫色叶鞘是由2对互补基因控制的显性遗传性状,而薏珠1号和安薏1号的紫色叶鞘则由单基因控制,属于单基因显性遗传。根据上述结果可知,薏苡苗期的紫色叶鞘性状在不同遗传背景中遗传方式不同,表现为2对基因控制的显性互补遗传和单基因显性遗传2种类型。
3 讨论
3.1 花色素苷积累及其性状遗传
植物花色素苷合成途径属于类黄酮代谢的分支途径,其代谢路径已经相当清楚。花色素苷合成路径中的关键酶主要有查尔酮合成酶(CHS)、查尔酮异构酶(CHI)、黄烷酮-3-羟化酶(F3H)、二氢黄酮醇-4-还原酶(DFR)、花青素合成酶(ANS)等[19-20],受多种转录因子和光照等环境因子调控[21]。已有研究表明,内蒙古紫皮小麦、山农066559小麦的紫粒性状均由2对独立的显性互补基因控制[22-23],但是紫色叶鞘性状由1对显性基因控制[22];水稻紫色叶片、叶鞘、稃尖、柱头的遗传方式也表现出受多对基因(2~4对)互作控制或受显性单基因控制[24]。李慧敏等通过旱稻和高粱的属间杂交发现,紫芒性状由2对隐性互作的上位基因控制,而紫色柱头性状则由2对互补基因控制[25]。在大白菜中,紫心性状则由1对显性基因控制[26]。本研究也表明,薏苡苗期叶鞘色分别受2对显性互作基因和1对显性基因控制,其遗传方式比较简单,与上述小麦、水稻等作物的紫色性状遗传规律基本一致。
3.2 标记性状在育种实践中的应用
由于长期以来薏苡很少進行定向遗传改良,因而现今在生产中应用的品种基本上都是对自然资源系统选择而育成的品种。一方面,因为薏苡分蘖和分枝的生物学特性,不同部位籽粒成熟度不一致,整枝、去雄程序繁琐;另一方面,由于关于薏苡的性状遗传规律研究空白,缺乏有效区分F1植株中是否存在自交个体的标记。本研究在杂交组合的配制过程中,采用人工标记总苞的方法,以保证薏苡单个柱头的定向授粉和本研究群体的有效性。通过本研究对紫叶鞘遗传规律的揭示可知,紫叶鞘可以作为F1植株除杂的有效标记性状,指导薏苡的遗传改良工作。此外,分子标记辅助育种作为现代育种的重要手段,能够极大提高选择效率,加快育种进程[27]。目前有关水稻的叶鞘、柱头、稃尖和芒的紫色性状[28]及小麦籽粒性状[29-30]的基因定位研究也时有报道。此外,花色素苷除了在叶鞘中合成外,在薏苡的茎秆、柱头、苞状鞘、幼果、成熟总苞等多个部位均有累积。本研究中F2遗传群体的构建,为进一步阐明作物紫色标记性状的遗传规律及其基因定位打下了坚实基础。
4 结论
通过构建不同的F2遗传群体,初步阐明了薏苡苗期紫叶鞘的遗传规律,在不同的遗传背景材料中,薏苡叶鞘色分别由2对显性互补基因和1对显性单基因控制,紫叶鞘可作为薏苡标记性状指导薏苡的定向遗传改良。
参考文献:
[1]黄锁义,李 容,潘 勇,等. 薏苡研究的新进展[J]. 食品研究与开发,2012,33(11):223-227.
[2]杨 爽,王李梅,王姝麒,等. 薏苡化学成分及其活性综述[J]. 中药材,2011,34(8):1306-1312.
[3]李英材,覃祖贤. 广西薏苡资源形状分析与分类[J]. 西南农业学报,1995,8(4):109-113.
[4]陆 平,左志明. 广西薏苡资源的分类研究[J]. 广西农业科学,1996(2):81-84.
[5]梁云濤,陈成斌,梁世春,等. 中日韩三国薏苡种质资源遗传多样性研究[J]. 广西农业科学,2006,37(4):341-344.
[6]王 硕,张世鲍,何金宝,等. 薏苡资源性状的主成分和聚类分析[J]. 云南农业大学学报,2013,28(2):157-162.
[7]李春花,王艳青,卢文洁,等. 云南薏苡种质资源农艺性状的主成分和聚类分析[J]. 植物遗传资源学报,2015,16(2):277-281,287.
[8]李祥栋,潘 虹,陆秀娟,等. 薏苡属种质资源的主要表型性状多样性研究[J]. 植物遗传资源学报,2019,20(1):229-238.
[9]李祥栋,潘 虹,陆秀娟,等. 薏苡种质的主要营养组分特征及综合评价[J]. 中国农业科学,2018,51(5):835-842.
[10]汪 灿,周棱波,张国兵,等. 薏苡种质资源萌发期抗旱性鉴定及抗旱指标筛选[J]. 植物遗传资源学报,2017,18(5):846-859.
[11]汪 灿,周棱波,张国兵,等. 薏苡种质资源苗期抗旱性鉴定及抗旱指标筛选[J]. 中国农业科学,2017,50(15):2872-2887.
[12]汪 灿,周棱波,张国兵,等. 薏苡种质资源成株期抗旱性鉴定及抗旱指标筛选[J]. 作物学报,2017,43(9):1381-1394.
[13]罗 凯,黎 青,杨成龙,等. 薏苡黑穗病田间发病形态观察及抗性鉴定[J]. 江苏农业科学,2019,47(20):126-128.
[14]乔亚科,李桂兰,高书国,等. 薏苡类型间杂交F2代的性状分离[J]. 河北农业技术师范学院学报,1993,7(4):48-51.
[15]乔亚科,李桂兰,高书国,等. 两种类型薏苡及其F1性状表现[J]. 中草药,1995(2):88-91.
[16]宋秀英,赵晓明,李明山,等. 薏苡属种间杂种形态的观察和比较[J]. 山西农业大学学报,1993,13(3):199-202,294.
[17]李贵全,赵晓明,宋秀英. 薏苡×川谷远缘杂交的研究[J]. 作物学报,1997,23(1):119.
[18]石 明,李祥栋,秦礼康. 薏苡种质资源描述规范和数据标准[M]. 北京:中国农业出版社,2017.
[19]张 龙,李卫华,姜淑梅,等. 花色素苷生物合成与分子调控研究进展[J]. 园艺学报,2008,35(6):909-916.
[20]王 蕾,韦灵林,徐 虹. 矮牵牛花色素苷合成途径中的关键酶及其转录调控[J]. 细胞生物学杂志,2009,31(5):641-650.
[21]李小兰,张明生,任明见,等. 紫粒小麦花青素合成分子调控机制研究进展[J]. 植物生理学报,2017,53(4):521-530.
[22]邵中子,邓景扬. 内蒙古紫皮小麦紫皮性状的遗传分析[J]. 遗传学报,1987,14(3):179-187,243.
[23]李楠楠,张卫东,高庆荣,等. 小麦山农066559紫色性状的遗传分析[J]. 麦类作物学报,2013,33(1):18-22.
[24]曾黎琼,段玉云,孙一丁,等. 水稻紫色性状的遗传分析和基因定位[J]. 西南农业学报,2010,23(1):256-260.
[25]李慧敏,李道亨,赵凤梧,等. 旱稻、高粱属间杂交紫色性状遗传分析[J]. 华北农学报,2006,21(增刊2):84-86.
[26]吴俊清,赵 静,秦美玲,等. 大白菜紫心性状遗传规律分析及其基因初步定位[J]. 园艺学报,2016,43(6):1079-1088.
[27]王亚琦,孙子淇,郑 峥,等. 作物分子标记辅助选择育种的现状与展望[J]. 江苏农业科学,2018,46(5):6-12.
[28]李 静,孙 妍,齐 兰,等. 基于染色体置换系的野生稻抽穗期及紫色性状QTL的鉴定[J]. 植物遗传资源学报,2018,19(3):568-577.
[29]Dobrovolskaya O,Arbuzova V S,Lohwasser U,et al. Microsatellite mapping of complementary genes for purple grain colour in bread wheat (Triticum aestivum L.)[J]. Euphytica,2006,150(3):355-364.
[30]Li X P,Lan S Q,Zhang Y L,et al. Identification of molecular markers linked to the genes for purple grain color in wheat (Triticum aestivum)[J]. Genetic Resources and Crop Evolution,2010,57(7):1007-1012.
收稿日期:2019-07-24
基金项目:贵州省科技计划(编号:黔科合重大专项字[2014]6023;黔科合支撑[2016]2608号);贵州省高层次创新型人才培养项目(编号:黔科合人才[2015]4016号)。
作者简介:陆秀娟(1989—),女,布依族,贵州册亨人,农艺师,主要从事作物遗传及栽培技术及研究。E-mail:lowaterve_643@126.cm。
通信作者:李祥栋,硕士,高级农艺师,主要从事植物生理与分子调控研究。E-mail:lixiangdongsiji@163.com;石 明,硕士,研究员,主要从事作物遗传育种方向的研究。E-mail:shiming1616@126.com。
