蠋蝽对小菜蛾幼虫的捕食作用
2020-08-25唐艺婷郭义潘明真毛建军陈红印张礼生王孟卿
唐艺婷 郭义 潘明真 毛建军 陈红印 张礼生 王孟卿
摘要 本试验在室内研究了蠋蝽3~5龄若虫及成虫对小菜蛾幼虫的捕食能力,采用HollingⅡ功能反应模型和Hassell模型,以明确蠋蝽对小菜蛾的捕食潜能以及蠋蝽自身密度对小菜蛾捕食作用的影响。结果表明,蠋蝽3~5龄若虫及成虫对小菜蛾幼虫的捕食模型均符合HollingⅡ模型;蠋蝽3~5龄若虫和成虫瞬时攻击率由大到小依次为1.461(3龄若虫)>1.322(4龄若虫)>1.086(5龄若虫)>0.803(雌成虫)>0.734(雄成虫);蠋蝽5龄若虫的日最大捕食量最高,为83.33头。随着小菜蛾幼虫密度增大,蠋蝽的搜寻效应逐渐降低,5龄若虫的搜寻效应明显大于其他虫态。蠋蝽3~5龄若虫及成虫自身密度对其捕食作用的干扰效应明显,其中蠋蝽3龄若虫的竞争参数最大,为0.862。本研究表明蠋蝽对小菜蛾具有较好的捕食潜力,尤其是蠋蝽5龄若虫。
关键词 蠋蝽; 小菜蛾; 功能反应; 生物防治
中图分类号: S 476.2
文献标识码: A
DOI: 10.16688/j.zwbh.2019243
Predation of Plutella xylostella larva by Arma chinensis
TANG Yiting1, GUO Yi2, PAN Mingzhen3, MAO Jianjun1, CHEN Hongyin1,
ZHANG Lisheng1, WANG Mengqing1*
(1. Institute of Plant Protection, Chinese Academy of Agricultural Sciences, USDA-ARS Sino-American
Biological Control Laboratory, Beijing 100193, China; 2. Institute of Plant Protection, Guangdong Academy
of Agricultural Sciences, Guangzhou 510640, China; 3. College of Plant Health and Medicine,
Qingdao Agricultural University, Qingdao 266109, China)
Abstract
The predation of Plutella xylostella by the 3rd-5th instar nymphs and adults of Arma chinensis was studied in the laboratory. Holling Ⅱ function response model and Hassell interference response models were used to determine the predation potential of A.chinensis and the effect of prey density. The results showed that 3rd-5th instar nymphs and adults of predator exhibited Holling Ⅱ functional responses. The sequence of attacking efficiency was as followed: 3rd instar nymph (1.461), 4th instar nymph (1.322), 5th instar nymph (1.086), female adult (0.803) and male adult (0.734). The maximum daily predation capacity of 5th instar nymph was 83.33 individuals. With the increase of preys density, predators searching efficiency decreased gradually, and the searching efficiency of 5th instar nymph was obviously higher than that of other stages. The interference effect of the population density to the predator was significant, and the competition parameter of 3rd instar nymph was the biggest (0.862). It indicated a substantial predation potential, especially the 5th instar nymph of A.chinensis against P.xylostella.
Key words
Arma chinensis; Plutella xylostella; functional response; biological control
小菜蛾Plutella xylostella (Linnaeus)属于鳞翅目,菜蛾科,是一种世界性农业害虫,幼虫主要为害十字花科蔬菜,咬食蔬菜叶片成网状或孔洞状,造成蔬菜减产甚至绝收[1]。长期以来化学农药的大量使用,使小菜蛾的抗药性增强。针对其抗药性,近些年小菜蛾的抗药机理研究较多[2-7],但是研究小菜蛾抗药机理不仅花费的时间长,且不能直接应用于田间。而利用生物防治的方法防治小菜蛾,符合当下“绿色发展”的要求,符合现代人们对绿色食品,健康生活的需求。目前用于防治小菜蛾的天敌多为寄生性昆虫[8-9],小菜蛾捕食性天敌昆虫的研究相对薄弱,国外仅见一篇南美尝试用小菜蛾饲养黑刺益蝽Podisus nigrispinus Dallas的报道,国内只有蜘蛛、微小花蝽Orius minutus Linnaeus、異色瓢虫Harmonia axyridis (Pallas)捕食小菜蛾研究的零星报道[10-13]。笔者发现蠋蝽能取食小菜蛾的各个虫态,具有一定的生防潜力,并对其进行了研究。
蠋蝽Arma chinensis (Fallou)属于半翅目,蝽科,益蝽亚科,蠋蝽属,在我国多省均有分布,它的成虫和若虫均能够捕食鳞翅目、鞘翅目、双翅目等多种害虫的成虫和幼虫,是一种优良的天敌昆虫[14]。张晓军等研究了蠋蝽对榆紫叶甲Ambrostoma quadriimpressum Motschlsky的捕食潜力[15]。目前蠋蝽的人工饲料、大量饲养和定殖行为已经取得明显的进展[16-22],但是田间应用中,针对确定靶标害虫的捕食量数据目前还较为缺乏。
本研究采用捕食功能反应模型[23]、搜寻效应[24]以及天敌自身密度干扰模型[25],在实验室内研究了蠋蝽若虫和成虫对小菜蛾幼虫的捕食作用,以期为确定蠋蝽控制小菜蛾的最佳释放比例,有效控制小菜蛾提供科学依据,通过合理利用天敌昆虫防治小菜蛾,有效降低化学农药对环境和农产品的污染。
1 材料与方法
1.1 供试虫源
蠋蝽和小菜蛾由中国农业科学院植物保护研究所廊坊基地提供。蠋蝽和小菜蛾幼虫均饲养于人工气候箱内,人工气候箱的环境条件设置为:温度(25±1)℃、光照L∥D=14 h∥10 h、相对湿度(70±5)%。
1.2 试验条件
在养虫盒(长21 cm,宽14 cm,高9 cm)内进行蠋蝽对小菜蛾幼虫的捕食功能反应和蠋蝽自身密度干扰反应试验。将养虫盒置于江南RXZ型人工气候箱内,温度(25±1)℃、光照L∥D=16 h∥8 h、相对湿度(70±5)%。
1.3 试验方法
1.3.1 功能反应和搜寻效应试验
在进行捕食功能反应试验前,将蜕皮24 h内的蠋蝽放入容积为300 mL的塑料杯内饥饿24 h,
每杯1头,以纱网覆盖塑料杯,并在纱网上放置一块蘸水的脱脂棉,为蠋蝽提供水分。将饥饿处理后的蠋蝽和小菜蛾4龄幼虫放入养虫盒进行试验,为避免小菜蛾之间相互残杀,在养虫盒内放入一片干净的新鲜甘蓝叶。
捕食功能反应密度设置:蠋蝽3龄若虫和4龄若虫捕食试验设置的小菜蛾密度梯度为5、10、15、20、40头/盒;蠋蝽5龄若虫和成虫试验设置的小菜蛾密度梯度为10、20、40、60、100头/盒。每个密度处理均设置10次重复,24 h后检查并统计小菜蛾幼虫的存活量。
1.3.2 自身密度干扰反应试验
以与捕食功能反应相同的方式对蠋蝽进行饥饿处理,干扰反应设置的蠋蝽密度均为1、2、3、4、5头/盒,小菜蛾4龄幼虫密度为40头/盒和100头/盒,40头/盒提供给蠋蝽3龄若虫和4龄若虫,100头/盒提供给蠋蝽5龄若虫和成虫。每个蠋蝽密度处理均设5次重复,24 h后检查并统计小菜蛾幼虫的存活量。
1.4 数据分析
HollingⅡ功能反应模型:
Na=aNTr/(1+aThN);
搜寻效应方程:S=aTr/(1+aThN);
Hassell的干扰模型:A=aP-b;
其中Na为捕食小菜蛾的头数,a为捕食者对猎物的瞬间攻击率,N是小菜蛾幼虫的密度,Tr是试验的总时间为1 d,Th是处理时间(即捕食者捕食1头猎物所用的时间);Hassell的干扰模型中,P为蠋蝽密度,A为平均每头蠋蝽捕食的小菜蛾幼虫数量,a为天敌的最大捕食量,b为竞争参数。先用Excel 2010处理数据,再利用GraphPad Prism 7拟合方程并作图。
2 结果与分析
2.1 蠋蝽對小菜蛾幼虫的捕食功能反应
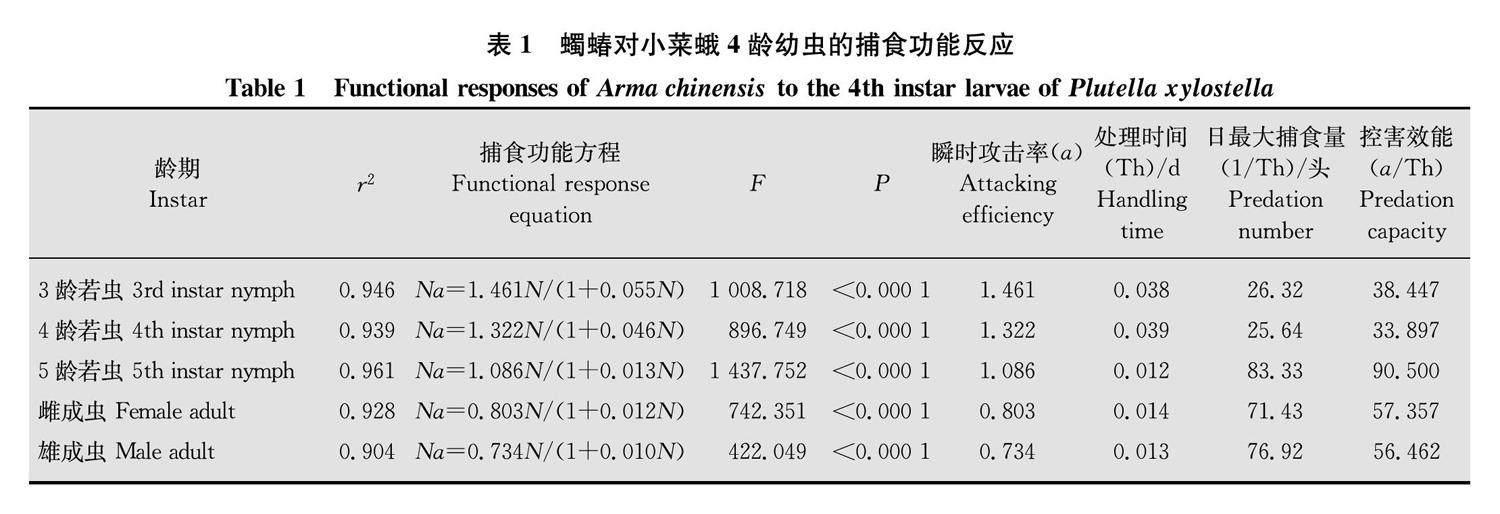
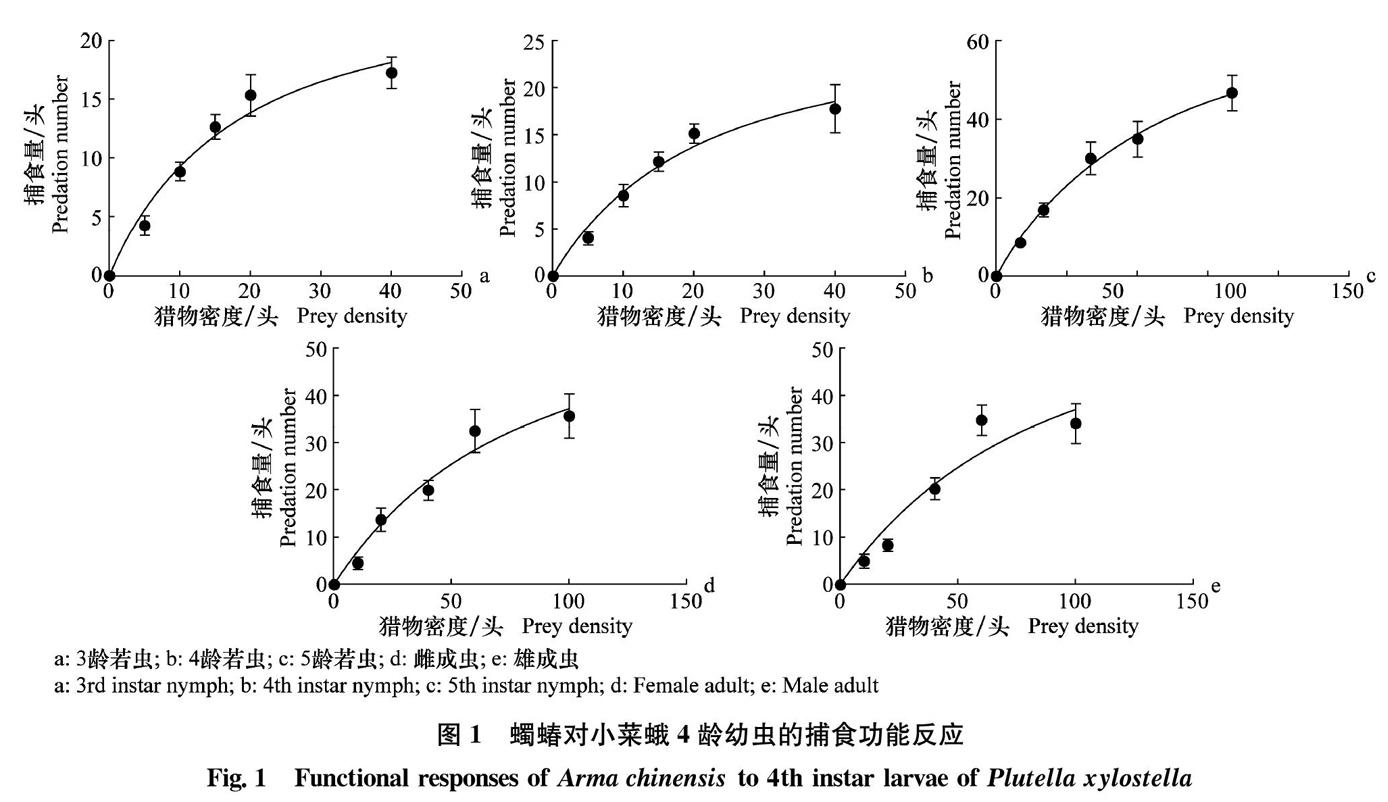
从表1中可以看出,蠋蝽3~5龄若虫和成虫对小菜蛾幼虫的捕食功能反应方程与HollingⅡ模型显著相关(r2>0.9, F >F0.01(1,58)=7.04,P<0.000 1)。蠋蝽5龄若虫处理小菜蛾幼虫的时间最短,只需0.012 d,其日最大捕食量(83.33头)以及控害效能(90.500)均较蠋蝽其他龄期若虫以及雌、雄成虫高。蠋蝽3~5龄若虫和成虫对小菜蛾幼虫的瞬时攻击率大小依次为:3龄若虫(1.461)>4龄若虫(1.322)>5龄若虫(1.086)>雌成虫(0.803)>雄成虫(0.734):从图1中可以看出,随着小菜蛾数量的增加,蠋蝽的捕食量虽然也在增加,但曲线渐渐趋于平缓。
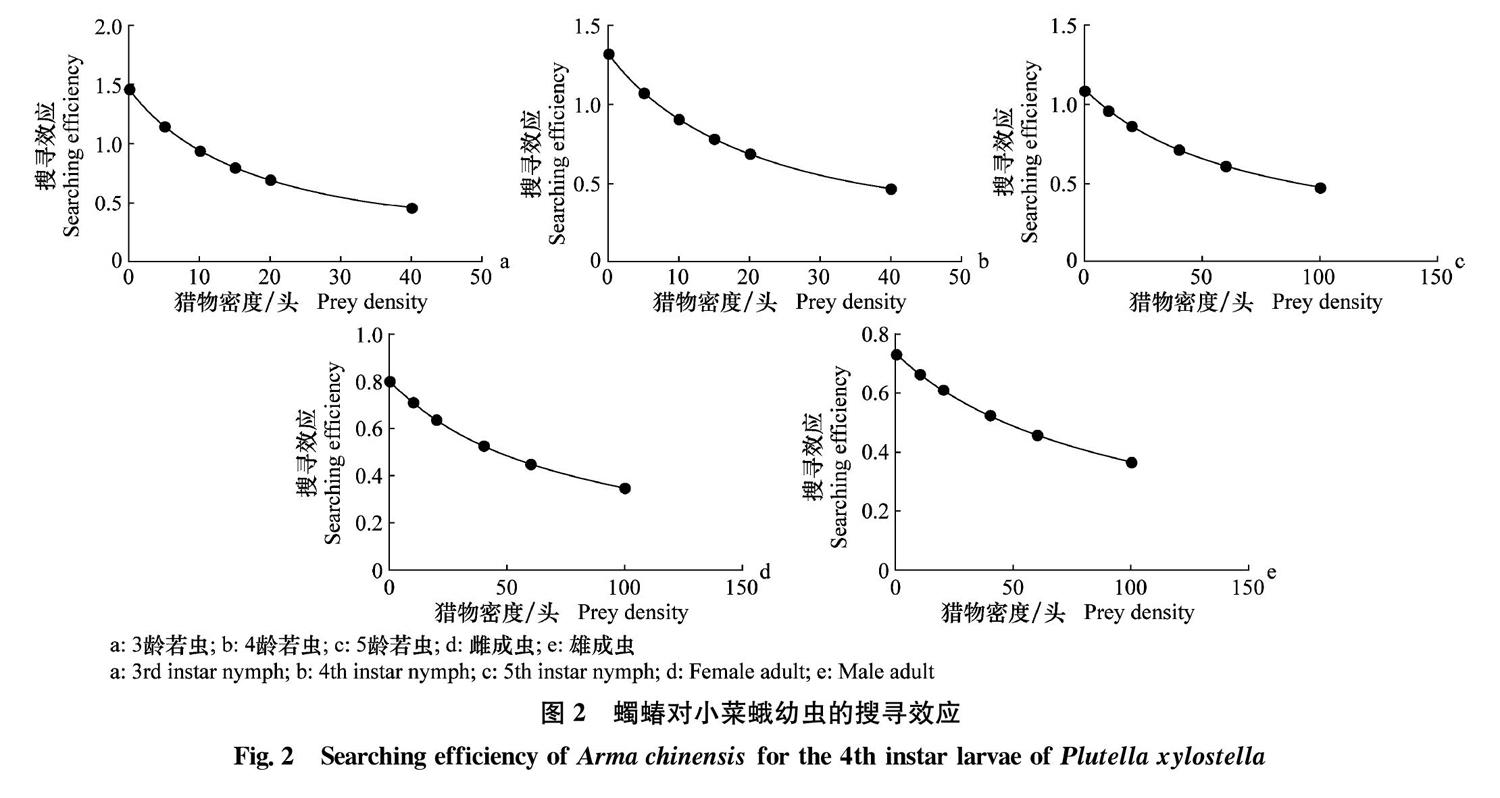
2.2 蠋蝽对小菜蛾幼虫的搜寻效应
蠋蝽对小菜蛾4龄幼虫的搜寻效应数值可根据搜寻效应S=a/(1+aThN)公式计算出,由图2看出,蠋蝽对小菜蛾幼虫的搜寻效应随着小菜蛾幼虫密度的增加而降低。当养虫盒内小菜蛾幼虫的种群密度为40头/盒时,蠋蝽3~5龄若虫和成虫的搜寻效应分别为0.457(3龄若虫)、0.465(4龄若虫)、0.714(5龄若虫)、0.528(雌成虫)、0.524(雄成虫)、蠋蝽3龄若虫的搜寻效应最低,5龄若虫的搜寻效应最高。
2.3 蠋蝽自身密度对捕食量的影响
从表2中可以看出:蠋蝽捕食小菜蛾幼虫存在着捕食者自身的种内干扰作用,随着蠋蝽数量的增加,蠋蝽种内干扰作用逐渐加强,表明蠋蝽自身密度变化与捕食量相关。根据干扰方程可计算出蠋蝽3~5龄若虫和蠋蝽成虫的种内干扰模型,且干扰反应方程与Hassell干扰模型具有显著相关性(r2>0.9,F>F0.01(1, 3)=34.12,P<0.05)。试验数据表明,单头蠋蝽5龄若虫平均可捕食47.06头小菜蛾,具有较大的捕食量。3龄若虫的种内干扰系数最大,b为0.862,而蠋蝽成虫的种内干扰系数相对较小,并且雌成虫的种内干扰小于雄成虫(表3)。
3 讨论
农林中重要的捕食性天敌—蠋蝽,捕食范围广,捕食量大,对小菜蛾等鳞翅目昆虫有着天然的控制力,本试验研究蠋蝽对小菜蛾幼虫的捕食潜力,对充分利用蠋蝽捕食能力,控制小菜蛾数量,科学指导蠋蝽防治小菜蛾具有重要意义。
本研究结果表明,蠋蝽3~5龄若虫和成虫均有较强的捕食能力,捕食功能反应模型与HollingⅡ圆盘方程呈显著相关,这与大多数捕食蝽对猎物的捕食功能反应模型一致[26-31]。试验结果表明:与蠋蝽其他龄期若虫以及成虫相比,蠋蝽5龄若虫对小菜蛾幼虫的处理时间最短,为0.012 d,其日最大捕食量最多,为83.33头,控害效能最高,为90.500。这与已报道的蠋蝽捕食榆紫叶甲卵的研究结果一致,蠋蝽5龄若虫表现出较大的捕食量[15]。
针对天敌昆虫对小菜蛾幼虫的捕食量已有一些报道,拟环纹豹蛛Pardosa psedoannulata(Bsenberg & Strand)对小菜蛾3龄幼虫的最大捕食量为26.2头,三突花蛛Ebrechtella tricuspidata(Fahricius)、沟渠豹蛛P.laura Karsch、星豹蛛P.astrigera L.的最大捕食量分别是12.5、20.9、12.6头[10],微小花蝽对低龄小菜蛾幼虫的日最大捕食量为41.3头[11],异色瓢虫对小菜蛾幼虫的日最大捕食量为50.4头[12],均远低于本试验结果蠋蝽5龄若虫对小菜蛾幼虫的日最大捕食量83.33头。且数据比较没有考虑猎物龄期的差别,本试验中所用猎物为小菜蛾4龄幼虫,而蜘蛛捕食试验用的是小菜蛾3龄幼虫,微小花蝽试验对象是小菜蛾1~2龄幼虫,异色瓢虫试验猎物为小菜蛾2~3龄幼虫,如果考虑猎物个体大小差异,上述3类捕食者对小菜蛾幼虫的捕食量与蠋蝽的数据差别更大。在利用天敌昆虫防治小菜蛾时,蠋蝽比蜘蛛、微小花蝽、异色瓢虫的捕食能力更强,捕食效应更高。
就捕食者的最大捕食能力而言,两种捕食蝽,蠋蝽和微小花蝽表现一致,都是5龄若虫对小菜蛾幼虫的捕食量最大。而异色瓢虫和几种蜘蛛,则表现为成虫对小菜蛾幼虫的捕食量最大:几种蜘蛛的雌蛛取食量大于雄蛛,异色瓢虫的日最大捕食量雄成虫大于雌成虫。
蠋蝽的攻击率表现为若虫比成虫更高,并且低龄若虫(3龄若虫)最高,成虫中雌虫的攻击率高于雄虫。分析原因可能是3龄若虫虽然体积小,但是对于24 h的饥饿处理反应明显,急于捕食,雌性成虫产卵需要预备能量多于雄性。微小花蝽、异色瓢虫和4种蜘蛛不同龄期对小菜蛾幼虫的攻击率也表现出与蠋蝽相同的趋势。微小花蝽5龄若虫、异色瓢虫4龄幼虫、拟环纹豹蛛雌成虫对小菜蛾幼虫的最大攻击率分别为1.007 0、0.708 0、1.121 9,均小于蠋蝽3龄若虫对小菜蛾4龄幼虫的攻击率,进一步说明蠋蝽比蜘蛛、微小花蝽、异色瓢虫的捕食能力更强。
搜寻效应是捕食者在捕食过程中对猎物攻击的一种行为效应[24]。本研究表明随着小菜蛾密度的增大,蠋蝽搜寻效应逐渐降低,与其他捕食者的搜寻效应表现一致[12]。
蠋蝽3~5龄若虫及成虫的搜寻效应以及对小菜蛾平均捕食量,均随着小菜蛾密度的增加而降低,而干扰作用随着天敌蠋蝽密度的上升而增加,蠋蝽自身密度的干扰效应可用Hassell模型(A=aP-b)反映。其中蠋蝽5龄若虫对小菜蛾幼虫的捕食量最大,与上述捕食功能反应表现一致,3龄若虫的自身密度干扰系数最大,b=0.862,可能因为蠋蝽等益蝽亚科昆虫的低龄若虫喜群集取食[32]。成虫的自身密度干扰系数远小于若虫,说明成虫更倾向于单独捕食。
综合考虑捕食量、攻击率、捕食率和搜寻效应,我们发现:蠋蝽3龄若虫对于24 h的饥饿处理反应明显,急于捕食,但同时也因为3龄若虫体积小、对食物需求量比高龄若虫低,因而捕食量并不高;5龄若虫日最大捕食量和控害效能均较其他龄期若虫以及成虫高,并高于已报道的其他几种天敌的数据。在田间释放蠋蝽时,应考虑到种间干扰效应和释放成本,应控制蠋蝽的释放量。
研究捕食者-猎物的关系模型可用于优化生防策略。蠋蝽对小菜蛾的功能反应模型和自身密度干扰模型可以预测蠋蝽在田间应用时的释放比例以及释放虫态,实现将来在田间应用时以最小的释放成本达到最佳的控害效果。然而本试验是在室内恒温条件下进行的,田间应用时存在着气候、温湿度、降雨量等多种不可控制的因素,影响捕食者的捕食能力,因此需进一步评价蠋蝽在田间自然条件下对小菜蛾的控制能力。
参考文献
[1] 吴青君, 张文吉, 朱国仁, 等. 小菜蛾的发生为害特点及抗药性现状[J].中国蔬菜, 2001(5): 49-51.
[2] 梁沛, 高希武, 郑炳宗, 等. 小菜蛾对阿维菌素的抗性机制及交互抗性研究[J]. 农药学学报, 2001, 3(1): 41-45.
[3] 罗万春, 张强. 苦豆子生物碱对小菜蛾体内部分杀虫剂代谢酶活性的影响[J]. 昆虫学报, 2003, 46(1): 122-125.
[4] 梁沛, 夏冰, 石泰, 等. 阿维菌素和高效氯氰菊酯亚致死剂量对小菜蛾谷胱甘肽S-转移酶的影响[J]. 中国农业大学学报, 2003, 8(3): 65-68.
[5] 王树栋, 罗万春, 高兴祥, 等. 曲酸对小菜蛾酚氧化酶抑制作用的研究[J]. 中國农业科学, 2004, 37(9): 1316-1321.
[6] 孙杨, 秦文婧, 黄水金, 等. 小菜蛾对三氟甲吡醚的抗性风险评价与抗性生化机制研究[J]. 应用昆虫学报, 2018, 55(3): 481-488.
[7] XU X X, ZHANG Y Q, FREED S, et al. An anionic defensin from Plutella xylostella with potential activity against Bacillus thuringiensis [J]. Bulletin of Entomological Research, 2016, 106(6): 790-800.
[8] SHI Zuhua, GUO Shijian, LIN Wencai, et al. Evaluation of selective toxicity of five pesticides against Plutella xylostella (Lep:Plutellidae)and their side-effects against Cotesia plutellae (Hym: Braconidae)and Oomyzusso kolowskii (Hym: Eulophidae) [J]. Pest Management Science, 2004, 60(12):1213-1219.
[9] LOHR B, GATHU R, KARIUKI C, et al. Impact of an exotic parasitoid on Plutella xylostella (Lepidoptera: Plutellidae) population dynamics, damage and indigenous natural enemies in Kenya [J]. Bulletin of Entomological Research, 2007, 97(4): 337-350.
[10]全晓宇. 蜘蛛对小菜蛾的捕食作用及其捕食效应的分子检测[D]. 武汉: 湖北大学. 2011.
[11]孙丽娟, 衣维贤, 郑长英. 微小花蝽对小菜蛾捕食控制能力[J]. 应用生态学报, 2017, 28(10): 3403-3408.
[12]陈元洲, 张大友, 张亚, 等. 小菜蛾主要捕食性天敌种类及捕食功能研究[J]. 河南职业技术师范学院学报, 2004, 32(3): 32-34.
[13]ALESSANDRA M V, SERGIO A D B, ROBERTO M G, et al. Comparison of eggs, larvae, and pupae of Plutella xylostella (Lepidoptera: Plutellidae)as prey for Podisus nigrispinus (Hemiptera: Pentatomidae) [J]. Arthropod Biology, 2013, 106(2): 235-242.
[14]邹德玉, 徐维红, 刘佰明. 天敌昆虫蠋蝽的研究进展与展望[J]. 环境昆虫学报, 2016, 38(4): 857-865
[15]张晓军, 张健, 孙守慧. 蠋蝽对榆紫叶甲的捕食作用[J]. 中国森林病虫, 2016, 35(1): 13-15.
[16]潘明真, 张海平, 张长华, 等. 饲养密度和性比对蠋蝽存活和繁殖生物学特性的影响[J]. 中国生物防治学报, 2018, 34(1): 52-58.
[17]李娇娇, 张长华, 易忠经, 等. 三种猎物对蠋蝽生长发育和繁殖的影响[J]. 中国生物防治学报, 2016, 32(5): 552-561.
[18]ZOU Deyu, COUDRON T A, LIU Chenxi, et al. Nutrigenomics in Arma chinensis: transcriptome analysis of Arma chinensis fed on artificial diet and Chinese oaksilk moth Antheraea pernyi pupae [J/OL]. PLoS ONE, 2013, 8(4): e60881.DOI:10.1371/journal.pone.0060881.g001.
[19]宋丽文, 陶万强, 关玲, 等. 不同宿主植物和饲养密度对蠋蝽生长发育和生殖力的影响[J]. 林业科学, 2010, 46(3): 105-110.
[20]廖平, 苗少明, 许若男, 等. 新型蠋蝽若虫液体人工饲料效果评价[J]. 中国生物防治学报, 2019, 35(1): 9-14.
[21]张健, 周毓麟, 孙守慧. 利用人工饲料连代饲养蠋蝽若虫效果评价[J]. 中国森林病虫, 2017, 36(4): 37-40.
[22]高长启, 王志明, 余恩裕.蠋蝽人工饲养技术的研究[J]. 吉林林业料技, 1993(2): 16-18.
[23]HOLLING C S. Some characteristics of simple types of predation and parasitism [J]. The Canadian Entomologist, 1959, 91(7): 385-398.
[24]丁岩钦. 昆虫数学生态学[M]. 北京: 科学出版社, 1994: 257-258, 303-304.
[25]HASSELL M P. A population model for the interaction between Cyzenisal bicans (Fall.) (Tachinidae)and Opero phterabrumata (L.) (Geometridae)at Wytham, Berkshire [J].Journal of Animal Ecology, 1969, 38(3): 567-576.
[26]唐艺婷, 郭义, 何国玮, 等. 不同龄期的益蝽对粘虫的捕食功能反应[J]. 中国生物防治学报, 2018, 34(6): 825-830.
[27]BRBARA D B, FRANCISCO S R, JOS B M, et al. How predation by Podisus nigrispinusis influenced by developmental stage and density of its prey Alabama argillacea [J]. Entomologia Experimentalis et Applicata, 2016, 158: 142-151.
[28]JOS C Z, CARLOS A D, ERALDO R, et al. Predation rate of Spodoptera frugiperda (Lepidoptera:Noctuidae) larvae with and without defense by Podisus nigrispinus (Heteroptera: Pentatomidae) [J]. Brazilian Archives of Biology and Technology, 2008, 51(1): 121-125.
[29]MOHAGHEGH J, CLERCQ P D, TIRRY L. Functional response of the predators Podisus maculiventris(Say)and Podisus nigrispinus (Dallas) (Het. Pentatomidae) to the beet armyworm, Spodoptera exigua (Lep. Noctuidae): effect of temperature [J]. Journal of Application Entomology, 2001, 125(3): 131-134.
[30]李文華, 贾彩娟, 陈惠平, 等. 叉角厉蝽对青野螟幼虫的捕食功能反应[J]. 环境昆虫学报, 2015, 37(4): 843-848.
[31]陈然, 梁广文, 张拯研, 等. 叉角厉蝽对斜纹夜蛾的捕食功能反应[J]. 环境昆虫学报, 2015, 37(2): 401-406.
[32]CARL W S, ANTONIO R P. Heteroptera of economic importance [M]. Florida: CRC Press, 2000: 737-769.
(责任编辑:王 音)
