云南草地贪夜蛾扩散区与首发地种群肠道细菌群落组成结构及多样性的比较
2020-08-25徐天梅符成悦苏造堂肖关丽李维薇陈斌
徐天梅 符成悦 苏造堂 肖关丽 李维薇 陈斌
摘要 草地貪夜蛾是一种新入侵我国的重大害虫。肠道细菌是昆虫肠道微生物的重要类群,在害虫生理生态功能中发挥着重要作用,为弄清云南省草地贪夜蛾扩散地与入侵地种群肠道细菌群落组成结构及多样性的差异,本研究采用Illumina Hiseq技术对云南草地贪夜蛾首发地普洱市宝藏镇、扩散区昆明市嵩明县两个地区草地贪夜蛾4龄幼虫肠道细菌的16S rDNA V3-V4变异区域序列进行了测定分析,应用USEARCH和QIIME软件统计样品序列数和操作分类单元(operational taxonomic unit,OTU)数量分析了肠道细菌群落组成结构及物种丰度与多样性。结果表明,来自普洱市宝藏镇、昆明市嵩明县的6个草地贪夜蛾样品共产生了302 786条有效序列,在97%相似度下可将其聚成564个OTUs,共发现细菌20门、39纲、86目、161科、318属、344种。其中宝藏镇草地贪夜蛾种群的肠道细菌分属于16门、29纲、60目、102科、178属共192种;优势门、纲、目、科、属、种分别为:厚壁菌门(35.09%)、梭菌纲(15.84%)、梭菌目(15.84%)、艾克曼菌科(12.80%)、艾克曼菌属(12.80%)、不可培养的艾克曼菌属细菌(12.80%)。嵩明县草地贪夜蛾肠道细菌共184种,分属于15门、27纲、54目、96科、172属;优势门、纲、目、科、属、种分别为:厚壁菌门(65.87%)、杆菌纲(48.07%)、乳杆菌目(47.32%)、肠球菌科(46.99%)、肠球菌属(46.99%)、NLAE-zl-C485(46.96%)。宝藏镇草地贪夜蛾4龄幼虫肠道内细菌群落Shannon、ACE和Chao1多样性指数分别为3.46、360.25和370.54,分别高于扩散地嵩明县种群的2.26、295.54和305.42;而Simpson指数(0.13)则低于嵩明县种群(0.36)。通过KEGG代谢途径差异分析发现,草地贪夜蛾肠道细菌基因主要参与的代谢途径是整体组织构建和碳水化合物代谢,且宝藏镇草地贪夜蛾肠道细菌基因在大多数的代谢通路中表达量高于嵩明县种群。以上结果说明,云南省草地贪夜蛾首发地种群肠道内细菌群落组成及多样性高于扩散区种群。本研究将为进一步研究不同地区草地贪夜蛾种群生物学差异及肠道微生物对其生态适应性的调控提供理论依据。
关键词 草地贪夜蛾; 肠道细菌; 16S rDNA; 基因测序
中图分类号:
S 435.1
文献标识码: A
DOI: 10.16688/j.zwbh.2020083
Composition and diversity of gut bacteria community of Spodoptera frugiperda from the first invasion site and the dispersal area in Yunnan province
XU Tianmei1, FU Chengyue1, SU Zaotang1, XIAO Guanli2*, LI Weiwei1,3, CHEN Bin1*
(1. College of Plant Protection, Yunnan Agricultural University, Kunming 650201, China;
2.College of Agriculture & Biology Technology, Yunnan Agricultural University, Kunming 650201, China;
3. Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming 650223, China)
Abstract
Fall armyworm, Spodoptera frugiperda, is a serious invaded pest insect of corn, it was firstly found in Baozang town, Yunnan province, China, in 2019. Gut bacteria played important role in the physiological and ecological function. To understand the difference of composition and diversity of gut bacteria communities in S.frugiperda from the first invasion site and the dispersal area, a comparable study on the intestinal bacteria of 4th instar larvae from Baozang and Songming populations of S.frugiperda was conducted using 16S rDNA V3-V4 variation region sequence based on the Illumina Hiseq technique. The sequence numbers of each sample and the numbers of operational taxonomic unit (OTU) were analyzed using the software of USEARCH and QIIME, and the species composition, abundance and diversity indexes of gut bacteria were determined. A total of 302 786 effective sequences were generated from six samples from two regions, which was aggregated into 564 OTUs under 97% similarity. A total of 20 phylum, 39 classes, 86 orders, 161 families, 318 genera and 344 species bacterial were annotated. For Baozang population, a total of 192 species which belong to 178 genus, 102 families, 60 orders, 29 classes and 16 phylum were identified, among which dominant phylum, class, order, family, genera and species were Firmicutes (35.09%), Clostridia (15.84%), Clostridiales (15.84%), Akkermansiaeae (12.80%), Akkermansia (12.80%) and uncultured Akkermansia bacterium (12.80%), respectively. A total of 184 species which belong to 172 genus, 96 families, 54 orders, 27 classes and 15 phylum were identified in Songming population. Of which, dominant phylum, class, order, family, genera and species were Firmicutes (65.87%), Bacilli (48.07%), Lactobacilliales (47.32%), Enterococcaceae (46.99%), Enterococcus (46.99%)and NLAE-zl-c485 (46.96%), respectively. The diversity indexes of Shannon index (3.46), ACE (360.25)and Chao 1 (370.54) in the Baozang population were higher than those in the Songming population, which were 2.26, 295.54 and 305.42, respectively. But the Simpson index in the Baozang population (0.13) was lower than that in the Songming population (0.36). Analysis of the KEGG metabolism pathway showed that the main metabolism pathway of gut bacterial was the whole tissue construction and carbohydrate metabolism, and the relative expression of gut bacterial genes in Baozang population was higher than that in Songming population. These results showed that the composition and diversity of gut bacterial community of S.frugiperda in the first invasion site was higher than those in the dispersal areas in Yunnan province. This study will provide a theoretical basis for further study on the variance of the biology of different populations and adaptation mechanism meditated by gut microorganisms.
Key words
Spodoptera frugiperda; gut bacteria; 16S rDNA; gene sequencing
草地贪夜蛾Spodoptera frugiperda(J.E.Smith),又名秋黏虫,属鳞翅目Lepidoptera夜蛾科Noctuidae灰翅夜蛾属Spodoptera[1]。该虫具有较强的迁飞能力,繁殖能力强,生活周期短,是一种杂食性害虫,能取食玉米、水稻、高粱和甘蔗等353种植物[2]。该虫源于美洲热带及亚热带地区[3],2016年传入非洲,2018年入侵也门、印度和缅甸等亚洲国家, 2019年1月入侵云南省普洱市江城县宝藏镇[4-5],此后迅速扩散,到2019年10月10日,已在我国26个省1 518个县发生,为害的作物达15种。草地贪夜蛾主要寄主为玉米,其次为甘蔗、高粱、谷子、小麦等作物,给玉米生产造成了严重影响[6]。
昆虫肠道微生物被认为是昆虫的第二套遗传密码[7],参与昆虫体内许多重要的生理生化反应,如参与合成昆虫所需营养物质[8]、帮助消化大分子物质[9]、介导抗药性[10]、影响交配行为[11]、调控寄主选择[12]和环境适应性[13]等。因此,关于昆虫肠道微生物群落组成结构及多样性的研究是近年来昆虫学研究的热点之一,这些研究为昆虫与其肠道微生物的协同进化及害虫防治提供了理论依据[14-16]。Almeida等[17]和Acevedo等[18]采用分离培养法分别对巴西和美国草地贪夜蛾幼虫肠道细菌群落组成结构进行了研究,发现草地贪夜蛾幼虫肠道细菌种类丰富;唐运林等[19]、顾偌铖等[20]、韦俊宏等[21]对我国重庆巫溪与巫山地区、李青晏等[22]对云南蒙自地区草地贪夜蛾肠道细菌多样性进行了测定分析,明确了当地草地贪夜蛾肠道可培养细菌种类,且发现其种类随地区不同而存在一定差异。云南省昆明市嵩明县为草地贪夜蛾入侵进入云南后的扩散地,该地区草地贪夜蛾种群肠道细菌组成与入侵地普洱市江城县宝藏镇草地贪夜蛾种群是否存在差异还不清楚。本文研究了两地草地贪夜蛾种群幼虫肠道细菌群落组成结构及其多样性,这对于系统研究该虫生物生态学具有重要意义。
目前,对草地贪夜蛾肠道微生物的研究主要是利用传统培养法进行分离、培养和鉴定,唐运林等采用该方法从重庆巫溪和巫山等地区玉米地草地贪夜蛾肠道中分离并鉴定得到5个属的细菌[19-21],然而采用该方法不能鉴定不可培养细菌种类。随着分子生物学技术的发展,16S rDNA基因序列测定、宏基因组测定(描述细菌的潜在功能)和宏转录组测序(描述活性基因的表达)[23]等技术为系统研究微生物提供了技术支撑,其中16S rDNA是一种重要的分子标记技术,通过获得待测微生物的16S rDNA序列,与公共数据库中已经研究的物种序列进行比较[24]实现种类鉴定。其他一些常用的分子标记则不具有16S rDNA基因的这种分类优势[25],因此,16S rDNA基因分析已成为微生物多样性调查和分类鉴定中的主要技术。由此,本研究基于16S rDNA分析云南草地贪夜蛾扩散区与首发地种群肠道内细菌群落组成结构与多样性及其差异,为进一步研究不同地区草地贪夜蛾种群生物学差异及肠道微生物调控其对生态适应性提供理论依据。
1 材料与方法
1.1 试验材料
供试昆虫为2019年6月20日分别从云南省普洱市江城县宝藏镇(22°41′10″N,101°38′30″E)、昆明市嵩明县(25°15′52″N,102°56′24″E)采集的草地贪夜蛾4龄幼虫。
1.2 肠道解剖
从两个取样点各选择健康的草地贪夜蛾4龄幼虫30头,饥饿24 h,然后于75%乙醇中表面消毒5 min,再用蒸馏水漂洗3次,在超净工作台上解剖,将解剖出的肠道内容物分别溶解于2 mL无菌水中[26]。每个地点设置3个样本,每个样本10头虫,共6个样本。肠道解剖完成后在-20℃冰箱中保存备用。
1.3 肠道细菌总DNA提取
使用PowerSoil DNA Isolation kit提取肠道DNA。
1.4 PCR扩增肠道细菌的16S rDNA
16S rDNA的V3-V4区域作为扩增和测序的目的片段[27],引物由北京百迈科生物科技有限公司设计。引物序列为:515F: 5′-ACTCCTACGGGAGGCAGCA-3′,806R: 5′-GGACTACHVGGGTWTCTAAT-3′。PCR体系为(20 μL):基因组DNA 50 ng±10 ng,上下游引物各0.6 μL,KOD FX Neo Buffer 10 μL,dNTPs(2 mmol/L each)4 μL,KOD FX Neo 0.4 μL,ddH2O補至20 μL。反应条件为:95℃预变性5 min;95℃变性30 s,50℃退火30 s,72℃延伸40 s,20个循环;72℃延伸7 min。PCR产物进行磁珠纯化,样品与磁珠按照1∶1.5混匀后进行磁珠片段筛选,最终用17 μL无菌水洗脱,再进行1次Solexa PCR,反应体系为(20 μL):目的区域PCR纯化产物5 μL,★MPPI-a(2 μmol/L)2.5 μL,★MPPI-b(2 μmol/L)2.5 μL,2×Q5 HF MM 10 μL。反应条件为:98℃预变性30 s;98℃变性10 s,65℃退火30 s,72℃延伸30 s,10个循环;72℃延伸5 min[28]。PCR产物用1.8%的琼脂糖凝胶电泳,电压120 V,电泳40 min,回收目的片段。
1.5 文库构建及数据处理
基于Illumina HiSeq测序平台,利用双末端测序(paired-end)的方法,构建小片段文库并进行测序。对原始数据进行拼接[29],将拼接得到的序列进行质量过滤[30],并去除嵌合体[31],得到高质量的tags序列。数据分析主要使用R语言。
1.6 肠道微生物鉴定和多样性分析
使用Usearch软件(v 7.0)对tags在97%的相似度水平下进行聚类、获得OTU,并基于Silva(细菌)和UNITE(真菌)分类学数据库对OTU进行分类学注释。利用QIIME(v 1.80)软件计算Chao1指数、ACE指数、Shannon指数、Simpson指数。Chao1指数、ACE指数反映样品中群落丰富度,而Shannon指数反映群落的多样性,Simpson指数反映群落中优势种的集中程度,Chao1指数、ACE指数、Shannon指数越大,Simpson指数越小说明样品中的物种越丰富多样。使用R语言分析样品内物种丰富度和多样性信息[32],并通过KEGG代谢途径差异分析宝藏镇和嵩明县的草地贪夜蛾肠道细菌基因在不同代谢通路中的表达情况。
2 结果与分析
2.1 序列拼接和组装
通过Illumina HiSeq测序后,6个样品共获各302 786条有效tags(表1),6个样本共获得20门39纲86目161科318属344种细菌,在97%的相似度水平下聚类共获得564个OTUs。其中宝藏镇有141 653条有效tags,聚类共获得465个OTUs,共注释到细菌16门29纲60目102科178属192种;嵩明县有176 551条有效tags,聚类共获得490个OTUs,共注释到细菌15门27纲54目96科172属184种。确定各样品中的OTU数目以后,对各样品中共有OTU数目进行分析,发现宝藏镇与嵩明县样品之间共有的OTU数目为391个。
2.2 草地贪夜蛾肠道细菌多样性及丰富度分析
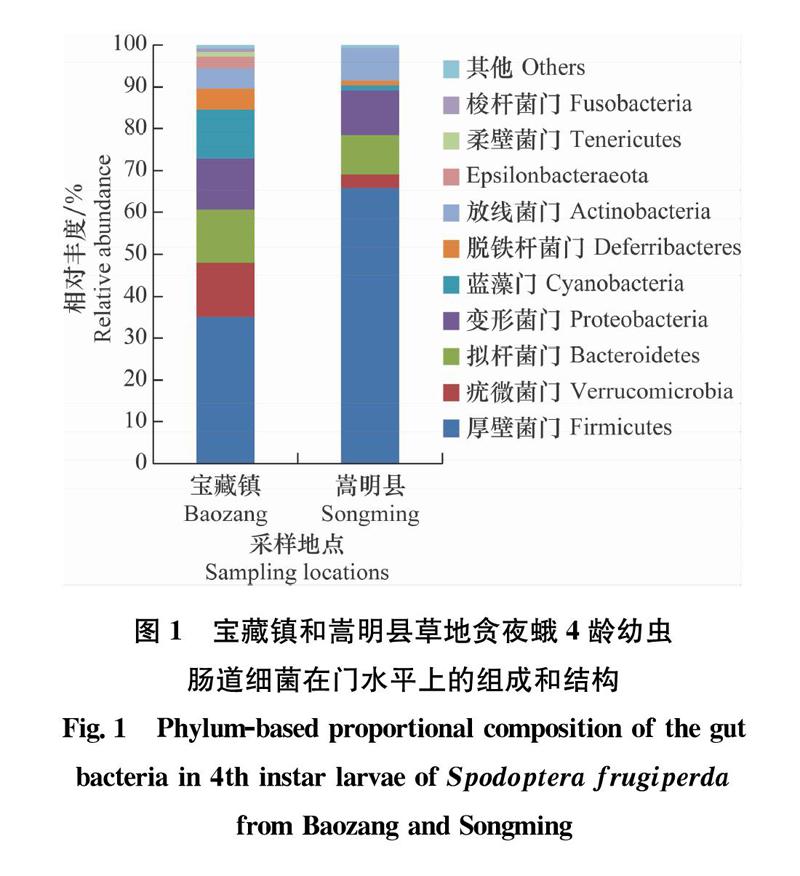
从门的分类水平上看(图1),首发地宝藏镇和扩散区嵩明县草地贪夜蛾4龄幼虫肠道细菌的16S rDNA基因序列均注释到了梭杆菌门Fusobacteria、柔壁菌门Tenericutes、Epsilonbacteraeota、脱铁杆菌门Deferribacteres、疣微菌门Verrucomicrobia、放线菌门Actinobacteria、拟杆菌门Bacteroidetes、蓝藻门Cyanobacteria、变形菌门Proteobacteria和厚壁菌门Firmicutes等共20门。其中,宝藏镇草地贪夜蛾4龄幼虫肠道细菌共注释到16门,丰度前10的门依次为厚壁菌门(35.09%)>疣微菌门(12.80%)>拟杆菌门(12.76%)>变形菌门(12.21%)>蓝藻门(11.71)>脱铁杆菌门(5.9%)>放线菌门(4.76%)>Epsilonbacteraeota(2.66%)>柔壁菌门(1.10%)>梭杆菌门(0.86%);扩散区嵩明县草地贪夜蛾肠道细菌的16S rDNA基因序列共注释到15门,丰度前10的门依次为厚壁菌门(65.87%)>变形菌门(10.73%)>拟杆菌门(9.32%)>放线菌门(7.72%)>疣微菌门(3.29%)>蓝藻门(1.22%)>脱铁杆菌门(1.04%)>梭杆菌门(0.06%)>柔壁菌门(0.03%)>Epsilonbacteraeota(0.02%)。由此可以看出,在门的水平上,宝藏镇和嵩明县草地贪夜蛾4龄幼虫肠道内优势细菌门组成不同,但二者的优势菌门均包括厚壁菌门,不过该门在两地的相对丰度差异较大,宝藏镇和嵩明县草地贪夜蛾幼虫肠道细菌中厚壁菌门的相对丰度分别为35.09%和65.87%。
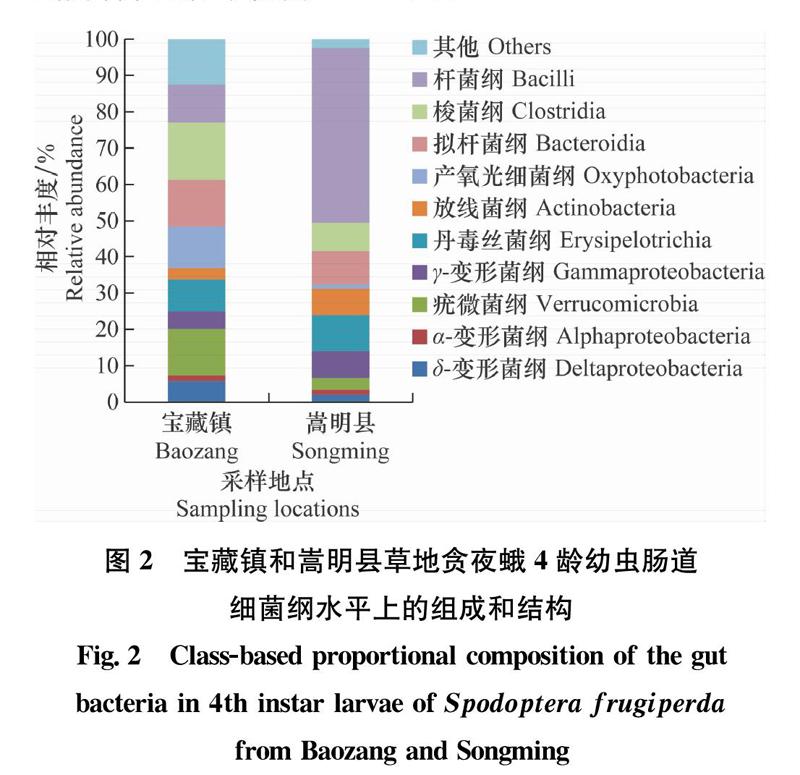
从纲水平来看(图2),宝藏镇和嵩明县草地贪夜蛾4龄幼虫肠道细菌16S rDNA基因序列均注释到了δ-变形菌纲Deltaproteobacteria、α-变形菌纲Alphaproteobacteria、疣微菌纲Verrucomicrobia、γ-变形菌纲Gammaproteobacteria、丹毒丝菌纲Erysipelotrichia、放线菌纲Actinobacteria、产氧光细菌纲Oxyphotobacteria、拟杆菌纲Bacteroidia、梭菌纲Clostridia、杆菌纲Bacilli等共计39纲。其中宝藏镇草地贪夜蛾肠道细菌的16S rDNA基因序列共注释到29纲,丰度前10的纲依次为梭菌纲(15.84%)>疣微菌纲(12.80%)>拟杆菌纲(12.76%)>产氧光细菌纲(11.69%)>杆菌纲(10.44%)>丹毒丝菌纲(8.73%)>δ-变形菌纲(5.71%)>γ-变形菌纲(4.84%)>放线菌纲(3.07%)>α-变形菌纲(1.66%);嵩明县草地贪夜蛾肠道细菌共注释到27个纲,丰度前10的纲为:杆菌纲(48.07%)>丹毒丝菌纲(9.94%)>拟杆菌纲(9.32%)>梭菌纲(7.81%)>γ-变形菌纲(7.35%)>放线菌纲(7.21%)>疣微菌纲(3.28%)>δ-变形菌纲(2.05%)>α-变形菌纲(1.34%)>产氧光细菌纲(1.18%)。由此表明,首发地宝藏镇草地贪夜蛾的肠道细菌优势纲为梭菌纲Clostridia,丰度为15.84%;而扩散地嵩明县草地贪夜蛾肠道细菌优势纲为杆菌纲,丰度为48.07%。
从目水平来看(图3),宝藏镇和嵩明县草地贪夜蛾4龄幼虫肠道细菌的16S rDNA基因序列均注释到了棒状菌目Corynebacteriales、脫硫弧菌目Desulfovibrionales、疣微菌目Verrucomirobiales、微球菌目Micrococcales、肠杆菌目Enterobacteriales、丹毒丝菌目Erysipelotrichales、Chloroplast、拟杆菌目Bacteroidales、梭菌目Clostridiales、乳杆菌目Lactobacillales等共计86目。其中宝藏镇草地贪夜蛾肠道细菌的16S rDNA基因序列共注释到60目,丰度前10的目依次为梭菌目(15.84%)>疣微菌目(12.80%)>拟杆菌目(11.69%)>Chloroplast(11.68%)>丹毒丝菌目(8.73%)>乳杆菌目(8.69%)>脱硫弧菌目(5.67%)>肠杆菌目(1.23%)>微球菌目(0.39%)>棒状菌目(0.37%);嵩明县草地贪夜蛾肠道细菌共注释到54个目,丰度前10的目依次为乳杆菌目(47.32%)>丹毒丝菌目(9.94%)>拟杆菌目(9.17%)>梭菌目(7.82%)>肠杆菌目(6.30%)>微球菌目(4.19%)>疣微菌目(3.29%)>脱硫弧菌目(2.00%)>棒状菌目(1.97%)>Chloroplast(1.18%)。由此表明,首发地宝藏镇草地贪夜蛾的肠道细菌优势目为梭菌目,丰度为15.84%;扩散地嵩明县草地贪夜蛾肠道细菌优势目为乳杆菌目,丰度为47.32%。
从科水平来看(图4),宝藏镇和嵩明县草地贪夜蛾4龄幼虫肠道细菌的16S rDNA基因序列均注释到了拟杆菌科Bacteroidaceae、瘤胃菌科Ruminococcaceae、短杆菌科Brevibacteriaceae、艾克曼菌科Akkermansiaeae、鼠杆菌科Muribaculaceae、肠杆菌科Enterobacteriaceae、毛螺菌科Lachnospiraceae、丹毒丝菌科Erysipelotrichaceae、微球菌科Micrococcaceae、肠球菌科Enterococcaceae等共计161科。其中宝藏镇草地贪夜蛾肠道细菌的16S rDNA基因序列共注释到102科,丰度前10的科依次为艾克曼菌科(12.80%)>微球菌科(11.63%)>鼠杆菌科(9.41%)>丹毒丝菌科(8.73%)>毛螺菌科(8.21%)>瘤胃菌科(6.77%)>肠球菌科(1.31%)>肠杆菌科(1.23%)>拟杆菌科(0.21%)>短杆菌科(0.06%);嵩明县草地贪夜蛾肠道细菌共注释到96个科,丰度前10的科为肠球菌科(46.99%)>丹毒丝菌科(9.94%)>肠杆菌科(6.30%)>毛螺菌科(5.34%)>鼠杆菌科(5.05%)>艾克曼菌科(3.29%)>短杆菌科(3.08%)>瘤胃菌科(1.99%)>拟杆菌科(1.25%)>微球菌科(1.02%)。由此表明,首发地宝藏镇草地贪夜蛾的肠道细菌优势科菌为艾克曼菌科,丰度为12.80%;扩散地嵩明县草地贪夜蛾肠道细菌优势科菌为肠球菌科,丰度为46.99%。
从属水平上看(图5),宝藏镇和嵩明县草地贪夜蛾4龄幼虫肠道细菌的16S rDNA基因序列均注释到了黏液杆菌属Mucispirillum、异杆菌属Allobaculum、ZOR0006、拟杆菌属Bacteroides、短杆菌属Brevibacterium、肠杆菌属Enterobacter、艾克曼菌属Akkermansia、Muribaculaceae科的一未知属、节杆菌属Arthrobacter、肠球菌属Enterococcus等共计318属。宝藏镇草地贪夜蛾4龄幼虫肠道细菌共注释到178属,丰度前10的属依次为艾克曼菌属(12.80%)>节杆菌属(11.63%)>Muribaculaceae科的一未知属(9.30%)>黏液杆菌属(5.10%)>异杆菌属(1.82%)>肠球菌属(1.31%)>ZOR0006(0.40%)>拟杆菌属(0.21%)>肠杆菌屬(0.14%)>短杆菌属(0.05%);嵩明县草地贪夜蛾4龄幼虫肠道内细菌共注释到172个属,丰度前10的属依次为肠球菌属(46.99%)>肠杆菌属(6.20%)>异杆菌属(5.60%)>黏液杆菌属(4.99%)>艾克曼菌属(3.29%)>短杆菌属(3.08%)>拟杆菌属(1.25%)>Muribaculaceae科的一未知属(1.04%)>节杆菌属(1.02%)>ZOR0006(0.06%)。由此表明,首发地宝藏镇的草地贪夜蛾肠道优势属菌为艾克曼菌属,丰度为12.80%;扩散地嵩明县的草地贪夜蛾肠道优势属菌为肠球菌属,丰度为46.99%。
从种水平上看(图6),宝藏镇和嵩明县草地贪夜蛾4龄幼虫肠道细菌的16S rDNA基因序列均注释到了沙德勒氏黏菌Mucispirillum schaedleri、不可培养的异杆菌属细菌uncultured Allobaculum bacterium、细菌ZOR006、不可培养的拟杆菌属细菌uncultured Bacteroides bacterium、短杆菌NXYK Brevibacterium sp. NXYK、细菌HA01、不可培养的艾克曼菌属细菌uncultured Akkermansia bacterium、不可培养的鼠杆菌科一未知属细菌uncultured Muribaculaceae bacterium、烟草节杆菌Arthrobacter nicotianae以及细菌NLAE-zl-C485等344个种。宝藏镇草地贪夜蛾4龄幼虫肠道细菌共注释到192个种,其中丰度前10的种依次为不可培养的艾克曼菌属细菌(12.80%)>烟草节杆菌(11.63%)>鼠杆菌科一未知属细菌(9.30%)>沙德勒氏黏菌(5.10%)>不可培养的异杆菌属细菌(1.28%)>细菌NLAE-zl-C485(0.94%)>不可培养的拟杆菌属细菌(0.16%)>细菌HA01(0.14%)>短杆菌NXYK(0.06%)>细菌ZOR006(0.04%);嵩明县草地贪夜蛾幼虫肠道内细菌共注释到184种,丰度前10的种依次为细菌NLAE-zl-C485(46.96%)>细菌HA01(6.20%)>不可培养的异杆菌属细菌(5.60%)>鼠杆菌科一未知属细菌(4.99%)>不可培养的艾克曼菌属细菌(3.28%)>短杆菌NXYK(3.07%)>不可培养的拟杆菌属细菌(1.25%)>沙德勒氏黏菌(1.04%)>烟草节杆菌(1.02%)>细菌ZOR006(0.06%)。由此表明,首发地宝藏镇草地贪夜蛾肠道优势菌为不可培养的艾克曼菌属细菌,丰度为12.80%;扩散地嵩明县草地贪夜蛾肠道优势种为细菌NLAE-zl-C485,丰富度为46.96%。
2.3 草地贪夜蛾肠道细菌群落多样性分析
由表2可知,草地贪夜蛾幼虫肠道细菌有较高的丰富度和多样性,宝藏镇和嵩明县的草地贪夜蛾肠道细菌ACE指数分别为360.25和295.54,Chao1指数分别为370.54和305.42,Simpson指数分别为0.13和0.36,Shannon指数分别为3.46和2.26。宝藏镇的草地贪夜蛾肠道细菌ACE指数、Chao1指数、Shannon指数均高于嵩明县,说明宝藏镇的草地贪夜蛾肠道细菌多样性更高,而宝藏镇的Simpson指数低于嵩明县,说明宝藏镇草地贪夜蛾肠道细菌的优势种集中程度高于嵩明县。
2.4 功能预测及分析
通过KEGG代谢途径差异分析(图7),无论是首发地宝藏镇,还是扩散区的嵩明县,草地贪夜蛾幼虫肠道细菌所富集到的代谢通路一致,主要富集到的代谢通路为信号传导、膜运输、整体组织构建、翻译、氨基酸代谢、核苷酸代谢、能量代谢、辅助因子和维生素代谢、复制和修复、碳水化合物代谢。但在部分代謝通路中基因表达存在差异,从信号传导、整体组织构建、翻译、氨基酸代谢、核苷酸代谢、能量代谢、辅助因子和维生素代谢、复制和修复通路基因表达来看,首发地BZ601样品中分离出的肠道细菌基因表达量高于扩散地Sm601样品;而在膜运输、碳水化合物代谢通路中,首发地BZ601样品中分离出的肠道细菌基因表达量低于扩散地Sm601样品。
3 讨论
从当前报道的草地贪夜蛾肠道可培养细菌组成来看,各地存在一定差异,其中巴西草地贪夜蛾种群肠道分离获得的细菌有肠球菌属、代尔夫特菌属、勒克氏菌属、细杆菌属、假单胞菌属、节杆菌属和葡萄球菌属7属[17],美国草地贪夜蛾种群中分离的细菌有泛菌属、肠杆菌属、拉恩氏菌属、拉乌尔菌属和克雷伯氏菌属5属[18],我国重庆种群分离的肠道细菌有克雷伯氏菌属、不动杆菌属、假单胞菌属、肠杆菌属和气单胞菌属5属[19],云南省蒙自地区的草地贪夜蛾幼虫肠道分离到的细菌有克雷伯氏菌属、肠球菌属、沙雷氏菌属和摩根菌属4属[22]。本研究通过16S rDNA和Illumina Hiseq技术研究发现云南省草地贪夜蛾首发地种群与扩散地种群肠道内细菌共计20门39纲86目161科318属344种,肠道细菌种类显著多于美国和巴西草地贪夜蛾的肠道微生物类群,也多于重庆和云南蒙自地区草地贪夜蛾肠道细菌类群,究其原因,可能是不同地域环境条件及寄主植物的差异造成,也有可能是不同区域内施用的农药等化学物质不同造成,这些都值得进一步深入研究。从肠道细菌的优势属组成来看,本研究中首发地和扩散地草地贪夜蛾幼虫肠道内细菌与唐运林的报道存在差异,产生差异的原因在于16S rDNA和Illumina Hiseq技术不仅可以获得可以培养的细菌,还可以检测到许多不可培养的细菌,相对于传统的分离培养技术,16S rDNA和Illumina Hiseq技术能够更加全面地揭示肠道微生物组成结构及多样性。
从草地贪夜蛾幼虫肠道细菌类群组成来看,首发地与扩散地存在明显差异,其中宝藏镇草地贪夜蛾种群肠道内细菌隶属于16门29纲60目102科178属192种,嵩明县种群肠道内细菌隶属于15门27纲54目96科172属184种。宝藏镇和嵩明县两个地区草地贪夜蛾肠道细菌的优势菌门均为厚壁菌门,但扩散地优势菌门的相对丰度显著高于首发地,而在其他分类水平上,两个地方的菌群组成及优势菌种都存在差异,其原因可能是由于草地贪夜蛾迁飞扩散进入另一个生态环境后,两个地方的生态环境及作物组成结构不同。此外,本研究通过多个生物多样性指数分析,发现宝藏镇的草地贪夜蛾肠道细菌丰富度高于嵩明县。由此可以看出,首发地宝藏镇田间草地贪夜蛾肠道细菌种类较扩散地的嵩明县丰富,原因可能是两地不同的气候环境或两地点寄主食物有差异,还有待进一步探讨。本研究也表明,云南草地贪夜蛾首发地和扩散地种群肠道中细菌丰度前10的细菌门主要有梭杆菌门、柔壁菌门、Epsilonbacteraeota、脱铁杆菌门、疣微菌门、放线菌门、拟杆菌门、蓝藻门、变形菌门和厚壁菌门。同时,厚壁菌门细菌在稻纵卷叶螟Cnaphalocrocis medinalis(Guenée)、棉铃虫Helicoverpa armigera(Hübner)、小菜蛾Plutella xylostella(Linnaeus)、家蚕Bombyx mori Linnaeus、斜纹夜蛾 S.litura (Fabricius)等昆虫肠道内都被分离获得[32-33],说明厚壁菌门在昆虫肠道内广泛存在。此外,唐运林等运用传统培养法研究发现重庆地区草地贪夜蛾的肠道细菌属于变形菌门、厚壁菌门和放线菌门[19],这3个门的细菌在本研究的云南省草地贪夜蛾首发地和扩散地种群肠道中也存在,进一步表明16S rDNA和Illumina Hiseq技术可用于昆虫肠道内可培养和不可培养细菌群落组成结构及多样性分析。
昆虫肠道细菌在昆虫消化食物、抗逆和生殖调控中发挥着重要作用[18,34]。据报道,草地贪夜蛾肠道中存在着大量的可降解农药的细菌[20],而本研究中对于草地贪夜蛾肠道内具有降解农药作用细菌还未做深入研究,这也将是下一步深入研究的重要内容。此外,本研究通过KEGG代谢途径差异分析,发现在多数代谢通路中,宝藏镇草地贪夜蛾肠道细菌的基因表达量高于嵩明县,仅在膜运输、碳水化合物代谢通路中,宝藏镇低于嵩明县。造成该差异的原因可能是由于宝藏镇与嵩明县地理位置及气候差异,草地贪夜蛾为适应不同的环境产生了适应性变化。
参考文献
[1] 郭井菲, 赵建周, 何康来, 等. 警惕危险性害虫草地贪夜蛾入侵中国[J]. 植物保护, 2018, 44(6): 1-10.
[2] MONTEZANO D G, SPECHT A, SOSA-GMEZ D R, et al. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas [J]. African Entomology, 2018, 26(2): 286-300.
[3] SPARKS A N. A review of the biology of the fall armyworm [J]. The Florida Entomologist, 1979, 62(2): 82-87.
[4] 杨学礼, 刘永昌, 罗茗钟, 等. 云南省江城县首次发现迁入我国西南地区的草地贪夜蛾[J]. 云南农业, 2019, 34(1): 72.
[5] 姜玉英, 刘杰, 朱晓明. 草地贪夜蛾侵入我国的发生动态和未来趋势分析[J]. 中国植保导刊, 2019, 39(2): 33-35.
[6] 姜玉英, 刘杰, 谢茂昌, 等. 2019年我国草地贪夜蛾扩散为害规律观测[J]. 植物保护, 2019, 45(6): 10-19.
[7] ZILBER-ROSENBERG I, ROSENBERG E. Role of microorganisms in the evolution of animals and plants: the hologenome theory of evolution [J]. FEMS Microbiology Reviews, 2008, 32(5): 723-735.
[8] SALEM H, BAUER E, STRAUSS A S, et al. Vitamin supplementation by gut symbionts ensures metabolic homeostasis in an insect host [J/OL]. Proceedings of the Royal Society B: Biological Sciences, 2014, 281(1796): 20141838. DOI: 10.1098/rspb.2014.1838.
[9] LUNDGREN J G, LEHMAN R M. Bacterial gut symbionts contribute to seed digestion in an omnivorous beetle [J/OL]. PLoS ONE, 2010, 5(5): e10831. DOI: 10.1371/journal.pone.0010831.
[10]MANSOURI S M, GANBALANI G N, FATHI S A A, et al. Nutritional indices and midgut enzymatic activity of Phthorimaea operculella (Lepidoptera: Gelechiidae) larvae fed different potato germplasms [J]. Journal of Economic Entomology, 2013, 106(2): 1018-1024.
[11]SHARON G, SEGAL D, RINGO J M, et al. Commensal bacteria play a role in mating preference of Drosophila melanogaster [J]. Proceedings of the National Academy of Sciences, 2010, 107(46): 20051-20056.
[12]MCLEAN A H C, VAN ASCH M, FERRARI J, et al. Effects of bacterial secondary symbionts on host plant use in pea aphids [J]. Proceedings of the Royal Society B: Biological Sciences, 2010, 278(1706): 760-766.
[13]TSUCHIDA T, KOGA R, HORIKAWA M, et al. Symbiotic bacterium modifies aphid body color [J]. Science, 2010, 330(6007): 1102-1104.
[14]魯迎新, 刘彦群, 李群, 等. 昆虫肠道微生物多样性研究进展[J]. 河南农业科学, 2016, 45(11): 1-7.
[15]张振宇, 圣平, 黄胜威, 等. 昆虫肠道微生物的多样性、功能及应用[J]. 生物资源, 2017, 39(4): 231-239.
[16]邴孝利, 陆益佳. 水果害虫斑翅果蝇体内微生物菌群研究进展[J]. 微生物学报, 2019, 59(10): 1880-1888.
[17]ALMEIDA L G, MORAES L A, TRIGO J R, et al. The gut microbiota of insecticide-resistant insects houses insecticide-degrading bacteria: A potential source for biotechnological exploitation [J/OL]. PLoS ONE, 2017, 12(3): e0174754. DOI: 10.1371/journal.pone.0174754.
[18]ACEVEDO F E, PEIFFER M, TAN C W, et al. Fall armyworm-associated gut bacteria modulate plant defense responses [J]. Molecular Plant-Microbe Interactions, 2017, 30(2): 127-137.
[19]唐运林, 吴燕燕, 顾偌铖, 等. 重庆地区草地贪夜蛾肠道细菌的分离鉴定[J]. 西南大学学报(自然科学版), 2019, 41(7): 8-14.
[20]顾偌铖, 唐运林, 吴燕燕, 等. 重庆地区取食高粱的草地贪夜蛾与玉米黏虫肠道细菌比较[J]. 西南大学学报(自然科学版), 2019, 41(8): 6-13.
[21]韦俊宏, 唐运林, 顾偌铖, 等. 重庆地区玉米叶内生细菌的分离及其与草地贪夜蛾肠道菌的比较[J]. 西南大学学报(自然科学版), 2019, 41(9): 17-23.
[22]李青晏, 唐运林, 蒋睿轩, 等. 云南地区草地贪夜蛾肠道细菌的分离及鉴定[J]. 西南大学学报(自然科学版), 2020, 42(1): 1-8.
[23]GOODRICH J K, DI RIENZI S. C, POOLE A. C, et al. Conducting a microbiome study [J]. Cell, 2014, 158(2): 250-262.
[24]McDONALD D, PRICE M N, GOODRICH J, et al. An improved greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea [J]. The ISME Journal, 2012, 6(3): 610-618.
[25]KUCZYNSKI J, STOMBAUGH J, WALTERS W A, et al. Using QIIME to analyse 16S rRNA gene sequences from microbial communities [J/OL]. Current Protocols in Bioinformatics Unit, 2011, 36(1): 10.7.1-10.7.20.DOI:10.1002/0471250953.bi1007s36.
[26]蓝波妙. 斜纹夜蛾肠道细菌多样性及其功能研究[D]. 福州: 福建农林大学, 2016.
[27]FLORES R, SHI Jianxin, YU Guoqin, et al. Collection media and delayed freezing effects on microbial composition of human stool [J]. Microbiome, 2015, 3(1): 1-11.
[28]魏丹峰, 王秀吉, 杨锦, 等. 取食不同食料的美国白蛾幼虫肠道细菌多样性及差异性研究[J]. 环境昆虫学报, 2017, 39(3): 515-524.
[29]MAGOCˇ T, SALZBERG S L. FLASH: fast length adjustment of short reads to improve genome assemblies [J]. Bioinformatics, 2011, 27(21): 2957-2963.
[30]BOLGER A M, LOHSE M, USADEL B. Trimmomatic: a flexible trimmer for Illumina sequence data [J]. Bioinformatics, 2014, 30(15): 2114-2120.
[31]EDGAR R C, HAAS B J, CLEMENTE J C, et al. UCHIME improves sensitivity and speed of chimera detection [J]. Bioinformatics, 2011, 27(16): 2194-2200.
[32]刘小改, 杨亚军, 廖秋菊, 等. 稻纵卷叶螟肠道细菌群落结构与多样性分析[J]. 昆虫学报, 2016, 59(9): 965-976.
[33]孙博通, 蓝波妙, 王倩, 等. 斜纹夜蛾幼蟲肠道细菌分离鉴定及其功能初步分析[J]. 生物资源, 2017, 39(4): 264-271.
[34]SU Qi, ZHOU Xiaomao, ZHANG Youjun. Symbiont-mediated functions in insect hosts [J]. Communicative & Integra Biology, 2013, 6(3): e23804. DOI: 10.4161/cib.23804.
(责任编辑:田 喆)
