青海湖流域植被碳利用效率时空动态研究
2020-08-24兰垚曹生奎曹广超李晓东杨羽帆雷义珍
兰垚, 曹生奎, *, 曹广超, 李晓东, 杨羽帆, 雷义珍
青海湖流域植被碳利用效率时空动态研究
兰垚1, 2, 3, 曹生奎1, 2, 3, *, 曹广超2, 3, 李晓东4, 杨羽帆1, 2, 雷义珍1, 2, 3
1.青海师范大学 地理科学学院,西宁 810008 2. 青海师范大学 青海省自然地理与环境过程重点实验室,西宁 810008 3.青海省人民政府-北京师范大学 高原科学与可持续发展研究院,西宁 810008 4. 青海省气象科学研究所,西宁 810001
植被碳利用效率(CUE)是度量生态系统植被固碳能力和效率的标尺, 对评估生态系统碳储量和揭示碳平衡机理具有重要意义。以青海湖流域为研究对象, 利用MOD17A2H数据估算了2000—2018年青海湖流域植被CUE, 揭示了植被CUE的动态变化特征, 并分析其影响因子。得到如下结果: (1)青海湖流域植被CUE年内变化表现为从3月开始逐渐增加, 6、7月达到最大, 然后减小; 年均植被CUE为0.58, 其值在0.55—0.63之间变化且呈略微下降趋势, 线性递减率为0.01 (10 a)–1; (2)青海湖流域内植被CUE呈现以青海湖为中心“高—低—高”的环带状空间分布特征, 其值在0.54—0.76之间变化(除水域之外), 且平均值为0.58; (3)青海湖流域植被CUE主要受气温和降水影响, 气温对植被CUE的影响较降水明显, 随气温升高和降水增加, 植被CUE呈减小趋势。研究结果可为明确该区碳循环过程中的源/汇问题提供数据支撑, 并为高寒内陆地区植被CUE的研究提供方法借鉴, 同时对理解全球变化和预测生态系统对全球变化的响应具有重要意义。
MODIS; 青海湖流域; 植被; 碳利用效率; 时空变化
0 引言
生态系统植被碳利用效率(carbon use efficiency, CUE)作为植被净初级生产力(net primary production, NPP)与总初级生产力(gross primary production, GPP)的比率[1], 其值大小代表植被固定大气二氧化碳(CO2)的能力, 在一定程度上反映了生态系统植被的固碳潜力和对碳同化能力的强弱, 是决定陆地生态系统碳储量的关键性因子[2-5]。植被CUE也是度量生态系统植被固碳能力和效率的标尺, 植被CUE越高表明生态系统单位固碳的生长转移越高, 生态系统固碳潜力越大[6]。研究和量化生态系统植被CUE可为确定生态系统碳源/汇提供数据支撑, 并对全球变化背景下生态系统响应的研究有重要意义[7-8]。
近年来, 生态系统植被CUE的研究受到国内外诸多碳循环和气候变化等学者的关注, 并开展了大量关于陆地生态系统植被CUE的研究工作[9-12], 但是, 目前对陆地生态系统植被CUE是否为恒定值尚存在较大争议。一种观点认为植被呼吸与GPP成线性同步变化, 且与光合作用同化的含糖量有关, 植被CUE为恒定值[13-14]。另一种观点则认为光合作用和呼吸作用的驱动过程受光合有效辐射、温度和降水等环境因子和物种类型的控制,植被CUE并非恒定值[2,6]。例如, Delucia[8]等通过生物计量法和模型估算法对森林CUE研究表明, 森林CUE的值在0.23—0.83之间波动。目前, 大多数有关植被CUE的研究是利用模型和涡度相关系统测量等方法所进行的, 且以森林、草地等单一的生态系统为主[15,16], 以综合性的生态系统为单位的研究较少, 而关于植被CUE空间变化特征也鲜有报道。据此, 笔者认为造成上述观点分歧的主要原因在于研究方法以及研究对象的单一, 而生态系统植被 CUE的研究更需要通过多种生态系统尺度下的对比性研究才能得到较为可靠的结论。
青藏高原作为中国面积最大、世界海拔最高的高原, 是全球气候变化的敏感区和感应器, 也是维系国内乃至全球生态平衡的重要屏障[17]。青海湖位于青藏高原东北部, 是中国最大的断层陷落型内陆咸水湖[18], 因地处青藏高原高寒区、西北干旱区和东部季风区的交界处, 使得青海湖成为了维系青藏高原东北部生态安全的重要水体[19]。青海湖流域作为青藏高原的重要组成部分, 在区域乃至亚洲都具有重要的生态地位。故此, 研究青海湖流域植被CUE对于区域乃至全球陆地生态系统碳平衡都有重要意义。
本文利用MODIS影像, 分析青海湖流域植被CUE的时空变化特征, 探究青海湖流域植被CUE是否为恒定值。所得结论可为明确该区碳循环过程中的源/汇问题提供数据支撑, 并对高寒内陆地区植被CUE研究提供方法借鉴, 同时对理解全球变化和预测生态系统对全球变化的响应具有重要意义。
1 材料与方法
1.1 研究区概况
青海湖流域位于青藏高原东北部(36°15′— 38°20′ N, 97°50′—101°20′ E), 是一个四面环山的内陆盆地, 流域内西北高而东南低, 海拔在3169— 5268 m之间, 总面积约2.96×104km2[19]。青海湖流域属于典型的高原半干旱高寒气候, 流域内干旱少雨, 风力强劲且蒸发量大, 昼夜温差大, 生长季短, 非生长季长[20], 季节性冻土广泛发育。流域内多年降雨量在290—580 mm左右, 多年平均降雨量为386 mm, 且主要集中在生长季; 流域内生长季主要以东南风为主, 非生长季主要以西风为主, 年均潜在蒸发量在839—912 mm之间; 青海湖流域年均气温在-1.1—4.0 ℃之间, 无霜期约110 d左右[21-22]。青海湖流域水系主要由布哈河、沙柳河(伊克乌兰河)、甘子河、倒淌河等河流和青海湖构成。研究区植被类型主要包括灌丛、草原、沙生植被、盐生植被、沼泽草甸植被、高山流石植被等[23]。

图1 青海湖流域位置
Figure 1 Location of the Qinghai Lake Basin
1.2 数据与方法
1.2.1 数据来源
本研究运用了MODIS产品中的2000—2018 年的GPP和NPP数据, MOD17A2H遥感产品来自于NASA网站(http://modis.gsfc.nasa.gov/), 该产品可提供连续的全球监测初级生产力数据。MOD17A2H数据的时间分辨率和空间分辨率分别是8 d和500 m, 且该数据已经过去云、辐射校正、大气校正和几何校正等步骤。利用ArcGIS 10.2软件和ENVI 5.3软件对MODIS数据进行处理, 其中包括, 投影转换、拼接、裁剪、按照元数据文件指示去除异常值、转换比例因子等操作, 以恢复原始GPP和NPP的值。本研究所使用的增强型植被指数(enhanced vegetation index, EVI)来自MODIS数据产品的MOD13Q1数据, 其时间分辨率和空间分辨率分别是16 d和250 m, 数据格式为HDF, 该产品已经过大气校正、几何校正和去云等处理, 获取数据后进行投影转换和最大值合成(MVC), 最后利用研究区矢量边界裁剪后即可应用。
1.2.2 CUE计算方法
CUE定义为生态系统植被净初级生产力与总初级生产力的比值, 则其表达式为[24-25]:
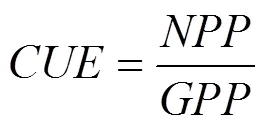

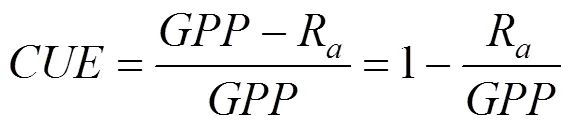
式中:代表生态系统植被碳利用效率;为植被净初级生产力(g·m-2);为植被总初级生产力(g·m-2);R为植被自养呼吸(g·m-2)。
1.2.3 CUE年际变化率计算
本文基于一元线性回归分析方法来模拟2000— 2018年青海湖流域生态系统植被CUE的时空变化趋势。计算公式如下[26]:

式中:为植被的趋势斜率;为时间序列长度, 即2000—2018 年共计19 年;为1—19 年的序列号;CUE为第年的值。> 0, 表示在19年间的趋势为增长, 反之则为减少。
2 结果
2.1 青海湖流域植被CUE年内变化特征
近19年青海湖流域植被CUE年内各月均值变化显示(表1), 植被CUE年内变化明显。近19年青海湖流域植被CUE各月平均值从3月开始增加, 6月和7月达到最大值0.58, 而后开始下降; 11月到来年2月NPP接近于0, 故此判断其CUE接近于0。植被在不同生长阶段的CUE空间分布特征显示(图2), 5月青海湖流域植被CUE在0.39—0.8之间变化(除水域之外), 且植被CUE值在东部偏高, 西部较低, 最高值出现在青海湖东北岸(图2a); 8月青海湖流域植被CUE在0.53—0.78之间变化(除水域之外), 植被CUE值在西部边界区偏高, 青海湖北岸中部和西岸中部较低, 最高值出现在青海湖东北岸和西北边界地区(图2b); 9月青海湖流域植被CUE在0.60—0.79之间变化(除水域之外), 植被CUE在青海湖西岸的中部地区较8月有明显降低的趋势(图2c); 10月青海湖流域植被CUE在0.34—0.80之间变化(除水域之外), 青海湖流域植被CUE由东南向西北减小(图2d)。5月和10月青海湖流域植被CUE总体由东南向西北逐渐递减, 最高值区域分布在青海湖北岸附近。8月和9月植被CUE依然以青海湖为中心, 呈“高—低—高”的环带状分布。总体上, 青海湖环湖地区为植被CUE最大值分布区域, 青海湖流域中部地区在生长季初期和末期CUE值偏高, 生长旺盛期较低, 青海湖边界地区与中部地区相反(图2)。
2.2 青海湖流域植被CUE年际变化特征
2000—2018年青海湖流域植被CUE的年际变化趋势显示(图3), 近19年青海湖流域植被CUE整体呈略微下降趋势, 线性递减率为0.01·10–1a。青海湖流域年均植被CUE在0.55—0.63之间变化, 平均值为0.58, 波动幅度较小, 最小值出现在2018年, 最大值出现在2009年。青海湖流域典型年份植被CUE结果显示(图4), 2000年青海湖流域植被CUE在0.54—0.8之间变化(除水域之外)(图4a), 最小值区域出现在青海湖北岸的中部地区; 2009年青海湖流域植被CUE在0.54—0.8之间变化(除水域之外), 流域大部分地区植被CUE值高于2000年和2018年(图4b); 2018年青海湖流域植被CUE在0.42—0.80之间变化(除水域之外), 流域大部分地区植被CUE值低于2000年和2009年(图4c)。这3年的最小值区域都出现在青海湖北岸的中部地区, 最大值出现在流域西北边界处和青海湖东北岸, 2009年整体值偏大, 2018年最大值出现在青海湖东北岸小部分区域。在青海湖流域中部地区(青海湖以西), 2009年值最大, 2000年次之, 2018年最小, 由此可见, 2009年和2018年青海湖流域植被CUE并非异常值。

表1 青海湖流域不同月份植被CUE均值变化

图2 2000—2018年青海湖流域典型月份的CUE空间变化
Figure 2 Spatial variation of CUE valuein typical month in Qinghai Lake Basin during 2000-2018
2.3 青海湖流域植被CUE空间变化趋势
近19青海湖流域植被年均CUE的空间分布显示(图5), 青海湖流域内植被CUE在0.54—0.76之间变化(除水域之外), 且流域内平均值为0.58, 青海湖流域植被CUE分布呈现以青海湖为中心的“高—低—高”环带状分布特征, 从东南向西北环带范围扩大。在青海湖环湖地区出现了第一条环带状“高”值区, 且青海湖北岸的高值区大于南岸, 这一高值区主要以农田和干旱草原为主, 其值大约在0.70左右。在青海湖流域中部以灌丛和高寒草甸生态系统为主的地区形成了“低”值区, 主要分布在青海湖四周的沙柳河、哈尔盖河、甘子河、布哈河、黑马河中游一带, 其值在0.59左右。最后“高”值区, 出现在青海湖流域的边界地区, 其中最大区域出现在青海湖流域的西北部, 此高值区主要以高山流石植被为主, 其值大约在0.70左右。由于环境因子随时间的变化存在差异, 植被CUE的年际变化在空间上也存在差异(图6), 变化趋势(SLOPE值)范围在-0.02—0.03之间, 将变化趋势分为减少(< 0)、不变(≈ 0)和增加(> 0)3部分。青海湖流域有81.02 %的区域为植被CUE减少区域, 有15.17 %的区域为植被CUE不变区域, 其中青海湖湖面占14.76 %, 即青海湖流域植被CUE只有0.41 %的区域保持不变, 只有3.81 %的区域为植被CUE增加区域, 而其中不变的区域主要是青海湖东北岸, 增加的区域零星散布在青海湖东部、北部和西部。

图3 2000—2018年青海湖流域CUE动态变化
Figure 3 Dynamics of annual CUE value in Qinghai Lake Basin during 2000-2018

图4 2000—2018年青海湖流域典型年份CUE空间变化
Figure 4 Spatial variation of CUE valuein typical year in Qinghai Lake Basin during 2000-2018

图5 2000-2018年青海湖流域CUE年均值空间分布
Figure 5 Spatial distribution of CUE value in the Qinghai Lake Basin

图6 2000-2018年青海湖流域CUE空间变化趋势
Figure 6 Spatial variation trend of CUE value in Qinghai Lake Basin during 2000-2018
3 讨论
青海湖流域植被CUE的时空分布是受植被类型、气温、降水、海拔和土地营养和管理措施等诸多因素综合影响的结果。青海湖流域内植被CUE从3月开始增大, 6、7月达到最大值, 随后开始减小(表1)。5月和10月植被CUE呈现由东南向西北递减的趋势(图2a、d), 这主要是由于海拔由低到高呈环带状分布, 自东南向西北升高, 西北部生长季较东部时间短, 使得5月和10月青海湖流域植被CUE呈现由东南向西北递减的规律。6月到9月, 青海湖流域植被处于生长季内, 故此植被CUE变化较为稳定(图7b、c)。
青海湖流域GPP、NPP与气温和降水存在正相关关系(图7), GPP、NPP与温度为极显著相关(图7a、b),与降水为显著相关(图7c、d)。Lieth等[27]研究发现, 干冷生态系统植被NPP随年降水量和年均气温的升高呈线性增加。闫敏等[28]对黑河上游地区总初级生产力研究中发现, 高寒草甸地区植被GPP与温度和降水呈正相关关系。温度、降水与GPP的拟合斜率远大于与NPP的拟合斜率(图7), 当温度和降水增加量相同时, GPP的增加量大于NPP的增加量, 从而间接导致植被CUE降低。温度降低时, 植被维持自身组织所需的能量减少[29], Ra降低, 植被CUE增大。降水较多时, 云量较多, 土壤出现缺氧状态, 有机质的分解速率下降, 从而导致了CUE减小[30]。温度和降水影响生态系统植被光合作用和自养呼吸, 间接影响植被CUE的变化。近19年青海湖流域植被CUE呈现略微下降趋势(图3, 图6), 这一研究与前人研究结果相吻合[1,12,31], 近19年平均气温升高使Ra增加, 导致植被CUE减小。2009年青海湖流域植被CUE出现最大值为0.63, 其原因主要是2009年的生长旺盛时期(7月和8月)平均气温(12.11 ℃)比近19年平均气温(13.01 ℃)低0.9 ℃, 低温环境致使生态系统植被自养呼吸作用减弱, 使植被CUE增大(图4b)。2018年青海湖流域植被CUE出现最小值为0.55, 其原因主要是2018年的生长旺盛时期(7月和8月)降水达到近19年的最大值127.88 mm, 降水较多时, 土壤出现缺氧状态, 有机质的分解速率下降, 同时植被的光合作用效率下降, 使得植被CUE减小(图4c)。

图7 青海湖流域年均NPP、GPP与年均气温、年降水量以及VPD与CUE的关系
Figure 7 The relationship between annual NPP, GPP and annual temperature, precipitation, VPD and CUE in Qinghai Lake Basin
饱和水汽压差(vapor pressure deficit, VPD)是指在一定温度下, 饱和水气压和空气之中实际水汽压的差值, 是空气干燥程度的一个指标, 其值大小可以反映空气水分亏缺状况, VPD还是叶片和大气界面物质交换的驱动力, 对植物体的生理代谢有重要影响[32]。植物气孔是植物体与外界大气进行气体交换的通道, 在光合作用过程中, CO2通过气孔进入叶片, 参与植物的光合作用, 而VPD则是通过调节植被的气孔来影响植被的光合和呼吸速率[33]。青海湖流域的植被CUE与VPD存在较显著的负相关关系, 即植被CUE随VPD的增加而减小(图7f)。温度和降水的正相关性混淆了植被CUE对于气温降水的相关性[29], 同时, 植被CUE受氮沉降、环境条件、光照强度等的综合影响, 且各影响因子之间的相互关系, 更加混淆了植被CUE与影响因子的相关性。青海湖流域地处干旱区, 温度较低, 多年平均VPD为0.32 KPa, 温度增加, VPD开始增大(VPD < 0.5 KPa), 气孔开放, 蒸腾作用加强而光合作用却不受影响[34], 呼吸作用增加, 导致了高Ra/GPP和低植被CUE(1-Ra/GPP)。降水增加时, 植被叶片气孔关闭, 阻碍了植被光合作用, 同时在一定程度上促进了呼吸作用, 同样造就了高Ra/GPP和低植被CUE[35]。
青海湖流域植被CUE在0.54—0.76之间变化(除水域之外), 平均值为0.58。袁旻舒等[36]研究中国陆地生态系统总体植被CUE变化范围在0.33—0.62之间, 且平均值保持在0.5左右, 变化幅度不大; Street[37]等对欧洲亚北极区植被CUE的研究结果表明, 矮灌丛生态系统植被CUE值在0.58—0.74之间。上述植被CUE研究与本文所得结果基本吻合。植被CUE不但受气温、降水的影响, 同时也受植被盖度和植被自养呼吸的影响。增强型植被指数(enhanced vegetation index, EVI)是表征生态系统植被覆盖状况变化的一个重要指标, EVI比归一化植被指数(NDVI)对植被变化的敏感性更强[38], EVI值越大表示植被覆盖度越大。Chen等[39]研究认为EVI与生态系统GPP有正相关关系。安相等[15]研究认为EVI对生态系统光合作用和呼吸作用都有重要影响, 且植被生长状况越好, 生态系统GPP越高, 但对NPP的影响不大。2000—2018年EVI与GPP和NPP的对比显示(图8), 青海湖流域EVI与GPP、NPP的变化趋势一致, GPP和NPP随EVI的增大而增加。GPP、NPP与EVI的相关分析表明, 青海湖流域GPP、NPP与EVI呈极显著正相关关系(<0.01), 确定系数为R2=0.38和R2=0.42, 其拟合方程显示, EVI的增大量相同时(y=1.268x-97.597, y=0.862x-62.785), GPP的增加量大于NPP的增加量, 从而间接导致植被CUE的值降低。
2000—2018年青海湖流域EVI和Ra平均值显示(图9), 青海湖流域EVI和Ra以青海湖为中心由东南向西北形成了“低—高—低”的环带状分布特征, 与植被CUE分布特征恰好相反(图5)。Ra是植物为维持自身生存所消耗的同化物, 在一个生态系统中植物的自养呼吸大小决定了生态系统植被固碳能力的强弱。He等[29]在用不同方法估算全球CUE时发现, 植被CUE的主要差异取决于Ra方面, 而不在GPP方面。青海湖流域以青海湖为中心, 海拔由低到高呈环带状分布, 自东南向西北升高, 气温和降水以青海湖为中心呈现由暖到寒、由湿到干的环带状分布[40]。青海湖环湖地区主要以农田和草地为主, EVI和Ra值较小, 农田灌溉和施肥能引起植被根部呼吸的减少[41], 植被覆盖度小则地上呼吸较少, 植被CUE较大(图5), 研究表明农田生态系统、草地生态系统、森林生态系统植被CUE的平均值依次减小[42-43]。青海湖流域中部地区以灌丛和高寒草甸为主, EVI和Ra值较大, 温度和降水充足, 植被生长茂盛, 自养呼吸消耗量较大, 导致植被CUE值较小(图5)。青海湖流域边缘地区主要以流石坡植被为主, 植被稀疏, 海拔较高, 随海拔升高温度和降水减少, 植被用于生存所耗费的能量较少[44-45], 植被固碳转移效率更高[46], 植被CUE较大(图5)。在适应干旱或寒冷环境时, 生态系统通常出现以短茎或稀疏覆盖为特征的生命形式[13], 由此造就了稀疏植被生态系统(EVI和Ra较小)的植被CUE值较大。Zhang等[13]在基于MODIS数据的基础上分析CUE变化时表明, 北温带地区茂盛植被地区的CUE比稀疏植被地区低, 与本研究结果相似。

图8 青海湖流域年均NPP、GPP与EVI的关系
Figure 8 The relationship between annual NPP, GPP and EVI in Qinghai Lake Basin

图9 2000—2018年青海湖流域EVI和Ra空间变化
Figure 9 Spatial variation of EVI andavalue in Qinghai Lake Basin during 2000-2018
4 结论
本文基于MODIS数据估算并分析青海湖流域2000—2018年间的植被CUE时空变化规律, 认为青海湖流域植被CUE并非恒定值。得出如下初步结论: (1)青海湖流域内植被CUE从3月开始增大, 6、7月达到最大值, 随后开始减小。青海湖流域海拔由西北向东南递减, 西北部生长季较东部时间短, 使得5月和10月青海湖流域植被CUE呈现由东南向西北递减的规律。(2)温度和降水增加量相同时, GPP的增加量大于NPP的增加量, 从而间接导致植被CUE的值降低。(3)青海湖流域EVI和Ra以青海湖为中心由东南向西北形成了“低—高—低”的环带状分布特征, 与CUE分布特征恰好相反。生态系统植被生长茂盛时, EVI和Ra增大, EVI对GPP影响大于对NPP的影响, 同时自养呼吸消耗量增大, 导致植被CUE值较小。
[1] CHAMBERS J Q, TRIBUZY E S, TOLEDO L C, et al. Respiration from a tropical forest ecosystem: partitioning of sources and low carbon use efficiency[J]. Ecological Applications, 2004(14): 72–88.
[2] RYAN M G, LAVIGNE M B, Gower S T. Annual carbon cost of autotrophic respiration in boreal forest ecosystems in relation to species and climate[J]. Journal of Geophysical Research Atmospheres, 1997, 102(D24): 28871–28883.
[3] DILLAWAY D N, KRUGER E L. Trends in seedling growth and carbon-use efficiency vary among broadleaf tree species along a latitudinal transect in eastern North America[J]. Global Change Biology, 2014, 20(3): 908–922.
[4] ALLISONll S D, WALLENSTEIN M D, BRADFORD M A. Soil-carbon response to warming dependent on microbial physiology[J]. Nature Geoscience, 2010, (3): 336–340.
[5] MANZONI S, TAYLOR P, RICHTER A, et al. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils[J]. New Phytologist, 2012, 196: 79–91.
[6] 朱万泽. 森林碳利用效率研究进展[J]. 植物生态学报, 2013, 37(11): 1043–1058.
[7] LINDORTH A, GRELLE A, MOREN A S. Long-term measurements of boreal forest carbon balance reveal large temperature sensitivity[J]. Global Change Biology, 1998, 4(17): 443–450.
[8] DELUCIA E H, DRAKE J E, THOMAS R B, et al. Forest carbon use efficiency: Is respiration a constant fraction of gross primary production[J]. Global Change Biology, 2007, 13(6): 1157–1167.
[9] DEWAR R C, MEDLYN B E, MCMURTRIE R E. A mechanistic analysis of light and carbon use efficiencies[J]. Plant Cell and Environment, 1998, 21(6): 573–588.
[10] 陈光水, 杨玉盛, 高人, 等. 杉木林年龄序列地下碳分配变化[J]. 植物生态学报, 2008, 32( 6): 1285–1293.
[11] MALHI Y, ARAGAO L, METCALFE D B, et al. Comprehensive assessment of carbon productivity, allocation and storage in three Amazonian forests[J]. Global Change Biology, 2009, 15(5): 1255–1274.
[12] KWONA Y, LARSEN C P S. Effects of forest type and environmental factors on forest carbon use efficiency assessed using MODIS and FIA data across the eastern USA[J]. International Journal of Remote Sensing, 2013, 34(23): 8425–8448.
[13] ZHANG Yangjian, XU Ming, CHEN Hua, et al. Global pattern of NPP to GPP ratio derived from MODIS data: effects of ecosystem type, geographical location and climate[J]. Glob Ecology and Biogeography. 2009, 18(3): 280–290.
[14] LEVY P E, CANNELL M G R, FRIEND A D. Modelling the impact of future changes in climate, CO2concentration and land use on natural ecosystems and the terrestrial carbon sink[J]. Global Environmental Change, 2004, 14(1): 21–30.
[15] 安相, 陈云明, 唐亚坤. 东亚森林、草地碳利用效率及碳通量空间变化的影响因素分析[J]. 水土保持研究, 2017, 24(5): 80–92.
[16] PIAO Shilong, LUYSSAERT S, CIAIS P, et al. Forest annual carbon cost: a global-scale analysis of autotrophic respiration[J]. Ecology, 2010, 91, 652–661.
[17] 吕鑫, 王卷乐, 康海军, 等. 基于MODIS NPP的2006-2015年三江源区产草量时空变化研究[J]. 自然资源学报, 2017, 32(11): 1857–1868.
[18] 翟俊, 侯鹏, 赵志平, 等. 青海湖流域景观格局空间粒度效应分析[J]. 国土资源遥感, 2018, 3(3): 159–166.
[19] 曹生奎, 曹广超, 陈克龙, 等. 青海湖高寒湿地生态系统服务价值动态[J]. 中国沙漠, 2014, 34(5): 1402–1409.
[20] 潘虹, 顾海敏, 史建桥, 等. 基于RS和GIS的青海湖流域植被覆盖度变化与驱动因子研究[J]. 资源开发与市场, 2016, 32(7): 827–831.
[21] 苏芬, 刘宝康, 张翠花, 等. 青海湖流域牧草物候期对气候变化的响应[J]. 青海草业, 2018, 27(4): 12–18.
[22]肖雄, 李小雁, 吴华武, 等. 青海湖流域高寒草甸壤中流水分来源研究[J]. 水土保持学报, 2016, 30(2): 230–236.
[23] 乔凯, 郭伟. 青海湖流域植被的净初级生产力估算[J]. 水土保持通报, 2016, 36(6): 204–209.
[24] 方精云, 柯金虎, 唐志尧, 等. 生物生产力的“4P”概念、估算及其相互关系[J]. 植物生态学报, 2001, 25(4): 414– 419.
[25] WARING R H, LANDSBERG J J, WILLIAMS M. Net primary production of forests: a constant fraction of gross primary production[J]. Tree physiology, 1998, 18(2): 129–134.
[26] 穆少杰, 李建龙, 周伟, 等. 2001-2010年内蒙古植被净初级生产力的时空格局与气候变化的关系[J]. 生态学报, 2013, 33(12): 3752–3764.
[27] LIETH H, WHITTAKER R H. Modeling the primary productivity of the world. Primary productivity of the biosphere[M]. New York: Springer Verlag Press, 1975(12): 237–263.
[28] 闫敏, 李增元, 田昕, 等. 黑河上游植被总初级生产力遥感估算及其对气候变化的响应[J]. 植物生态学报, 2016, 40(1): 1–12.
[29] HE Yue, PIAO Shilong , LI Xiangyi, et al. Global patterns of vegetation carbon use efficiency and their climate drivers deduced from MODIS satellite data and process-based models[J]. Agricultural and Forest Meteorology, 2018, 256-257: 150–158.
[30] MATSON S P A . Net Primary Productivity and Nutrient Cycling across a Mesic to Wet Precipitation Gradient in Hawaiian Montane Forest[J]. Oecologia, 2001, 128(3): 431–442.
[31] ZHANG Yangjian, YU Guirui, YANG Jian, et al. Climate-driven global changes in carbon use efficiency[J]. Global Ecology and Biogeography, 2014, 23(2): 144–155.
[32] 杜清洁, 宋小明, 柏萍, 等. 不同水汽压差对番茄气体交换参数和生长的影响及综合评价[J]. 西北农业学报, 2020, 29(01): 66-74.
[33] 牛亚毅, 李玉强, 王旭洋, 等. 干旱年份沙质草地生态系统净 CO2通量年变化特征[J]. 草业学报, 2018, 27(1): 215–221.
[34] 阚飞, 岳宁, 丁林凯, 等. 不同气象条件下陇中玉米农田生态系统水分利用效率研究[J]. 灌溉排水学报, 2018, 37(10): 116–122.
[35] 陈小平. 科尔沁沙丘-草甸湿地水热碳通量变化及响应机制研究[D]. 呼和浩特: 内蒙古农业大学, 2018.
[36] 袁旻舒, 李明旭, 程红岩, 等. 基于CMIP5 模型结果的中国陆地生态系统未来碳利用效率变化趋势分析[J]. 中国科学院学学报, 2017,34(4): 452–461.
[37] STREET L E, SUBKE J A, SOMMERKORN M, et al. The role of mosses in carbon uptake and partitioning in arctic vegetation[J]. New Phytologist, 2013, 199: 163–175.
[38] 杨怡, 吴世新, 庄庆威, 等. 2000—2018 年古尔班通古特沙漠 EVI 时空变化特征[J]. 干旱区研究.2019, 36(06): 1512–1520.
[39] CHEN Zhi, YU Guirui, GE Jianping, et al. Roles of climate, vegetation and soil in regulating the spatial variations in ecosystem carbon dioxide fluxes in the Northern Hemisphere[J]. PloS One, 2015, 10(4): 1–l4.
[40] 张涛, 曹广超, 曹生奎, 等. 2000—2012年青海湖流域NPP时空分布特征[J]. 中国沙漠, 2015, 35(4): 1072– 1080.
[41] RYAN M G, HUBBARD R M, PONGRACIC S, et al. Foliage, fine root, woody tissue and stand respiration in Pinus radiata in relation to nitrogen status[J]. Tree Physiology, 1996, 16, 333–343.
[42] CHEN Zhi, YU Guirui, WANG Qiufeng. Ecosystem carbon use efficiency in China: Variation and influence factors[J]. Ecological Indicators, 2018, (90): 316–323.
[43] MUHAMMAD K, NADIR A E, LARS R, et al. Spatio-temporal variations in climate, primary productivity and efficiency of water and carbon use of the land cover types in Sudan and Ethiopia[J]. Science of the Total Environment, 2018, (624): 790–806.
[44] RYAN M G, LINDER S, VOSE J M, et al. Dark respiration of pines[J]. Ecological bulletins, 1994, 43: 50–63.
[45] CANNEL M G R. Physiological basis of wood production: a review[J]. Scandinavian journal of forest research, 1989, 4(1/4): 459–490.
[46] ENQUIST B J, KERKHOFF A J, STARK S C, et al. A general integrative model for scaling plant growth, carbon flux, and functional trait spectra[J]. Nature, 2007, 449, 218–222.
Temporal and spatial dynamics of vegetation carbon use efficiency in Qinghai Lake Basin
LAN Yao1, 2, 3, CAO Shengkui1 2, 3,*, CAO Guangchao2, 3, LI Xiaodong4, YANG Yufan1, 2, LEI Yizhen1, 2, 3
1. College of Geographical Sciences, Qinghai Normal University, Xining 810008, China 2. Qinghai Provincial Key Laboratory of Physical and Environmental Processes, Qinghai Normal University, Xining 810008, China 3. Academy of Plateau Science and Sustainability, People’s Government of Qinghai Province & Beijing Normal University, Xining 810008, China 4. Qinghai Institute of Meteorological Science, Xining 810001, China
The carbon use efficiency (CUE) is a scale to measure the carbon sequestration capacity and efficiency of ecosystem vegetation; at the same time, it is of great significance to evaluating the carbon storage and reveal the mechanism of carbon balance.This paper takes Qinghai Lake Basin as the research object and uses MOD17A2H to estimate the vegetation CUE from 2000 to 2018 and to reveal it characteristics of dynamic change and impact factors.The results showed that: (1) The vegetation CUE of annual variation was increasing from March and reaching the maximum in June and July then decreasing; and it was 0.58 of average annual, and it varied between 0.55 and 0.63 and slightly decreased, and the linear decline rate was 0.01/10a in Qinghai Lake Basin.(2) The CUE of vegetation spatial distribution characteristics was the “high-low-high” ring-like shape centered, and it varied between 0.54 and 0.76 (except the waters) and the average was 0.58 in the Qinghai Lake Basin. (3) The CUE of vegetation was mainly affected by temperature and precipitation, and the effect of temperature was more obvious than precipitation. With the increase of temperature and precipitation, the CUE of vegetation decreased in Qinghai Lake Basin.The results of research can provide data support for clarifying the source/sink problem in the carbon cycle of the region and provide reference for the study of vegetation CUE in areas of alpine inland. It is of great significance to understanding global change and predicting the response of ecosystem to global change.
MODIS; Qinghai Lake Basin; vegetation; carbon use efficiency; spatial and temporal variations
10.14108/j.cnki.1008-8873.2020.04.020
兰垚, 曹生奎, 曹广超, 等. 青海湖流域植被碳利用效率时空动态研究[J]. 生态科学, 2020, 39(4): 156–166.
LAN Yao, CAO Shengkui, CAO Guangchao, et al. Temporal and spatial dynamics of vegetation carbon use efficiency in Qinghai Lake Basin[J]. Ecological Science, 2020, 39(4): 156–166.
Q148
A
1008-8873(2020)04-156-11
2019-11-25;
2019-12-31
青海省“135高层次人才培训工程”; 青海省“高端创新人才千人计划”项目(青人才字[2016] 11号); 青海省重点实验室发展专项
兰垚(1993—), 男, 汉族, 陕西岐山人, 硕士研究生, 研究方向为生态水文及生态系统碳循环方面, E-mail: 1246522453 @qq.com
曹生奎, E-mail: caoshengkui@163.com
