不同植物根际土壤碳氮水解酶活性热点区的空间分布特征
2020-08-19张心昱
刘 霜,张心昱,*
1 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101 2 中国科学院大学资源与环境学院, 北京 100190
植物根际是植物与土壤微生物相互作用最活跃的界面,是研究根系与土壤微生物养分获取机制的热点场所[1]。在根际,受根系分泌物影响,土壤微生物大量富集,植物和微生物分泌的土壤酶能高效催化土壤有机质分解,如,β-葡萄糖苷酶(βG)降解纤维素,将纤维素二糖水解为葡萄糖,为根际土壤微生物提供能量;N-乙酰氨基葡萄糖苷酶(NAG)降解几丁质和肽聚糖,将壳二糖水解为氨基葡萄糖(葡萄糖胺),为植物和微生物提供氮养分。因为βG在纤维素降解,NAG在几丁质降解中起同样关键作用,并且是土壤碳、氮循环过程的代表性指示物研究最多[2- 3],因此本文以这两种水解酶为代表进行根际土壤碳氮水解酶活性热点区空间分布特征的研究[1]。植物根际酶活性热点区的分布范围可以代表根系和根际微生物所需能量和养分的空间范围,而根际土壤的碳氮酶活性的特征则可用来揭示根系和根际微生物能量和氮养分获取策略。
土壤酶活性已成为量化陆地生态系统土壤质量和功能的重要指标,而精确、合理、高效的测定方法是研究的基础。常用的土壤酶活性测定技术测得的酶活性仅为土壤的最大潜在酶活性,不能准确反映原位土壤环境中的真实活性。土壤原位酶谱法的出现为研究植物根系和根际微生物的养分获取策略提供了新方法[4]。利用土壤原位酶谱法,Razavi等发现酸性土壤种植的玉米和扁豆根际酶活性热点区分布规律显著不同,玉米由于根尖分泌物丰富,酶活性热点区主要集中在根尖(大约2 mm),而扁豆无论是根尖还是根伸长区热点区都是均匀分布(1—1.5 mm),这是由于扁豆根系各个部位都能与根瘤菌共生固氮,所以扁豆根系土壤微生物为了固氮而维持一种潜在的均质的根际环境[5]。Liu等通过土壤原位酶谱法研究了将有机肥施于青藏土壤表面或与土壤混匀两种处理方式下青藏大麦根系的养分获取机制,结果发现根际酶活性在有机肥施于土壤表面时比将有机肥与土壤混匀处理更高。这是因为有机肥与土壤混匀后植物和微生物要适应新的养分来源,降低根对土壤养分的吸收以及土壤微生物的活性[6]。Ma等通过土壤原位酶谱法发现水稻根际土壤磷酸酶热点区范围(3 mm)远远大于氮水解酶热点区范围(1.5 mm)[7],Razavi等也发现玉米和扁豆磷水解酶的热点区范围(2.5—3.5 mm)大于碳氮水解酶热点区范围(分别为1.5—2 mm和1—1.5 mm),这些结果也表明土壤中的有效磷相对匮乏,而磷是植物生长的关键因子以及细胞合成腺苷三磷酸(ATP)、核酸磷脂等的重要组分,所以根系和根际微生物对不同养分元素的需求不同[5]。
中国西南喀斯特土壤分布区是世界三大喀斯特土壤分布区之一,过去由于人地矛盾突出,坡地开垦为耕地,结果导致土壤侵蚀、养分淋失、土壤质量下降,影响植被恢复。20世纪初,中国政府在该区域坡度>15°的耕地实施退耕还林、还草政策,过去耕地以种植玉米为主,退耕后,大量杂草(如莎草)生长,部分农田退耕后种植牧草,如苜蓿。中国西南喀斯特土壤类型为石灰土,pH值7—8[8]。受根系分泌物影响,根际土壤酸化,土壤pH低于非根际土壤[9],根际土壤微生物活性更高,根系和根际微生物的养分获取更高[10]。但是目前关于喀斯特植物根际不同部位土壤碳、氮酶活性热点区的空间分布特征的研究较少,关于玉米、苜蓿和莎草根尖区和伸长区获取碳、氮养分的机制尚不清楚。
本文采用典型喀斯特坡耕地0—20 cm土壤,选择玉米、苜蓿和莎草进行室内根盒培养试验,利用土壤原位酶谱法分析技术,分析不同植物根际土壤碳氮水解酶活性热点区的空间分布规律。1)因为玉米根系比苜蓿和莎草更发达,根尖分泌物数量多,所以我们假设玉米根尖的根际酶活性热点区范围比莎草和苜蓿大;2)由于植物根尖分泌物比根伸长区更丰富,我们假设根尖的水解酶热点区范围要大于根伸长区;3)苜蓿为固氮植物,与非豆科的莎草和玉米相比,苜蓿根际土壤分泌氮水解酶获取氮的需求较低,所以我们假设苜蓿氮水解酶的根际酶活性热点区小于碳水解酶。本研究将揭示喀斯特植被根际土壤碳氮水解酶活性空间分布规律,可为喀斯特土壤退耕后植被的选择、地下养分管理提供科学依据。
1 材料和方法
1.1 研究区概况
土壤采集于贵州省普定县陈旗小流域(26°15′N-26°16′N,105°46′E—105°47′E),平均海拔为1400 m。研究区属于典型的亚热带季风气候,年均温15.1℃,年降水量1390 mm。最高温7月,最低温1月,雨季在5月—10月期间。根据中国土壤分类,本研究区土壤类型为石灰土,是美国发生分类的松软始成土[11]。研究区主要农物是玉米、红薯、大豆等[12]。
在2018年7月,选取2 m×2 m坡耕地样方4个,每个样方间距大于10 m,每个样方采用5点混合的方法,采集0—20 cm土壤,将所有采集土壤样品混匀。土壤含水量13%,pH值 8.27,土壤总碳含量36.7 g/kg,总氮含量2.78 g/kg,总磷含量487 mg/kg,可溶性有机碳含量29 mg/kg,铵态氮含量11 mg/kg,硝态氮含量12 mg/kg,有效磷含量4 mg/kg。
1.2 根盒培养试验
采用室内根盒培养的方法,选择3种植物,即当地粮食作物玉米(Zeamays)、牧草苜蓿(Cyperusrotundus)及退耕后常见植物莎草(Medicagosativa),在内尺寸为20 cm×20 cm×3 cm的根盒中培养,每种植物5次重复,总计15个根盒。将根盒能打开的一面向上水平放置,把过2 mm筛的土壤缓慢且均匀地填满根盒,关闭根盒并垂直轻轻摇动土壤,避免土壤分层。用滤纸在室温育种72 h,挑选长势良好的种子,移种在根盒能打开面的土体表层距顶端1 cm处。将根盒向能打开面倾斜45°放置,保持室内温度(20±1)℃和光照16 h,培养30 d。在培养期间,每天对根盒称重,用蒸馏水补充水分,保持土壤含水量控制在田间持水量的60%左右。
1.3 根际土壤原位酶谱分析
在根盒培养30 d后,进行根际土壤原位酶谱分析实验。主要参考Spohn和Kuzykov的方法[13]。用浸泡了含有甲基伞形酮(MUB)的底物溶液浸泡滤膜,当特定底物被专一水解酶分解后会释放荧光物质,可以在紫外灯(Philips, TL-D 18W BLB)下显色。βG活性采用4-甲基伞形酮酰-β-D-吡喃葡萄糖苷为底物,NAG活性用4-甲基伞形酮酰-乙酰基-β-D-氨基葡萄糖苷为底物。将每种底物分别溶解于10 mmol/L MES(化学分子式:C6H13NO4SNa0.5)缓冲溶液中,缓冲液的pH为8.27,与土壤pH一致。用尺寸为10 cm×10 cm,孔径为0.45 μm尼龙滤膜(桃园 N66亲水尼龙微孔滤膜)浸满底物,打开根盒,将浸满βG、NAG底物的滤膜依次覆盖在根系表面培养1 h,取出后用镊子轻轻地剔除滤膜表面土壤颗粒,在紫外灯(455 nm)下激发1 min后照相,照相时固定相机(Leica D-LUX 6)、滤膜与紫外灯的距离。通过预实验和文献[14]确定1 h的培养时间。
为了定量计算酶谱图像的酶活性值大小,拟合了不同浓度MUB酶谱图像的灰度值与酶活性之间的函数关系。参考上述根际土壤原位酶谱分析方法,用2 cm×2 cm的滤膜浸满100 μL 的0、0.5、1、2、3、4、6、8、10 mmol/L 9个浓度梯度的MUB,根据滤膜吸收溶液的量及滤膜尺寸计算各个浓度滤膜上MUB的量(10-12mol/cm2)。将膜放在紫外灯下激发1 min后照相,在软件中分析酶谱图像。
1.4 酶谱图像处理过程与数据分析
酶谱图像中酶活性定量分析的方法主要根据Razavi等[14]的方法进行图像处理。图像分析的步骤如下,首先,在Image J中将图像转化为16 bit灰度图像;然后,确定土壤背景值校正图像,根据标准曲线将灰度值转换为酶活性值;最后在Matlab中将酶谱图像转换成彩色图像(图1)。土壤背景值是基于酶谱图像中非根际土壤的灰度值的平均值计算,本研究中为50×10-12mol cm-2h-1。

图1 根盒培养、与底物结合培养、紫外光激发成像与酶活性定量过程示意图Fig.1 Schematic diagram of rhizo-box cultivation, substrate-combing incubation, Ultraviolet light excitation imaging and enzyme activity quantification
根据酶谱图和标准曲线,计算所有像素的酶活性值,并计算所有酶谱图像根系周围5 mm(沿根尖向上及根伸长区向外)的酶活性数值,经统计软件分析符合正态分布,根据酶活性正态分布曲线,将大于30%的酶活性定义为热点区[15],在本研究中酶活性大于200×10-12mol cm-2h-1为热点区。
根据不同酶活性热点值下限(200×10-12mol cm-2h-1),计算3种植物沿根尖向上(记为X)和沿根伸长区向外(记为Y)两个方向上的碳氮水解酶活性的热点区分布范围,即根际酶活性热点区范围。由此可得到3种植物根系两个部位的碳氮水解酶的根际酶活性热点区范围,取碳、氮水解酶根际酶活性热点区最大值记为Xn和Yn(n=玉米,苜蓿,莎草)。在酶谱图像中以每种酶的Xn和Yn为半径设置缓冲区,其中由最大(Xn,Yn)值组成的缓冲区面积为该种植物的最大根际酶活性热点区面积,表示植物根系能够利用土壤碳和氮的潜在根际酶活性热点区面积。并计算碳或氮水解酶活性热点区面积占每种植物最大根际酶活性热点区面积的比值,表示该植物与根际微生物对土壤碳氮偏好度。公式如下:

1.5 统计分析
采用Levene检验和Shapiro Wilk W检验3种植物从根沿根尖向上及根伸长区向外两个方向分布的酶活性,3种植物根尖、根伸长区根际酶活性热点区范围、根系对土壤碳氮养分偏好度符合正态分布及方差齐性。采用单因素方差分析和LSD检验,分析3种植物根尖、根伸长区根际酶活性热点区范围,同一植物不同酶根尖、根伸长区根际酶活性热点区范围,根系对土壤碳氮养分偏好度的显著性分析,P<0.05为差异显著。所有统计分析在SPSS 19.0软件中进行,用SigmaPlot 10.0软件作图。数据为平均值±标准误(n=3)。
2 结果
2.1 不同植物根际土壤酶活性的空间分布
经过30 d根盒培养后,根的半径大小为玉米根(0.45 mm)>苜蓿(0.3 mm)>莎草(0.2 mm)(图2)。总体上,βG的根际酶活性热点区范围是0.93—1.98 mm,NAG的根际酶活性热点区范围是0.59—1.86 mm(图2和图3)。其中,碳水解酶活性热点区为苜蓿根伸长区(1.98 mm)>玉米在根尖和根伸长区(约为1.14 mm)>莎草根伸长区(0.93 mm)(图4)。氮水解酶活性热点区为莎草根尖(1.86 mm)>玉米在根尖和根伸长区(约为1.12 mm)>苜蓿根伸长区(0.59 mm)(图4)。该结果表明苜蓿根际土壤和微生物可能对能量的需求最强烈,而莎草根际土壤和微生物可能对氮养分的需求最强烈。
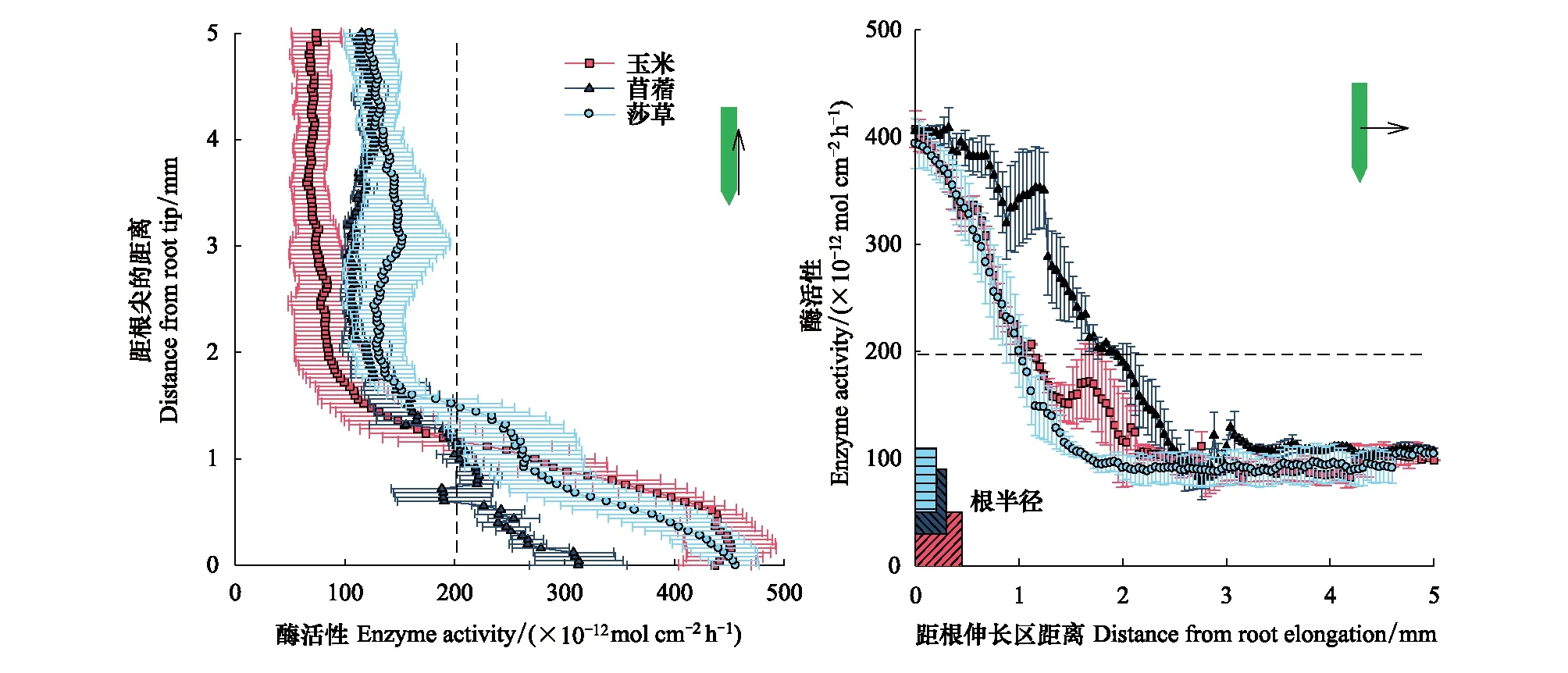
图2 不同植物根系土壤β-葡萄糖苷酶活性沿根尖向上和根伸长区向外分布Fig.2 The distribution of β-glucosidase activities from the root tip upward and the root elongation outward along different plant roots
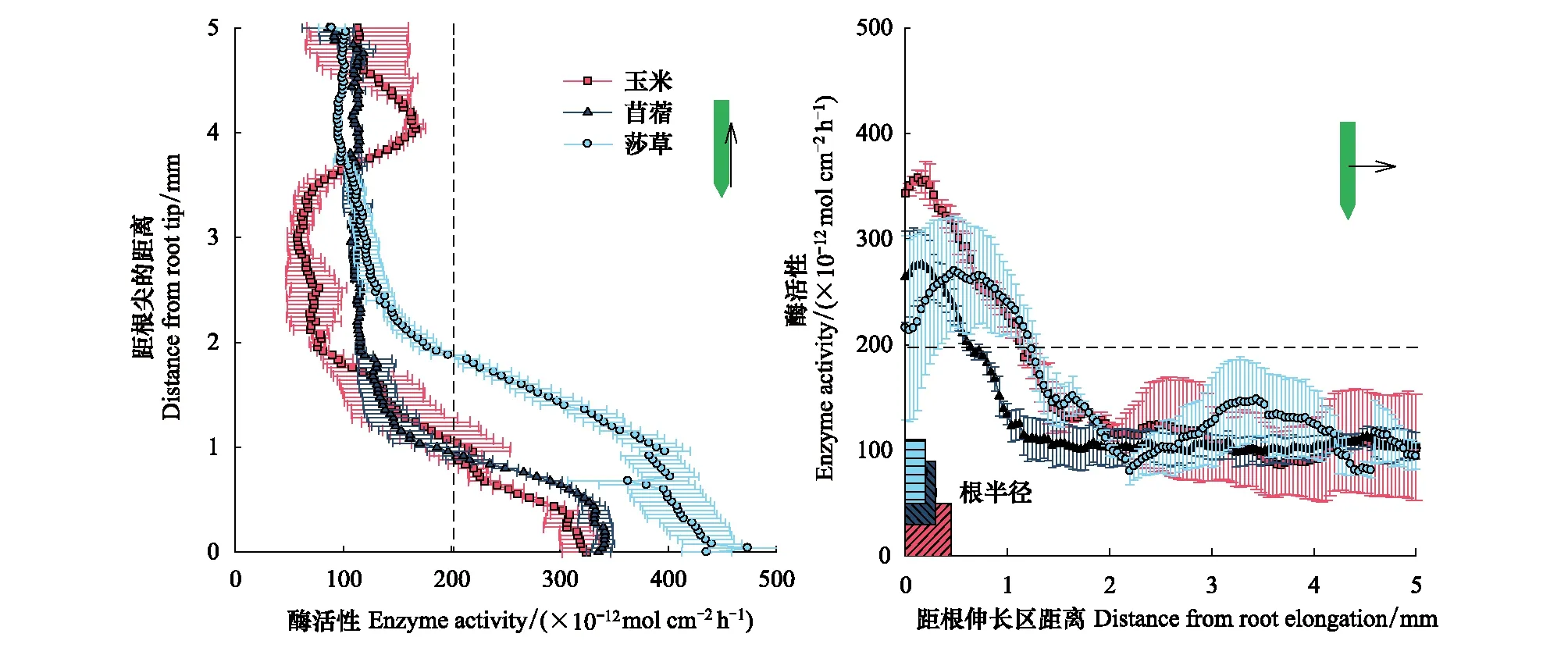
图3 不同植物根系土壤N-乙酰氨基葡萄糖苷酶活性沿根尖向上和根伸长区向外分布Fig.3 The distribution of N-acetyl glucoaminosidase activities from the root tip upward and the root elongation outward along different plant roots
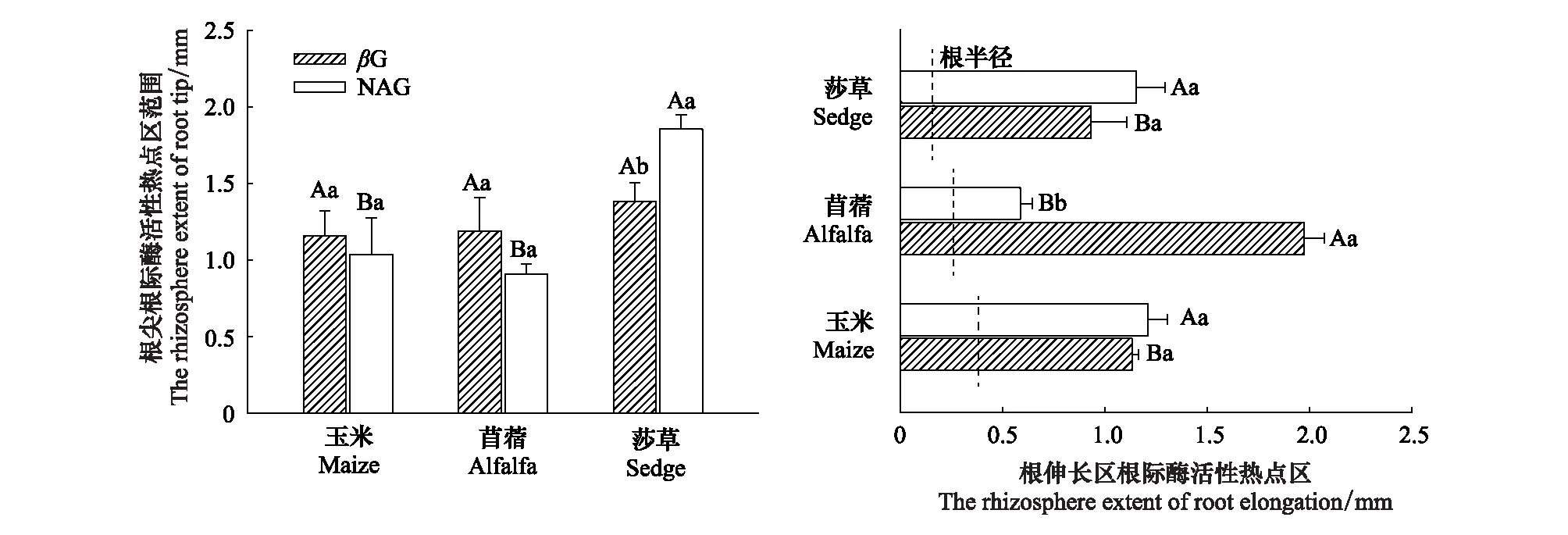
图4 三种植物根尖和根伸长区酶活性热点区的范围Fig.4 The rhizosphere extent of hotspot in root tip and root elongation zones for three plants大写字母(A、B)表示同种酶在不同植物根际酶活性热点区范围之间的显著性差异,小写字母(a、b)表示同种植物不同酶之间的显著性差异(P< 0.05 LSD检验);βG:β-葡萄糖苷酶β-glucosidase; NAG :N-乙酰氨基葡萄糖苷酶 N-acetyl glucoaminosidase
玉米碳氮水解酶活性热点区在根尖与根伸长区范围相近(1.13 mm左右);而苜蓿在根不同部位对能量和氮养分的需求不同,体现为βG根际酶活性热点区范围是根伸长区(1.98 mm)>根尖(1.19 mm),NAG根际酶活性热点区范围为根尖(0.91 mm)>根伸长区(0.59 mm);莎草根不同部位对能量和氮养分的需求一致为βG、NAG根际酶活性热点区范围均是根尖(1.38—1.86 mm)>根伸长区(0.93—1.16 mm)(图5)。该结果表明3种植物根际土壤和微生物对能量和氮养分的需求不同,玉米对碳氮需求一致,而苜蓿根尖对氮的需求大,在根伸长区对碳的需求大;莎草则是对氮养分的需求更大(图5)。
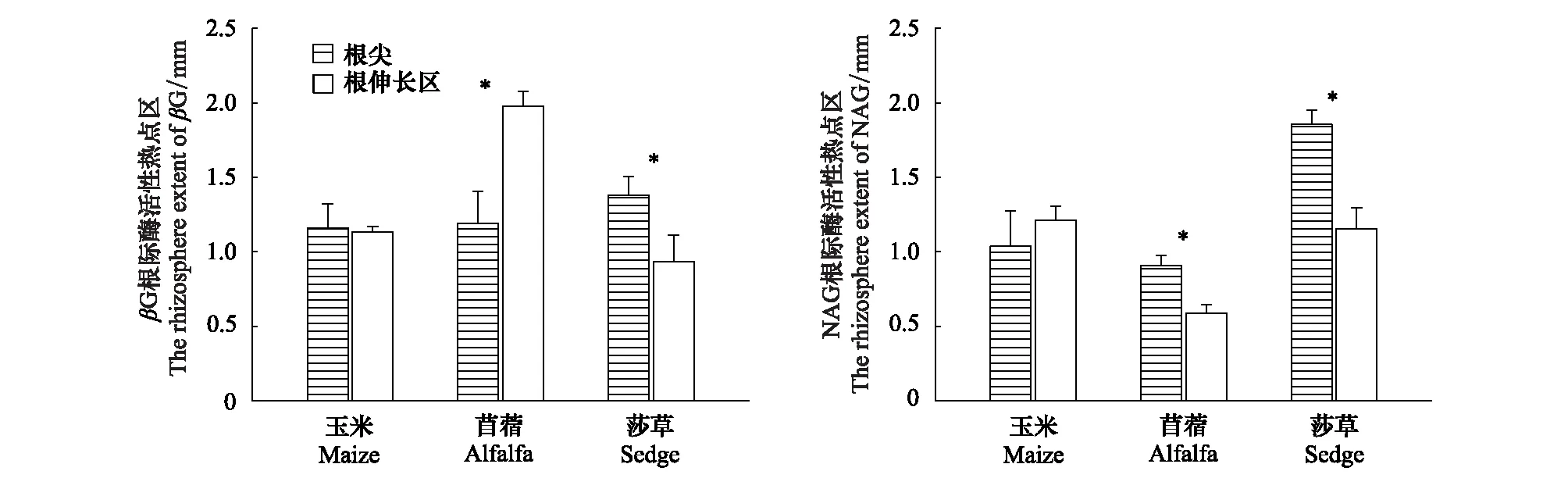
图5 三种植物根际β-葡萄糖苷酶和N-乙酰氨基葡萄糖苷酶活性热点区的范围Fig.5 The rhizosphere extent of β-glucosidase and N-acetyl glucoaminosidase activity for three plants*表示同种酶在根系不同部位根际酶活性热点区范围之间的显著性差异(P< 0.05 LSD检验)
2.2 植物根系或根际微生物对土壤碳氮偏好
玉米根际的根系或土壤微生物对土壤碳和氮偏好度之间无差异(0.8左右)(P>0.05)(图6)。苜蓿根际的根系或土壤微生物对碳偏好(0.85)>氮(0.34)(P<0.05)(图6)。莎草根际的根系或土壤微生物对氮偏好(0.82)>碳(0.62)(P<0.05)(图6)。该结果同样表明了玉米根际的根系或土壤微生物对土壤碳、氮偏好无差异,苜蓿则偏好碳,莎草更偏好氮(图6)。
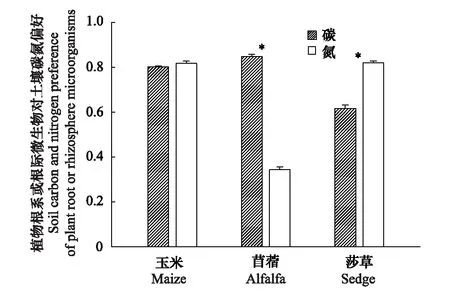
图6 植物根系或根际微生物对土壤碳氮偏好 Fig.6 Soil carbon and nitrogen preferences for plant root or rhizosphere microorganisms *代表同种植物β-葡萄糖苷酶和N-乙酰氨基葡萄糖苷之间的显著性差异(P< 0.05LSD检验)
3 讨论
3.1 植物根尖和根伸长区碳氮水解酶活性的根际酶活性热点区范围
与假设1不同的是3种植物的碳氮水解酶最大根际酶活性热点区范围为是苜蓿>莎草>玉米。虽然在相同的培养条件下,玉米根系半径(0.45 mm)最粗、且侧根数量远远大于同样是须根系的苜蓿和根系较少的直根系莎草,但是Frank等的研究认为玉米侧根多,扩大了根系在土壤中的空间分布范围和对土壤接触的总面积。这不仅增加了植物根系对土壤水分和养分的吸收,而且使得玉米比其他植物根系吸水和养分的效率更高[16],因此可能降低了根系和根际微生物通过酶分解有机质获得碳、氮养分的需求。而苜蓿和莎草根系与土壤接触面积小,需要借助土壤酶分解有机质,间接获取碳、氮维持自身生长,所以根际土壤酶分布范围较广[17]。另外,Zhang等的研究发现,当玉米与豆科植物共同种植时,玉米的根生物量和养分利用效率提高,原因就是豆科根际土壤水解酶含量多、分布广,供给植物和微生物的有效养分含量高,促进植物生长[18]。这也说明根系作为植物生长发育所需的矿质元素的主要吸收器官,其性状和功能都具有高度的可塑性[19]。在有效养分贫瘠的喀斯特土壤中,豆科植物根际酶活性热点区的范围最大,与玉米一起种植可能提高二者的养分利用效率。
与假设2一致的是莎草βG和NAG的根际酶活性热点区范围均是根尖>根伸长区,这是因为植物根系分泌物主要在根尖[20]。根际土壤微生物的生长和活性主要受分泌物的限制,所以根尖土壤微生物数量和代谢活性往往较高[21]。根系在穿透土壤过程中大量的细胞液和有机酸被释放出来[22],供土壤微生物吸收的小分子矿物质元素含量丰富[23],因此根尖土壤微生物活性高,βG和NAG根际酶活性热点区范围在根尖>根伸长区。
与假设2部分一致的是本研究发现苜蓿NAG根际酶活性热点区范围为根尖>根伸长区,但是βG根际酶活性热点区范围却相反,为根尖<根伸长区,这与Razavi等发现扁豆根际酶活性热点区范围NAG在根尖和根伸长区无显著差异不同[5]。因为豆科植物根系的根瘤多分布在根伸长区[24],为了满足微生物固氮所需的能量,根系和根际微生物分泌碳水解酶较多,这可能是根伸长区的βG根际酶活性热点区分布范围大于根尖的原因。相对地,根尖附近的土壤微生物受土壤氮养分限制[25],微生物分泌更多的NAG酶矿化有机氮,以满足根系和根际微生物对氮养分的需求,所以根尖的NAG根际酶活性热点区范围较大。
与假设2不一致的是玉米βG和NAG根际酶活性热点区在根尖和根伸长区范围相近。可能是因为玉米侧根发达,侧根之间的分泌物相互影响降低根尖和根伸长区的空间异质性[26],所以根尖和根伸长区水解酶根际酶活性热点区范围差异不显著。
3.2 植物根系或根际微生物对土壤碳氮偏好
与假设3一致的是苜蓿根际对碳的偏好大于氮,这与Razavi等发现扁豆根系对碳的需求高于对氮的需求一致[5],与土壤微生物分泌酶的经济学原理一致,即苜蓿根系通过与根瘤菌共生来满足对氮的需求,那么根际土壤分泌的酶则优先满足其他限制性因素(如碳)的需求[27]。另外,莎草根际对氮的偏好大于碳,这主要受喀斯特坡耕土壤氮养分限制影响[25]。土壤有效氮含量低,莎草根系和根际微生物分泌大量氮水解酶来获取氮养分[23]。但是研究发现玉米根际对碳氮偏好相近,主要受玉米根分泌物影响。Benizri等发现玉米根系分泌物能强烈刺激土壤微生物生长,改变微生物进化,使得根际微生物能高效利用分泌物中的养分,实现根系与微生物之间的高利用效率[28]。
4 结论
在喀斯特农田土壤中,玉米、苜蓿和莎草3种植物根系和根际土壤微生物对能量和氮养分的利用策略不同。玉米碳氮水解酶根际酶活性热点区范围相近且根际的碳氮养分偏好一致。莎草氮水解酶的根际酶活性热点区范围大于碳水解酶,根际偏好氮养分需求。而苜蓿碳水解酶的根际酶活性热点区范围大于氮水解酶,根际偏好碳需求。因为喀斯特退耕地土壤氮养分贫乏,退耕后种植苜蓿可以缓解氮需求,有助于喀斯特生态恢复。但是不同植物之间相互作用能否提高植物的养分利用效率、对生态环境的影响以及植物根分泌物对根际关键微生物和功能基因的影响尚未可知,本研究的土壤原位酶谱法为在根际尺度上微生物机制研究提供先进技术。
