珍稀濒危树种峨眉含笑种群结构与动态特征
2020-08-19秦爱丽马凡强许格希史作民陈其勇
秦爱丽,马凡强,许格希,史作民,3,*,陈其勇
1 中国林业科学研究院森林生态环境与保护研究所,国家林业和草原局森林生态环境重点实验室, 北京 100091 2 四川米亚罗森林生态系统定位观测研究站, 阿坝州 623100 3 南京林业大学南方现代林业协同创新中心, 南京 210037 4 四川省沐川县沐川国有林场, 乐山 614500
种群是指在同一时期内占有一定空间的同种生物个体的集合,是物种存在的基本单位[1],是联系个体、群落和生态系统之间的纽带[2]。种群结构是种群的最基本特征,既能反映种群不同年龄个体的数量和配置状况,也能反映种群与环境间的相互关系[3- 4]。种群动态是研究种群数量在空间和时间上的变动规律[1]。种群结构与动态是种群生态学研究的核心内容[5- 6],对分析种群现实特征和预测其未来变化趋势具有重要意义。
峨眉含笑(MicheliawilsoniiFinet et Gagn.)是木兰科(Magnoliaceae)含笑属(Michelia)常绿乔木,高可大于20 m,为中国特有种,自然种群较小,被列为国家二级重点保护野生植物[7]。该树种分布范围狭窄,其自然种群分布于中国四川省的中部和西部的峨眉、沐川、洪雅和平武等地,分布海拔600—2000 m。该树种树形优美,枝干笔直挺立,叶色亮绿,四季常青,花重瓣,乳黄色,有芳香,是园林绿化的新兴树种。目前对峨眉含笑的研究多集中在该树种的遗传多样性研究[7]、形态解剖和生理特性研究[8- 11]、引种栽培研究[12]和开发利用研究[13]等,关于该树种种群结构与动态的研究尚未见任何报道。鉴于此,本文通过在峨眉含笑天然种群的不同分布地点设置监测样地,进行实地调查,比较研究不同分布点峨眉含笑的种群情况。首次分析其天然种群目前生存状态,揭示其种群的年龄结构特征,阐明其种群更新与维持机制,为峨眉含笑种群的更新、保育和恢复提供科学参考。
1 研究区概况
研究地区位于四川省峨眉山市峨眉山周边地区、沐川县国有林场和平武县宽坝林场,覆盖了现存已知的峨眉含笑天然种群的集中分布区。
峨眉山市位于乐山市西部,四川盆地西南边缘,地理坐标为东经103°10′30″—103°37′10″,北纬29°16′29″—29°43′11″。该地区常年温暖、湿润和多雨,日照少、多云雾,属于亚热带和温带气候,自然植被为亚热带常绿阔叶林和针叶林,土壤类型多为黄壤土和红壤土[14]。沐川县国有林场,地理位置为东经103°47′—103°49′,北纬28°29′—29°54′。该区域内气候类型为亚热带湿润气候,全年日均温12.8 ℃,全年降水量1780 mm,林区内空气潮湿,年均相对湿度在90%,全年日照539 h。土壤以黄壤和黄棕壤为主[15]。平武县宽坝林场,地理坐标为东经104°19′,北纬 32°12′,在纬度上属亚热带的北缘,年均温度为11 ℃,1月份均温为1 ℃,7月份均温为20 ℃,≥10 ℃的年积温为4793℃,年降雨量为1187 mm,相对湿度为88%,日照时数达691 h,研究区植被类型为山地常绿阔叶林和山地落叶阔叶林,土壤以黄壤和黄棕壤为主[16]。
2 研究方法
2.1 样地设置与调查
2018年9—10 月,在四川省峨眉山市峨眉山、沐川县国有林场和平武县宽坝林场3个现存峨眉含笑天然分布区域,选择峨眉含笑分布较为集中的地段设置监测样地,根据地势和地形,样地面积为300 m2或400 m2,共设置了10块样地。每个样地内设置3或4 个10 m×10 m 的样方,调查并记录所有样方内峨眉含笑的个体数、基径(D,高度<1.3 m)、胸径(DBH, 高度≥1.3 m)、高度和冠幅等指标。同时测定和记录各样地的经纬度、海拔、坡度和坡向(表1)。
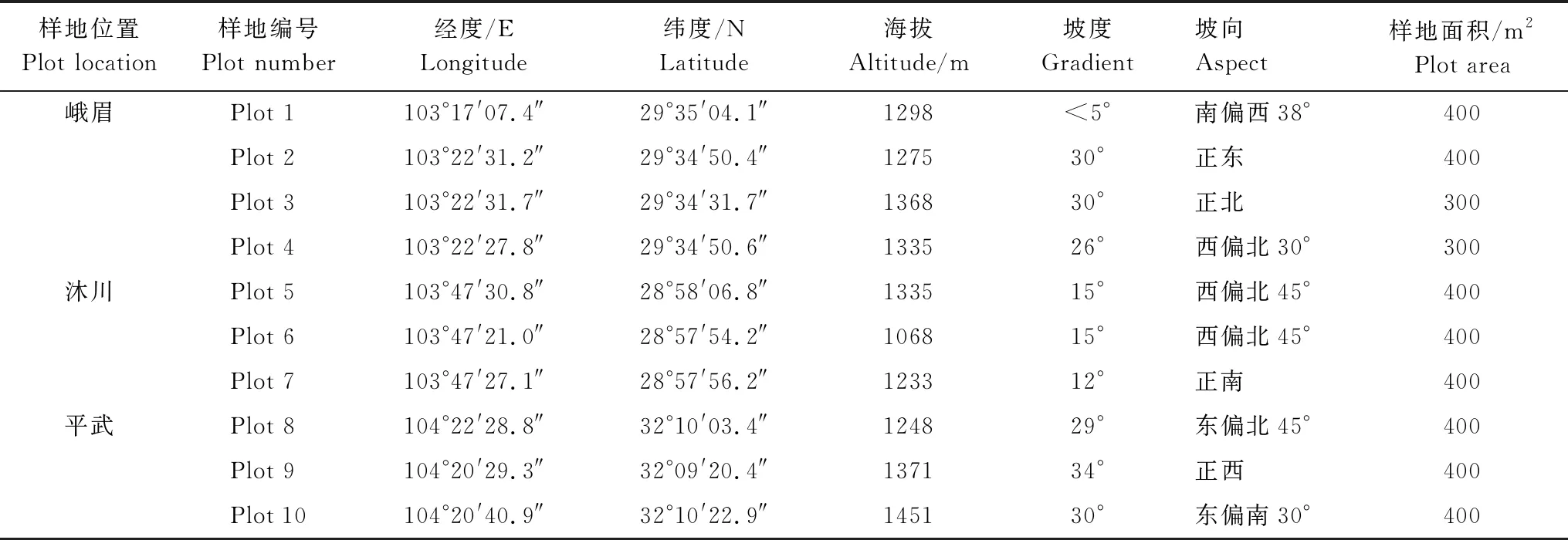
表1 研究样地概况
2.2 种群径级划分
基于同一树种的龄级和径级对相同环境的反应规律具有一致性的研究[17],针对峨眉含笑数量较少、年龄获取存在一定难度的特点,采用径级代替龄级分析其种群年龄结构。根据峨眉含笑的生长特点,参照曲仲湘对树木龄级的划分标准[18],将峨眉含笑种群划分为11个径级,基径(D)≤1 cm为第 1龄级;1 cm
2.3 静态生命表编制
静态生命表是用特定调查时间收集的种群所有个体的径级数据编制而成的[19]。由于静态生命表反映的是多个世代重叠的年龄动态历程中的一个特定时间,而不是对某一种群全部生活史的追踪[20- 21],且调查过程中也会存在系统抽样误差,导致在生命表的编制中有可能会出现死亡率为负的情况,这与生命表分析中的一些数学假设不符[22]。对此,多数学者均采用匀滑技术[23- 25]对统计数据进行处理。本研究参照江洪[23]在云杉种群生态学中所用的方法对峨眉含笑的初始调查数据进行匀滑处理,进而编制峨眉含笑的种群静态生命表。
2.4 存活曲线、死亡率曲线、消失率曲线的绘制及拟合
存活曲线的绘制方法一般有两种[26],一是以存活量的对数值lnlx为纵坐标,以年龄为横坐标作图;另一种是用存活数量对年龄作图,但年龄用平均寿命期望的百分离差来表示。本研究所绘制的存活曲线是以径级相对龄级为横坐标,以lnlx为纵坐标获得的。采用Hett和Loucks[27]提出的指数函数(Deevey-Ⅱ型:Nx=N0e-bx)和幂函数(Deevey-Ⅲ型:Nx=N0x-b)数学模型进行存活量的对数值lnlx和龄级关系的拟合,依据决定系数、F检验值来判定模型拟合效果。方程式中N0和Nx分别代表种群形成初期和第X 龄级内存活数的个体数,b为死亡率。
以径级相对龄级为横坐标,以各龄级的死亡率(qx)及损失度(Kx)为纵坐标作死亡率和消失率曲线。
2.5 种群年龄结构及动态分析
采用陈晓德[28]的种群结构分析方法量化峨眉含笑的种群结构动态,反映种群或相邻龄级间个体数量的动态关系[29- 30]。量化指数及公式如下:
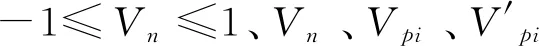
2.6 种群空间结构分析
种群的空间结构一般分为3种类型,即随机分布、均匀分布和集群分布[1]。为简化不同龄级立木的格局分析,本研究参照曲仲湘对树木龄级划分不同发育阶段的方法[18],结合实地考察中对该树种的观察和记录,将峨眉含笑的不同龄级划分成6个发育阶段,第Ⅰ龄级,为幼苗阶段;第Ⅱ龄级,为幼树阶段;第III龄级,为小树阶段;第IV—Ⅵ为中树阶段;第Ⅶ—Ⅸ为大树阶段;第Ⅹ和Ⅺ为老树阶段。对分布于不同区域的峨眉含笑种群在不同发育阶段上进行了空间结构分析。所采用的指标包括方差/均值比率法、Cassie 指标法(Ca)、平均拥挤度法(M*)以及丛生指数法(I);并通过负二项式指数(K)和聚块性指数(P)来分析分布格局的聚集程度[31]。
3 结果与分析
3.1 种群年龄结构
种群年龄结构能清晰地反映种群的生存状态[32],对了解种群历史,分析、预测种群动态具有重要价值[1]。本研究共调查峨眉含笑植株181株,其中峨眉山55株,种群密度为0.0393株/m2,种群年龄结构不完整,缺乏第Ⅲ、Ⅳ和Ⅵ龄级的个体,幼苗、幼树、小树、中树、大树和老树比例分别为58%、20%、0%、4%、11%和7%(图1),幼苗和幼树的比例较高,比例之和达到78%,大树和老树所占比例较小,仅有18%,种群表现为增长型;沐川县国有林场调查范围内仅有15株,种群密度最低,为0.0125株/m2,种群年龄结构不完整,缺乏第Ⅱ、Ⅲ、Ⅴ和Ⅹ龄级的个体。种群中个体集中分布在大树阶段,比例为53%,幼苗和中树比例分别为13%和14%,无幼树和小树,老树比例为20%(图1),种群中大树和老树比例偏高,种群表现为衰退型;平武县宽坝林场111株,种群密度最高,为0.0925株/m2,其幼苗比例最高,为32%,幼树、小树、中树和大树的比例差不多,老树比例偏低(图1),种群表现为增长型。
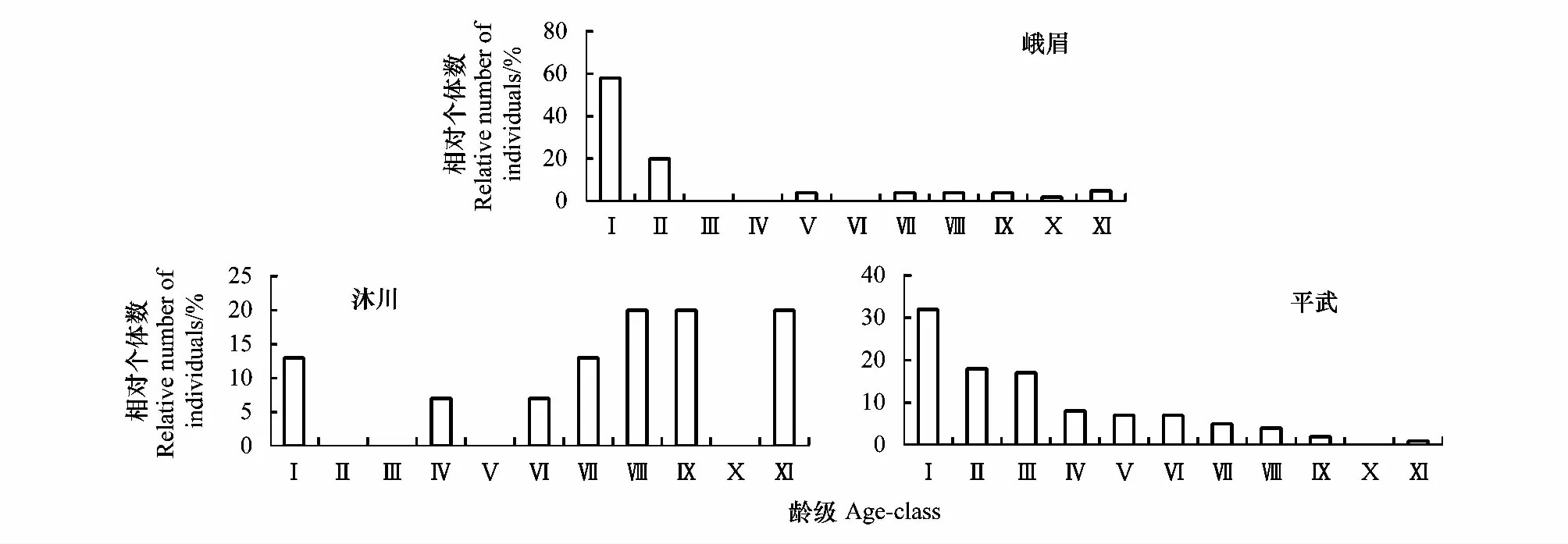
图1 不同分布区域峨眉含笑种群的年龄结构Fig.1 Age structure of M. wilsonii populations distributed in different regions Ⅰ:基径≤1cm;Ⅱ:1 cm<基径≤2.5 cm;Ⅲ:2.5cm<胸径 ≤ 7.5 cm;Ⅳ:7.5 cm<胸径≤12.5 cm;Ⅴ:12.5 cm<胸径≤17.5 cm;Ⅵ:17.5 cm胸径 ≤ 22.5 cm;Ⅶ:22.5 cm胸径 ≤ 27.5 cm;Ⅷ:27.5 cm<胸径≤32.5 cm;Ⅸ:32.5 cm<胸径≤37.5 cm,Ⅹ:37.5 cm<胸径≤42.5 cm;Ⅺ:胸径>42.5 cm
3.2 静态生命表与存活曲线、死亡率曲线和消失率曲线
对峨眉含笑原始调查数据进行匀滑修正[23]后编制的种群静态生命表见表2。
由表2知,随着年龄增加,标准化后存活个体数(lx)逐渐减小,个体的期望寿命在幼树、小树和中树阶段较大。具体表现为峨眉种群种群的静态生命表在小树和中树(Ⅲ龄级和Ⅳ、Ⅴ龄级)阶段生命期望值突然升高,之后再降低,说明峨眉含笑在小树和中树阶段生存能力较强,推测小树和中树阶段个体经历了幼苗期、幼树期的自然筛选与种内、种间竞争后存活下来的个体具有较强的生命期望寿命。沐川和平武种群的静态生命表在幼树阶段(第Ⅱ龄级)就表现了较强的生命期望值,之后生命期望在小树和中树阶段也较高,大树和老树阶段开始降低。
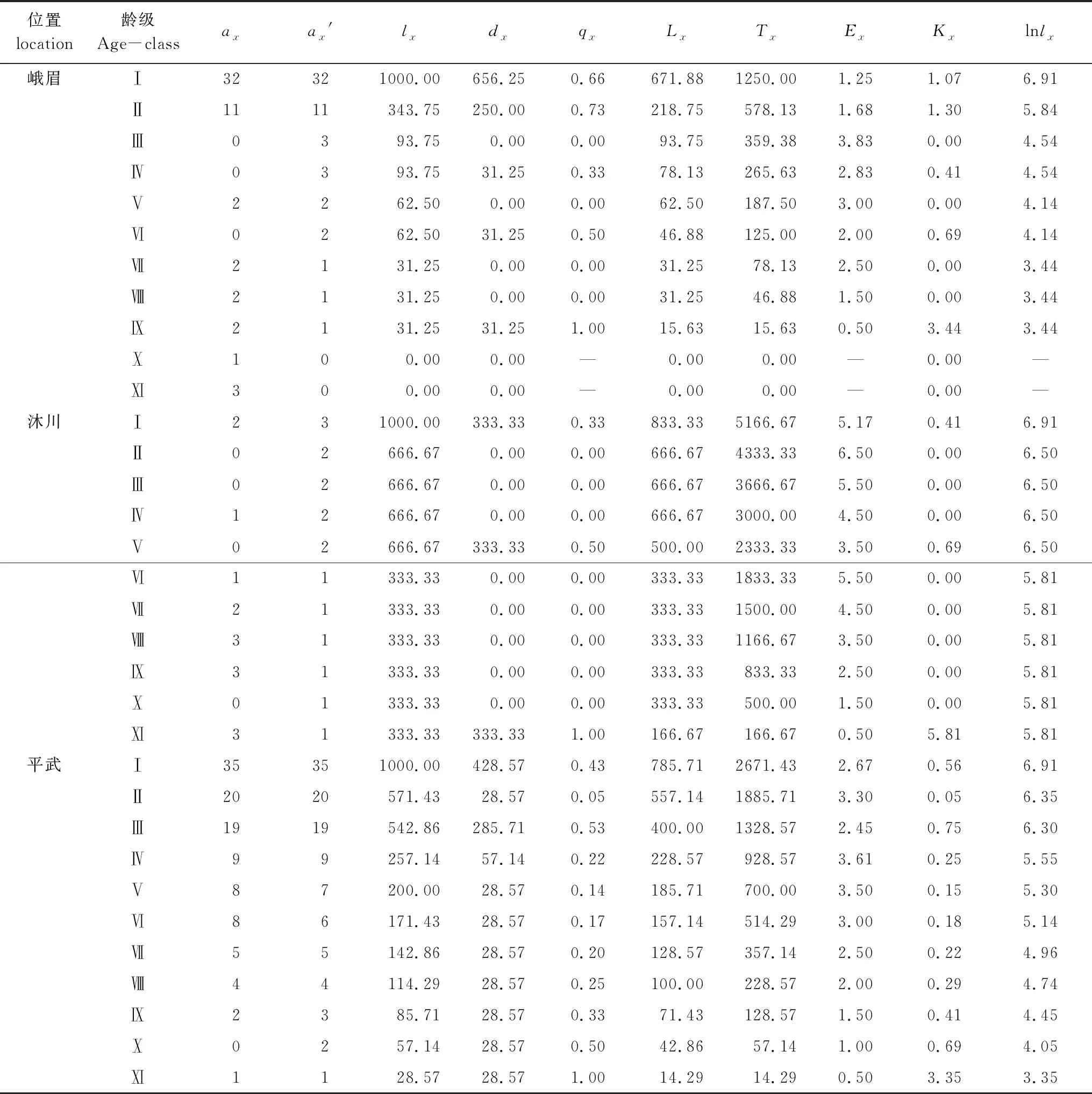
表2 峨眉含笑种群静态生命表
存活曲线是反映种群个体在各年龄级的存活状况曲线[26],可直观的表达种群个体在各龄级的存活过程[1, 5, 33]。自Pearl[34]首次提出了种群存活曲线后,Deevey[35]曾对存活曲线划分为3种类型:即Ⅰ型(曲线凸型),表示幼体存活率高,老年个体死亡率高,在接近生理寿命前仅有少数个体死亡;Ⅱ型(曲线对角线型),各龄级死亡率基本形同;Ⅲ型(曲线凹型),幼体死亡率很高。峨眉种群前期死亡率较高,后期较低(表2和图2),属于Deevey-Ⅲ曲线特征,存活曲线近似“凹”型(图2),模型检验亦支持其为Deevey-Ⅲ型(表3);沐川种群幼苗期和小树期死亡率较高(表2,图2),经模型检验与Deevey-Ⅲ型有较高的相关系数(表3),因而判断沐川种群的存活曲线亦为Deevey-Ⅲ型;平武种群各龄级的死亡率基本相同(表2和图2),存活曲线近似对角线型(图2),且模型检验结果亦为Deevey-Ⅱ型(表3)。
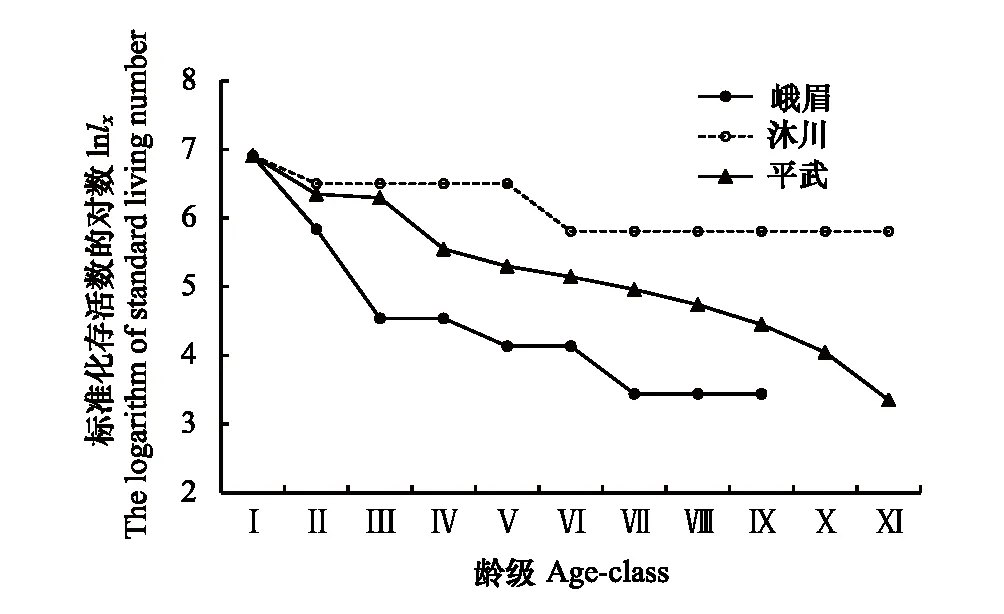
图2 峨眉含笑种群存活曲线Fig.2 Survival curve of M. wilsonii population
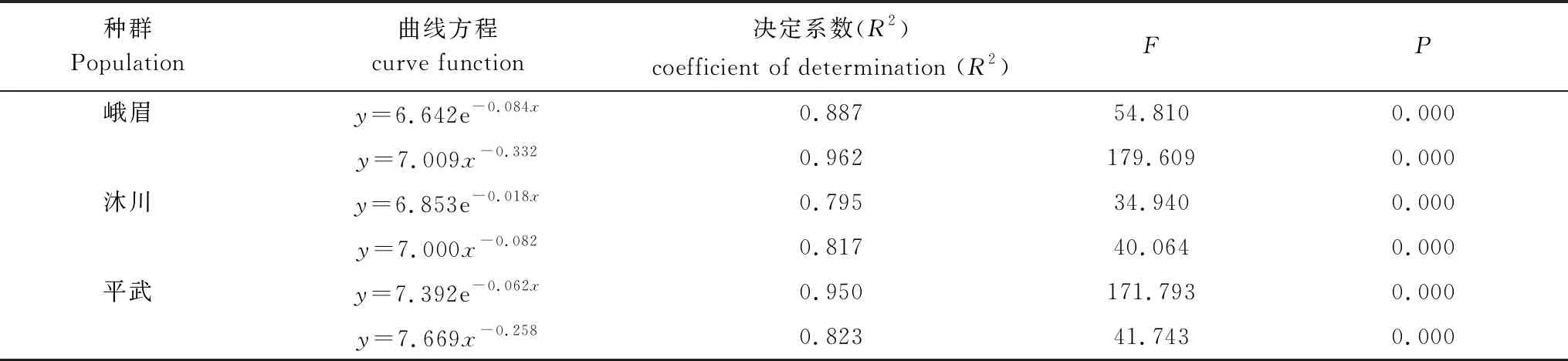
表3 峨眉含笑种群存活曲线的检验模型
死亡率曲线反映的是不同分布点峨眉含笑种群死亡率的动态变化。由图3可以看出:同一分布点峨眉含笑种群死亡率和消失率的变化趋势基本一致,不同分布点峨眉含笑种群死亡率和消失率的变化不同。峨眉种群死亡率在幼苗期和幼树期的死亡率偏高;说明峨眉种群在幼苗和幼树期就经历了环境筛的强烈筛选。沐川种群中树(第Ⅴ龄级)的死亡率最高,说明从小树向中树第Ⅴ龄级过度过程中,随着峨眉含笑径级的增加,种内或者种间对资源如光照、水分等的需求加剧,致使大量个体死亡。平武种群在小树(第Ⅲ龄级)的死亡率最高的原因除了该价段对资源的需求加剧之外,更可能的因素是人类活动的干扰。实地调查中发现,平武宽坝林场人为对生境的破坏严重,乱砍乱伐现象时有出现,导致大量幼树、小树没有成林就被砍伐。
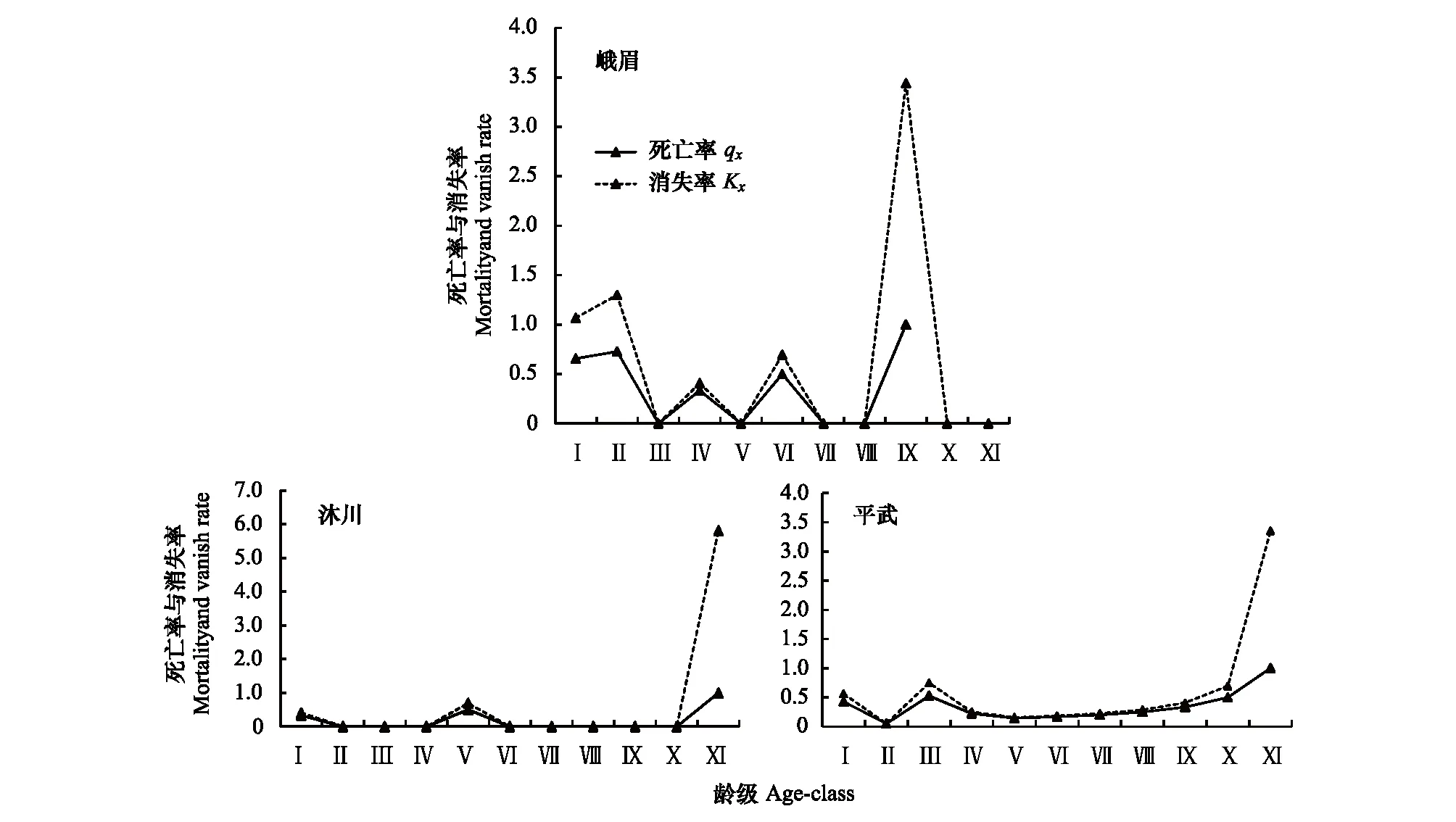
图3 峨眉含笑种群死亡率与消失率曲线Fig.3 Mortality and vanish rate of M. wilsonii population
3.3 种群年龄结构的动态变化分析
基于种群动态量化分析方法,对峨眉含笑种群年龄结构动态指数的分析结果见表4。

表4 峨眉含笑种群年龄结构动态指数/%
由表4知:峨眉种群在第Ⅰ、Ⅱ、Ⅴ和Ⅸ龄级种群数量动态指数分别是66、100、100和50,均大于0,呈现增长的结构动态,说明峨眉山的环境条件比较适合该树种从第Ⅰ-Ⅱ和第Ⅴ、第Ⅸ龄级的生长。从第Ⅲ向第Ⅳ龄级以及从第Ⅴ到第Ⅵ龄级过渡时种群数量动态系数小于0,呈现衰退的结构动态,说明环境筛在该龄级内存在。沐川种群从Ⅱ龄级向第Ⅲ龄级和从第Ⅴ和第Ⅵ龄级向第Ⅶ龄级过渡时动态指数均小于0,呈现衰退的结构动态,说明沐川分布区环境对幼树、小树和中树的生长压力大,该阶段存在严重的环境筛。平武种群数量动态指数在第Ⅴ龄级等于0,其他阶段(除老树正常死亡)均大于0,说明平武的环境比较适合该树种的生长。
种群总体动态变化指数分析结果显示:峨眉(Vpi= 0.57)、平武(Vpi= 0.31)和沐川(Vpi= 0.16)种群总体变化动态指数Vpi均大于0,种群均为增长型。但由于沐川种群总体动态变化指数仅为0.16,且由于种群年龄结构极不完整,显著缺乏幼树和小树,大树和老树比例又明显偏高,因此沐川种群应为衰退型种群。
3.4 峨眉含笑种群空间分布格局
峨眉含笑种群在不同龄级的空间分布格局见表5。由表5知,除峨眉种群小树和沐川种群幼树和小树发育阶段由于缺乏个体表现为随机分布外,峨眉和沐川种群在不同龄级上均呈聚集分布。平武种群的中树和大树呈均匀分布,其他龄级的空间分布格局也呈聚集分布。从聚集强度看,峨眉种群的中树、沐川种群的幼苗和中树、平武种群的老树聚集程度最大。

表5 峨眉含笑种群的空间分布格局
4 讨论
4.1 峨眉含笑种群结构特征
植物种群结构不仅能体现植物种群的动态变化规律,同时也是植物对立地环境条件长期适应的反映。在当前演替阶段,峨眉和平武种群年龄结构属增长型,沐川种群属衰退型,这与种群量化动态分析的结果基本一致。沐川种群衰退的主要原因可能是林场内林分密度和上层林冠郁闭度过大,导致林内光线不足,林下阴暗潮湿,致使大量种子腐烂,丧失发芽能力。另外,林场内林下大部分区域方竹生长茂密,盖度可达80%,方竹根系发达,占据了林下大部分的空间,限制了峨眉含笑种子的萌发,致使幼苗比例偏低,郁闭度大的阴暗潮湿的生境阻碍了幼苗的生长,导致沐川国有林场内峨眉含笑幼苗极少能发育成幼树和小树,种群年龄间存在断点,缺乏幼树和小树,导致种群呈现衰退状态。已有研究表明,幼苗和幼树阶段是森林更新过程中最敏感和最重要的阶段[36- 37],有限的资源和空间是限制种群更新的关键因子[38- 39, 25]。已有多个树种如海南龙血树[30]、羽叶丁香[26]、紫果云杉[25]的研究表明,幼苗和幼树不足是导致种群衰退或种群内局部龄级呈衰退现象的原因之一。
本研究中峨眉种群和平武种群虽是增长型,但峨眉种群动态量化分析显示局部龄级间仍存在衰退现象。峨眉种群缺乏小树,说明幼树向小树转变的过程中存在巨大的环境压力,其次在中树阶段,龄级间呈衰退状态,推测该阶段亦存在巨大的环境选择压力,中树长到一定高度后,空间、营养等有限资源的限制,致使部分竞争力弱的中树无法向下一级林木大量转化进入林冠层。平武种群虽呈增长型,但其幼树阶段的种群动态系数很低,且我们在实地调查中,发现平武种群存在大量的峨眉含笑实生幼苗,这是在其他两个种群未能发现的,但该龄级的年龄结构动态指数却仅为5,呈现衰退的结构动态,说明该环境下幼苗向幼树的转变过程中存在瓶颈,或者幼苗转换成幼树后存在人为活动的干扰,导致动态指数急剧下降,在实地调查中发现部分峨眉含笑被砍伐后,其原生地被栽植成其他经济树种厚朴(Magnoliaofficinalis)或者农作物的现象时有发生,人为活动干扰较多。
4.2 峨眉含笑种群的动态趋势
从生命表及相关的存活曲线、死亡率曲线、消失率曲线和种群动态量化分析看出,峨眉种群前期幼苗和幼树期死亡率较高,突破该瓶颈后,种群死亡率就一直降低,直到老树阶段,但种群由于缺乏小树抗外界干扰能力也一般。沐川种群在逐步衰退,不仅种群密度极低,且维持种群数量的中树的死亡率也最高,种群由于缺少幼树和小树其结构极不完整,种群现状堪忧。平武种群呈现从增长型向稳定型过度的迹象,种群在小树阶段呈现出的死亡高峰,可能是该地种群濒危的重要原因。死亡高峰的出现除了与该树种种内竞争有关,外界人为乱砍乱伐也是一个重要原因。故针对此地我们应首先加强对峨眉含笑栖息地的保护。
4.3 不同分布点峨眉含笑种群的空间分布格局
峨眉含笑种群的空间分布格局受该树种自身的生物学特性如种群历史、繁殖方式等的影响较大。整体而言,峨眉和沐川种群均呈聚集分布,主要是与母树种子的散布方式有关。峨眉含笑的种子较大,自然成熟后,由于重力作用,种子多散落在母树周围,萌发出的幼苗、幼树均分布在母树周围,呈聚集分布。此外,这种聚集分布的式样也可能与本文在样地设置时由于物种分布生境破碎化,所选的空间尺度较小有关。平武种群中树和大树呈现均匀分布的式样,原因可能是种群内个体间的竞争所致,研究发现平武种群密度最大且种群内幼苗最充足,幼苗到幼树、小树以及小树成长为中树的阶段都是对资源竞争最激烈的阶段,因而对光照因子、土壤养分的需求加大,竞争加剧,导致呈均匀分布的格局。
4.4 峨眉含笑的保护与恢复
根据本文的研究结果,对峨眉含笑不同分布地点种群的保护应采取不同的措施。对于幼龄死亡率高的峨眉种群可以采用人工辅助措施获得该种群的实生幼苗,待幼苗长成小树后将小树移植到该种群的野外分布区。对于林分密度和郁闭度大的沐川种群,宜利用人工辅助措施促进种群的天然更新,如首先伐除母树和幼苗周围的乔灌木、方竹等,增加种子入土后萌发呈幼苗的可能性,并为幼苗的成长创造条件,另外鉴于此地区苗木数量不足,宜采用人工育种方式,实行就地保护,人工育种后把苗木移栽到附近山区适于该树种生长的海拔区段,加强对该树种的保护。而对于生境破坏严重的平武种群,宜制订适宜的保护栖息地的有力措施,并呼吁当地相关部门及居民加强对该树种的保护,禁止乱砍乱伐,禁止破坏栖息地的行为,并巩固和保存好现有的种群结构及生境条件,促使种群的增长潜力进一步释放,促进种群的稳步增长。
致谢: 感谢四川省峨眉山生物资源站的李策宏先生在群落和种群调查中协助进行物种的鉴定工作,感谢四川省沐川县国有林场和平武宽坝林场的员工在群落和种群调查中给予的帮助和支持, 感谢审稿人和编辑对本文提出的宝贵意见和建议。
