盐度对沙蜇有性繁殖阶段早期发育的影响
2020-08-19李玉龙刘修泽王爱勇王小林于旭光
柴 雨,董 婧,段 妍,孙 明,王 彬,李玉龙,刘修泽,王爱勇,王小林,于旭光,郭 栋
辽宁省海洋水产科学研究院,辽宁省海洋生物资源与生态学重点实验室, 大连 116023
沙蜇(Nemopilemanomurai)是世界上最大的水母之一,成体水母直径可达2 m,体重可达200 kg。自20世纪90年代末起,频繁暴发于我国的渤海、黄海、东海北部以及日本海和韩国海域[1- 7]。沙蜇暴发期间对中、大型浮游动物的摄食压力超过浮游动物的生产力,这种捕食压力间接导致鱼类的数量减少,浮游植物增多引发赤潮[8- 11]。同时,沙蜇作为我国食用水母之一,伴随着海蜇自然资源的衰退,近年来沙蜇的捕捞量已经远超海蜇,成为市场上海蜇类产品的主要来源[12]。总之,沙蜇作为东北亚海区大型水母暴发优势种之一,因其生物量大,影响范围广,自暴发起便引起了国内外学者的广泛关注。
随着学者们在沙蜇生活史[3]、生物学特征[13-14]、形态学[15-16]、海区分布生态特性[17-20]等方面的研究开展,人们认识到沙蜇水母体来源于螅状体的横裂生殖,螅状体的种群数量是水母体暴发的基础。沙蜇螅状体来源于水母体有性繁殖与螅状体无性繁殖两种途径。沙蜇水母体巨大,成熟后个体怀卵量大,有性繁殖产生新生螅状体的基数大,螅状体无性繁殖生成新生螅状体的方式单一,仅为足囊繁殖并需要适宜环境下萌发[3,14,21-23]。所以,了解沙蜇通过有性繁殖对螅状体种群的补充机制十分重要。在沙蜇有性繁殖的过程中,早期发育阶段包括受精卵发育为浮浪幼虫、浮浪幼虫经短暂的浮游生活后附着变态发育为底栖的早期螅状体,这一过程通常需要3—7日完成。由于早期发育过程复杂且发育时间短不易观察,加之沙蜇成熟水母体体积巨大、培养困难、实验材料获取困难。目前,针对沙蜇有性繁殖阶段的相关研究鲜见报道。盐度作为海区重要的环境因子,有报道称其对其他种类大型水母有性繁殖阶段浮浪幼虫附着的影响显著[24-27]。沙蜇每年春季水母体幼体在海湾河口及浅滩被发现,同年的秋季成熟水母体在沿岸聚集进行有性繁殖。虽然到目前为止仍没有找到野生的沙蜇螅状体种群,但是通过沙蜇分布移动调查表明,沙蜇的发源地为河口、浅滩海域[19-20,28]。沙蜇产卵的季节正是夏季季风期,降雨和河流径流增加都会使沙蜇有性繁殖期间处于盐度多变的生态环境下。较低和多变的盐度可能对沙蜇有性繁殖阶段的早期发育产生重要作用,从而影响螅状体种群数量的补充。本文在辽东湾海区沙蜇产卵季节捕获到成熟亲体,在室内通过人工繁育获得实验材料沙蜇受精卵,结合室内模拟生态实验,研究了盐度对沙蜇有性繁殖阶段早期发育的影响。探讨了不同盐度培养下沙蜇有性繁殖阶段受精卵发育、浮浪幼虫的附着变态及早期螅状体的存活与生长的差异。研究结果可揭示盐度对沙蜇生活史繁殖及种群补充的作用机制,为探索沙蜇暴发的生态环境条件、寻找沙蜇的种群发源地提供科学依据。
1 材料和方法
1.1 试验材料
2015年9月在辽东湾辽宁省葫芦岛兴城市海岸采用海蜇网捕捞到成熟的沙蜇水母体作为亲体。将雌雄亲体在育苗室内分开暂养,取样生殖腺在显微镜下鉴定成熟度后,将性腺成熟的雌雄亲体移入同一培养池中,6 h后,每0.5 h取样观察。受精卵直径60—80 μm,镜下可见极体。受精卵出现后用孔径25 μm的筛绢进行收集。亲体培养用水均取自辽宁省葫芦岛兴城市海岸,经黑暗沉淀并沙滤后使用。试验期间亲体培养用水盐度30—32,水温随室温波动,范围在23—26 ℃之间。
1.2 试验方法
1.2.1试验设置
盐度对受精卵阶段发育的影响、对浮浪幼虫变态附着的影响及对初生螅状体生长存活的影响实验均设置4个盐度梯度,即S15、S20、S25、S30,实验用水来源与培养用水相同,温度实验前用孔径0.45 μm微孔滤膜过滤后与去离子水混合,配好盐度的实验用水静置1 d后使用,盐度测量采用PAL-06。
1.2.2受精卵发育阶段试验
在解剖镜(Nikon SMZ745T)下使用毛细吸管选取受精卵(卵膜完整),放入直径35 mm培养皿中观察培养,培养体积10 mL。每盐度组选取的受精卵数量为10个,每组设3个平行。余下的受精卵放入装有培养用水的800 mL方形透明PVC盒中培养。试验期间培养皿加盖避免培养期间水分蒸发,用解剖镜(Nikon SMZ745T)时时观察,以个体通过纤毛开始单向运动为浮浪幼虫形成标志统计数量。
1.2.3浮浪幼虫变态发育附着阶段试验
在解剖镜(Nikon SMZ745T)下,使用吸管选取约50个囊胚期受精卵,放入直径35 mm培养皿中,培养体积10 mL。待浮浪幼虫出现后,将未发育成功受精卵的及多余浮浪幼虫吸出然后进行实验观察。每盐度组余下的浮浪幼虫数量为30个,每组设3个平行。试验期间培养皿加盖避免培养期间水分蒸发。每日用解剖镜(Nikon SMZ745T)观察浮浪幼虫变态发育行为,当观察到浮浪幼虫旋转速度下降时在培养皿中加入剪裁后的波纹板作为附着基,以浮浪幼虫一段膨大生成早期螅状体(4触手螅状体)为标志统计数量,浮浪幼虫全部附着变态或解体时试验结束。
1.2.4早期螅状体发育阶段试验
将附着有沙蜇4触手螅状体的波纹板剪裁成小片,置于尺寸为100 mm×80 mm×40 mm的方形带盖塑料盒中,加入100 mL培养用水,解剖镜(Nikon SMZ745T)下每盐度组选取30个螅状体,多余螅状体用解剖针从螅状体底部剥离,每组设3个平行,每日投喂一次轮虫(Brachionusplicatilis),每日投饵前在解剖镜(Nikon SMZ745T)下观察螅状体状态,并测量早期螅状体口柄直径,实验持续到连续三日螅状体个数不变且个体状态健康(食物消化后触手舒展,捕捉食物时反应敏捷)为止,统计存活螅状体数量。
1.3 指标测定及数据统计分析
受精卵发育到各阶段的发育率Development rate of egg(DRE,%)=Ft/Fo× 100
浮浪幼虫变态附着率 Settlement rate of planula(SRP,%)=Pt/Po× 100
存活率Survival rate (SR,%) =Nt/No× 100
相对增长率Relative growth rate(RGR,%) =(Dt-Do)Do× 100
特定生长率 Specific growth rate(SGR,%/d)=(lnDt- lnDo)t× 100
式中,Fo为试验开始时受精卵细胞个数,Ft代表各发育阶段的受精卵细胞个数,Po为试验开始时浮浪幼虫的数量,Pt为试验结束时浮浪幼虫附着变态为早期螅状体的数量,No为试验开始时早期螅状体的个数,Nt为试验结束时早期螅状体的数量。Dt、Do分别为试验开始和结束时早期螅状体的口柄直径(mm)。螅状体口柄直径(mm):口面观圆周最大水平面的直径,当口面观不为圆形时,取几何图形最大直径与最小直径的平均值。
运用单因素方差分析(One—way ANOVA)检验不同盐度对沙蜇受精卵发育率、浮浪幼虫的变态附着率及初生螅状体的存活率的影响。试验数据用平均数±标准差(Mean ± SD)表示,用Excel 2016及SPSS 24.0软件对数据进行统计分析,采用Duncan′s多重比较检验组间差异,以P< 0.05作为显著性水平。
2 结果与分析
2.1 盐度对沙蜇受精卵发育的影响
由表1可见,沙蜇受精卵细胞发育时间短暂,试验8小时后各盐度组受精卵均发育为浮浪幼虫。其中,盐度20、25、30组受精卵的细胞发育速度基本一致,三组从受精卵出现到发育至囊胚期的细胞发育率均为100%,发育到原肠胚期时盐度30试验组发育率仍为100%,盐度25及盐度20试验组的细胞发育率为96.67%但三组间差异不显著(P> 0.05),在原肠胚发育到浮浪幼虫阶段受精卵发育率开始下降,发育至浮浪幼虫后盐度20组细胞发育率最高,为56.67%,显著高于其他组。盐度30、25试验组比例为50.00%和43.33%,两组间差异不显著(P> 0.05)。
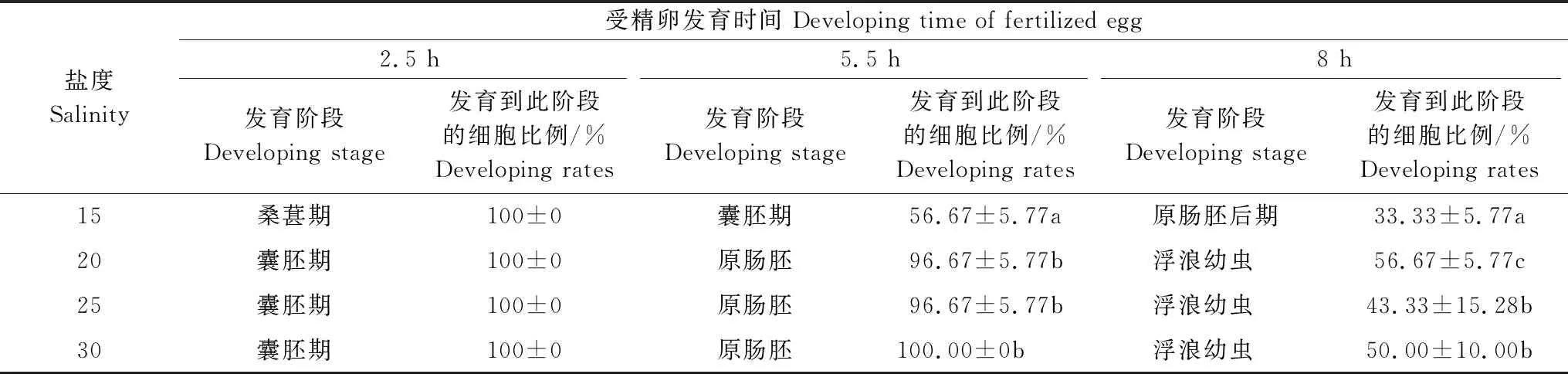
表1 不同盐度对沙蜇受精卵发育速度及发育率的影响
盐度15的试验组沙蜇受精卵发育慢于其他盐度组,发育到囊胚期的时间晚于其他盐度组,与其他盐度组不同,从受精卵至囊胚期期间试验组内的受精卵已出现细胞发育停止的现象,发育至囊胚期的发育率也显著低于其他盐度组,发育率为56.67%。试验至8 h后其他盐度组受精卵发育至浮浪幼虫,本组细胞体表出现纤毛尚未开始转动,处于原肠胚后期时,发育晚于其他盐度组,且发育率显著低于其他盐度组(P< 0.05)。
综上,沙蜇早期发育最适盐度在20左右,低盐度对受精卵发育影响大于高盐度。低盐度导致受精卵细胞发育延迟且发育率明显降低。高盐度不影响受精卵发育速度,但对发育率影响显著。
2.2 盐度对浮浪幼虫附着变态的影响
试验过程中观察到沙蜇浮浪幼虫在不同盐度培养下附着变态发育的速度不尽相同,状态也有差异。
培养后的第1天,盐度15组中浮浪幼虫一部分沉在培养皿底不动,另一部分缓慢自转且游动,状态不活泼;其他盐度组中,浮浪幼虫体型拉长,自转的同时在培养皿中游动,状态活泼。
培养后的第2天,盐度15组中浮浪幼虫均未进一步发育且前一天沉底的浮浪幼虫中一部分出现边缘溶解解体现象;盐度20、25组中观察到部分浮浪幼虫附着变态为早期螅状体,余下的浮浪幼虫自转及游动速度降低,此时在培养皿中浮浪幼虫和早期螅状体同时存在,镜下观察到附着的早期螅状体触手舒展、有刺细胞分布,胃囊内可见浮浪幼虫;盐度30组中少数浮浪幼虫附着形成锥形体,即虫体停止游动,在水中成垂直形态,一端附着,另一端膨大但尚未发育成触手。
培养后的第3天,盐度15组浮浪幼虫附着形成锥形体和早期螅状体;盐度20组附着的早期螅状体触手舒展,胃囊内可见浮浪幼虫,培养皿中已无浮浪幼虫;盐度25组附着的早期螅状体触手舒展,胃囊内可见浮浪幼虫,培养皿可见少量浮浪幼虫缓慢自转及游动;盐度30组附着的锥形体均已发育成早期螅状体,触手舒展、有刺细胞分布,胃囊内未见浮浪幼虫,同时可见培养皿中浮浪幼虫自转及游动速度下降。
培养后的第4天,盐度15组早期螅状体数量增多,同时可见培养皿中浮浪幼虫自转及游动,速度没有明显变化;盐度20组早期螅状体数量不变,触手舒展;盐度25组早期螅状体数量增多,触手舒展,培养皿中已无浮浪幼虫;盐度30组早期螅状体数量不变,触手舒展,培养皿中已无浮浪幼虫。
培养后的第5—7天,盐度15组早期螅状体数量不变,培养皿中可见浮浪幼虫缓慢自转及游动。其他盐度组早期螅状体数量不变,触手舒展,培养皿中无浮浪幼虫。
至培养后的第8天,盐度15组中早期螅状体数量相同,培养皿中浮浪幼虫全部分解消亡。
镜下观察可知,不同盐度下沙蜇浮浪幼虫附着变态发育成早期螅状体的时间为2—5 d,培养5 d后仅盐度15组有存活的浮浪幼虫但也没有再附着发育;盐度20、25组最早出现早期螅状体,触手舒展具备捕食能力;盐度15组浮浪幼虫自转及游动速度低,发育迟缓、早期螅状体出现时间最晚。
将各盐度组的浮浪幼虫附着率分析可知(图1),在培养后的第3d各盐度组的浮浪幼虫附着变态率开始出现差异,至第8天试验结束,盐度25组附着变态率最高与盐度20组差异不显著(P> 0.05),盐度15组附着变态率最低与盐度30组差异不显著(P> 0.05),盐度20、25组的浮浪幼虫的附着变态率显著高于盐度15、30组(P< 0.05)。
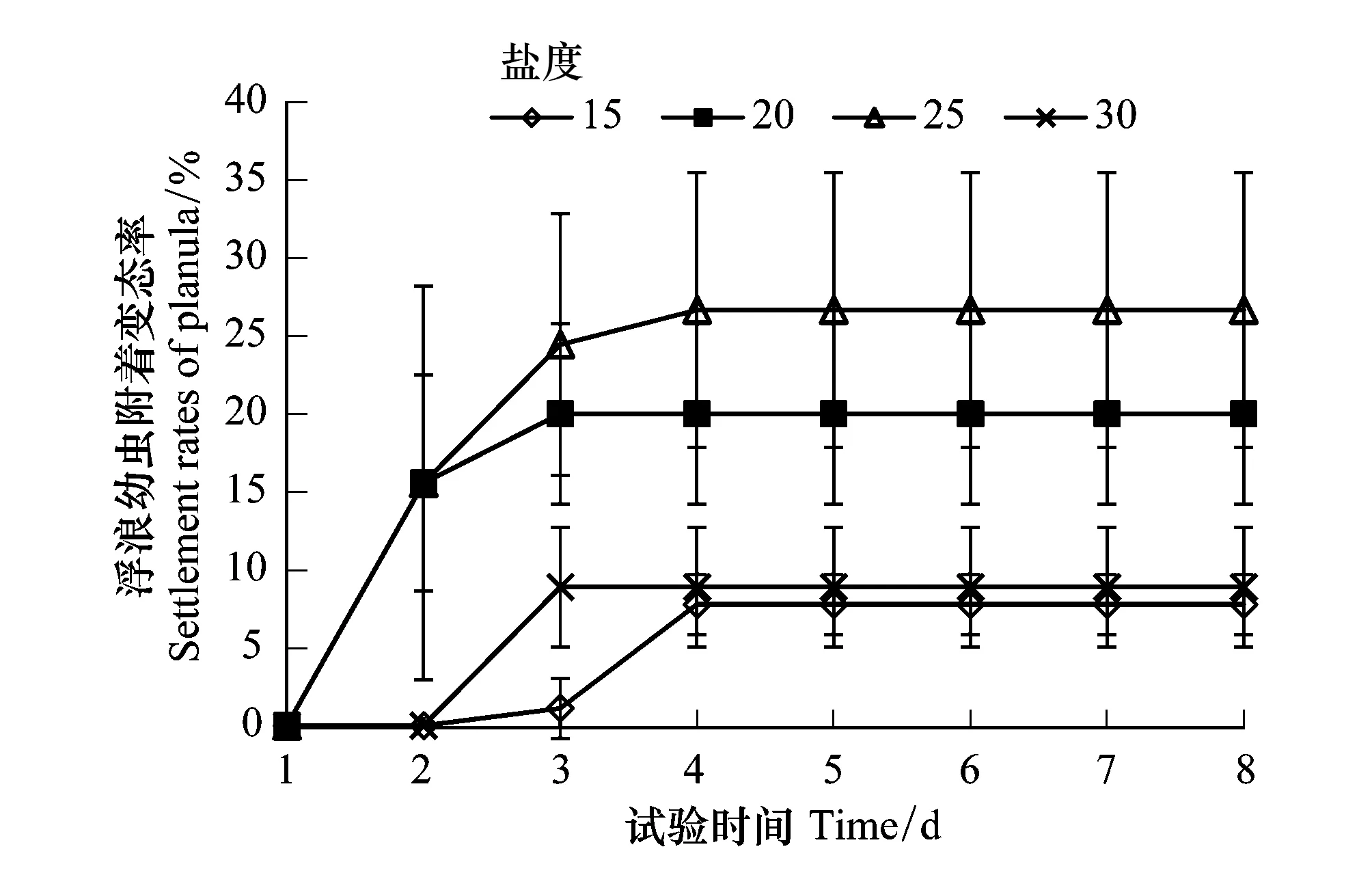
图1 不同盐度培养下的沙蜇浮浪幼虫附着变态率 Fig.1 Settlement rates of N. nomurai planula at different salinities
由图1可见,盐度20、30组在培养后的第3天浮浪幼虫的附着变态率开始稳定,盐度15、25组在培养后的第4天附着变态率开始稳定。但结合镜下观察可知,各盐度组附着变态率稳定的原因应该不尽相同。盐度20、25组培养后第2天在附着的早期螅状体的胃囊内发现浮浪幼虫,分别在第3天、4天培养皿中已无浮浪幼虫,分析两组附着变态率稳定的原因应该为浮浪幼虫被水体中的早期螅状体捕食。而盐度15组在培养后第2天起观察到浮浪幼虫的溶解解体、附着的早期螅状体胃囊内也未发现过浮浪幼虫,其附着变态率稳定的原因应该为低盐度导致的浮浪幼虫发育迟缓及停滞。
综上,本试验结果表明,适宜的盐度对沙蜇浮浪幼虫的附着变态发育及附着变态发育率十分重要。盐度20、25是沙蜇浮浪幼虫发育的适宜盐度,适宜盐度下先附着发育形成的早期螅状体已经具备捕食能力,且能捕食到水中游动的浮浪幼虫。低盐度导致浮浪幼虫活力降低、发育迟缓、溶解解体,虽然延长了浮浪幼虫的存活时间但后期存活的浮浪幼虫没有再发育附着。
2.3 盐度对早期螅状体存活及生长的影响
2.3.1盐度对早期螅状体存活的影响
试验期间,观察到健康的早期螅状体摄食前触手舒展,捕食时触手反应灵敏,摄食后触手收缩胃囊膨大,食物消化后触手恢复舒展;不健康的早期螅状体状态则相反,导致无法摄食,出现触手萎缩,口柄直径慢慢萎缩解体。试验中以早期螅状体解体为死亡标志。由图2可见,在各盐度组中沙蜇早期螅状体的存活率随着培养时间下降,其中盐度20、25、30组存活率分别为95.77%、87.97%和87.97%,三组间存活率差异不显著(P> 0.05)。盐度15组在培养后的第6天开始早期螅状体的存活率下降明显,试验结束时,存活率为14.35%,显著低于其他盐度组(P< 0.05)。
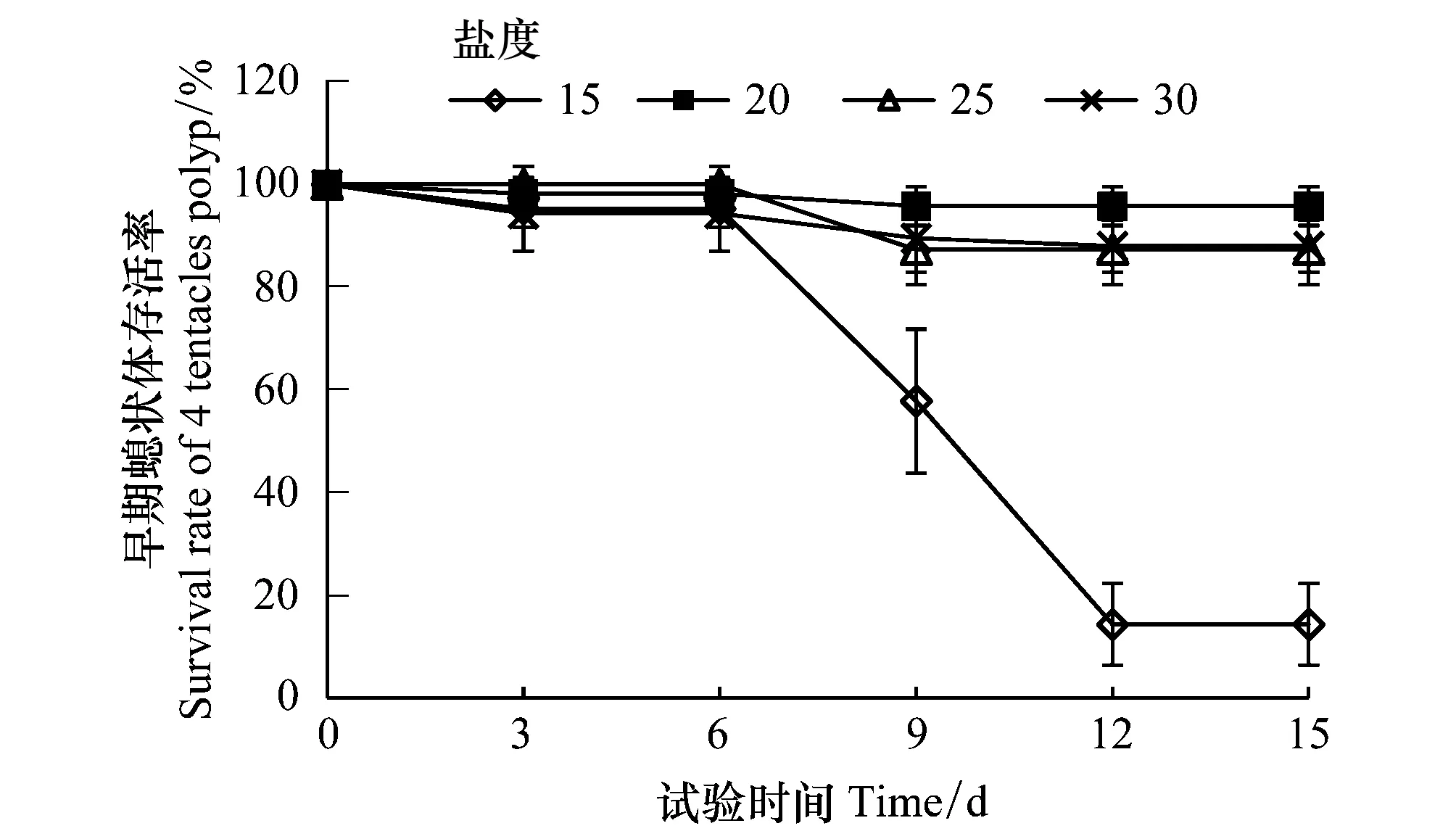
图2 不同盐度培养下的沙蜇早期螅状体存活率 Fig.2 Survival rates of N. nomurai 4 tentacles polyp at different salinities
试验结果表明,盐度在20—30间适宜沙蜇早期螅状体存活,低盐度可导致沙蜇早期螅状体萎缩解体存活显著降低。
2.3.2盐度对早期螅状体生长的影响
试验观察发现,不健康的早期螅状体在触手收缩后捕食能力丧失,随后会出现口柄萎缩,解体现象,且无恢复情况,所以在试验期间出现触手收缩口柄萎缩则不再测量其生长。试验开始时早期螅状体口柄直径平均为0.1 mm,试验结束时的螅状体口柄直径可达0.5 mm。各试验组早期螅状体的相对增长率和特定增长率可直观反映早期螅状体的生长情况。如图3可见,各盐度组早期螅状体的相对增长率随着培养时间增长而升高,在培养第9d后相对增长率出现差异,到试验结束时,盐度25、30、20组的相对增长率显著高于盐度15组(P< 0.05),但三组间差异不显著(P> 0.05)。各盐度组早期螅状体的特定增长率趋势并不相同,其中盐度15、20组的特定增长率在培养期间持续升高,盐度25、30组在培养12 d后特定增长率出现下降(图4),在试验结束时,盐度25、30、20组的相对增长率显著高于盐度15组(P< 0.05),但三组间差异不显著(P> 0.05)。
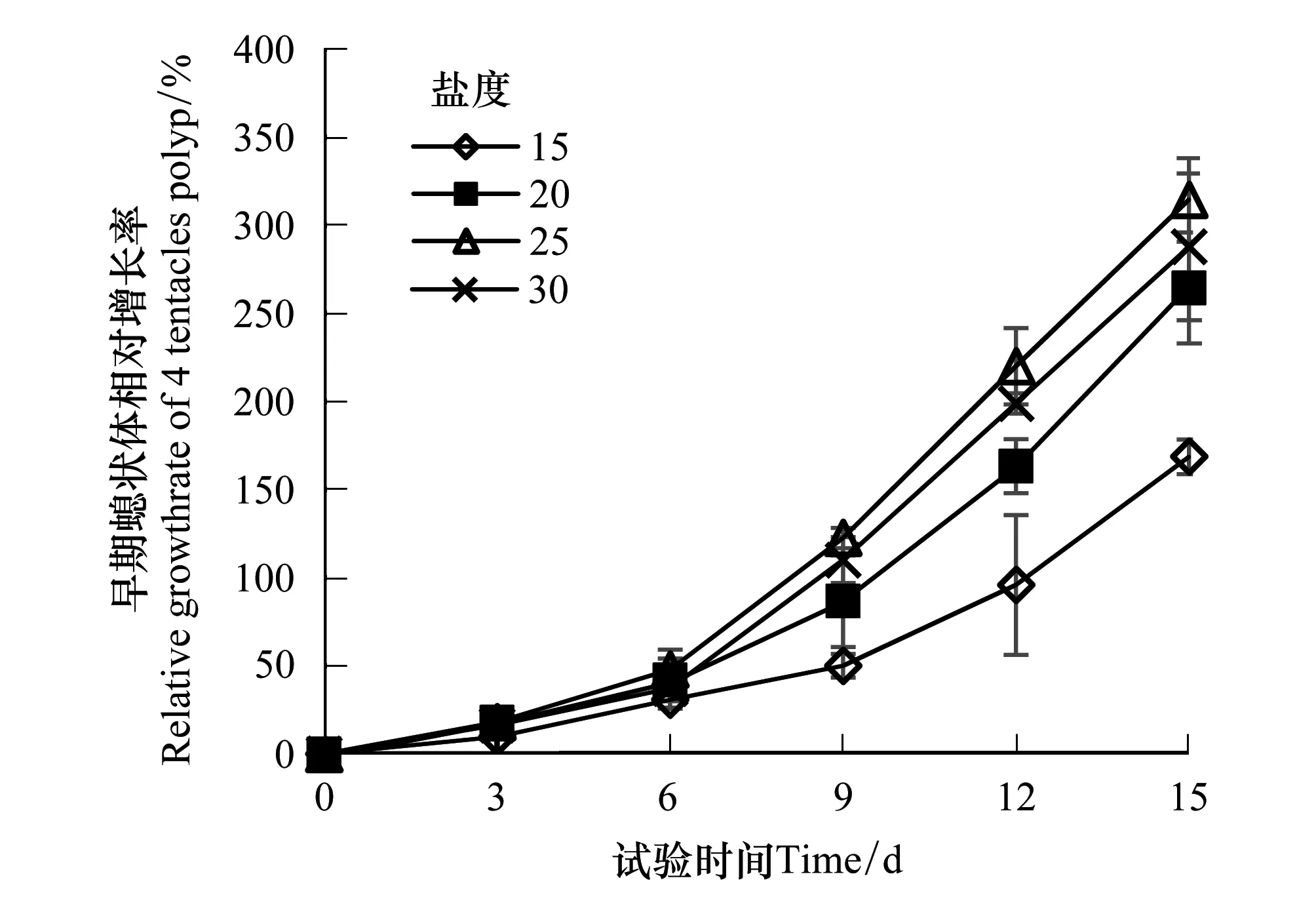
图3 不同盐度培养下的沙蜇早期螅状体相对增长率 Fig.3 Relative growth rate of N. nomurai 4 tentacles polyp at different salinities
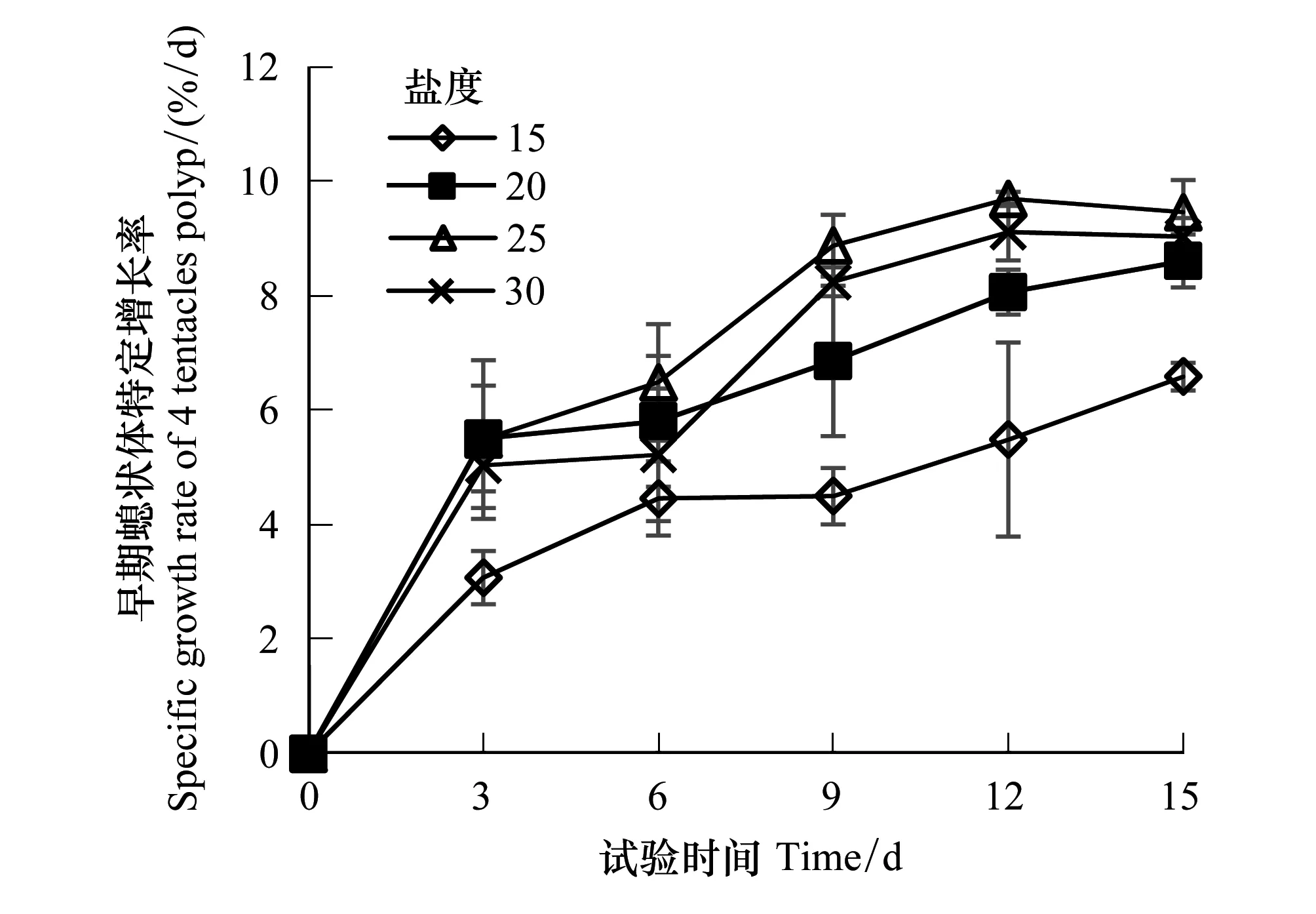
图4 不同盐度培养下的沙蜇早期螅状体特定增长率 Fig.4 Specific growth rate of N. nomurai 4 tentacles polyp at different salinities
试验结果表明,盐度在20—30间适宜沙蜇早期螅状体的生长,低盐度可导致沙蜇早期螅状体触手萎缩影响捕食,从而对螅状体生长产生不利影响。
3 讨论
水母暴发给海洋生态带来巨大挑战,同时水母暴发是全球变化下海洋生态系统演变的一种综合体现,与环境变化息息相关[29-38]。其中,盐度被认为是水母暴发的重要环境影响因子之一,对水母生活史的各发育阶段产生影响[39-46]。Dong 等[20]通过对辽东湾沙蜇连续10年的海区调查发现盐度对沙蜇生物量影响显著,在盐度较低(6—7月平均盐度25—26)的年份,沙蜇生物量远低于其他年份。同时,有研究报道沙蜇螅状体适宜的生存盐度范围较广为15—37.5[39]。那么盐度对沙蜇的影响可能发生在早期发育阶段。本文研究对象沙蜇捕获于辽东湾北部河口区,是沙蜇发源地之一[19-20],在沙蜇产卵的8—9月份,海区盐度受河流径流影响,每年的8月为主汛期,9月河口低盐区(盐度< 30)面积最大[47]。本研究结果表明。在盐度15—30范围盐度对沙蜇有性繁殖的早期阶段影响显著。
3.1 盐度对沙蜇受精卵及浮浪幼虫发育的影响
受精卵是沙蜇有性繁殖阶段早期发育的开始,受精卵发育至浮浪幼虫分为卵裂、胚层发育和器官发生3个阶段,发育时间非常短暂,本试验观察8h后盐度20、25、30组受精卵发育为浮浪幼虫,表现为体表布满纤毛在水体里游动,盐度15受精卵发育至原肠胚后期细胞体表已经出现纤毛但是尚未开始自转, Yuan[48]和陈昭廷等[49]对海月水母胚胎发育研究发现其受精卵从原肠胚期发育到浮浪幼虫是一个渐进的过程,纤毛开始于原肠胚期,浮浪幼虫出现开始于细胞单向转动,这于本试验观察相同。目前环境因子对受精卵发育影响尚未见公开报道,本文研究发现早在受精卵发育时期盐度已经开始产生影响,表现为低盐度(15)对发育的影响大于高盐度(30)。高盐度不影响受精卵发育,但对发育率产生不利影响。低盐度受精卵细胞发育延迟而且发育率明显降低,受精卵发育最适盐度为20。受精卵发育时期是形态变化和结构形成的最强烈时期,其发育率的下降受到盐度影响的同事也与自身发育有关。试验期间发现受精卵发育速度快,为了试验开始时间一致且便于观察,各试验组选取受精卵数量为10,试验在解剖镜下补充光源条件下观察及采集图片,样本数量及光照可能对试验结果产生影响。
沙蜇浮浪幼虫体型从圆形逐渐变为椭圆,在水中以纤毛活泼的游动。盐度对浮浪幼虫影响表现在活力、发育、个体消亡及存活几个方面。本文研究结果表明,盐度对沙蜇浮浪幼虫的附着变态发育影响显著。低盐度(15)导致浮浪幼虫活力降低、发育迟缓、附着发育率显著降低。这与其他种类水母的研究结果相同,Holst等[25]对德国北部海域的两种霞水母Cyaneacapillata和C.lamarckii进行了研究可见盐度对有性繁殖阶段的影响报道,指出在盐度低于20时浮浪幼虫无法附着变态。Conley等[26]研究结果表明,低盐对日本海域的海月水母(Aureliaaurita) 的浮浪幼虫附着变态产生显著影响低盐度20、15浮浪幼虫发育迟缓。孙婷婷等[24]研究发现黄渤海沿岸海月水母浮浪幼虫在低盐度环境下(18、22、26) 运动速度显著降低。本文观察发现盐度15时浮浪幼虫存活时间较长,但是附着时间集中在培养后的第3—4天,之后的浮浪幼虫虽存活多天但没有继续附着变态发育。Conley等[27]对海月水母研究中发现,盐度15时海月水母浮浪幼虫的存活时间显著增加,但附着变态后的螅状体发生了畸形,与本文观察到的结果相似。所以,本文认为低盐(15)时浮浪幼虫的存活时间增长对其附着发育没有帮助,是发育延迟的表现。本文还发现相比于受精卵发育,沙蜇的浮浪幼虫对高盐度的适宜能力有所增加,盐度20、25浮浪幼虫发育附着发育率差异不显著,适宜盐度下先附着的浮浪幼虫发育形成的早期螅状体已经具备捕食能力,且能捕食到水中游动的浮浪幼虫,从而获取到营养进行下一步的生长发育。试验期间盐度20、25组的浮浪幼虫附着变态成的早期幼体对试验组的浮浪幼虫进行了捕食,可能对两组的浮浪幼虫附着变态率产生影响,但不影响结果趋势。
3.2 盐度对沙蜇早期螅状幼体存活及生长的影响
沙蜇的早期螅状体指浮浪幼虫附着后发育成的4触手螅状体,口柄直径平均为100 μm。成熟螅状体具有16触手,口柄直径700—1300 μm。本文研究发现,盐度20、25、30是沙蜇的早期螅状体生长存活的适宜盐度,三组盐度存活率在87.79%—95.77%之间没有显著差异。盐度15时早期螅状幼体的存活显著降低,个体生长较慢。Dong 等[39]研究发现沙蜇的成熟螅状体在盐度10—30的环境下可存活且较好的成长。与成熟螅状体相比,早期螅状体对盐度的适应范围要窄一些,分析原因可能为触手少口柄直径小,捕食能力弱,体内增值细胞更处于分化中,对环境的适应性差,随着生长发育其触手增多、体积增大,对环境的适应能力也增强。研究结果表明,相对于受精卵和浮浪幼虫的发育早期螅状体对高盐度的适应能力进一步的增加,但盐度对早期螅状体的生长存活有显著影响。其中,低盐(15)导致沙蜇早期螅状幼体生长缓慢且存活率显著下降。在盐度对海月水母螅状体生长存活的研究中也发现低盐的不适应性,其中Sokolowski等[27]指出波罗的海的海月水母在盐度12以下不能正常生活。孙明等[45]发现盐度低于15后黄渤海海域海月水母螅状体无法存活。
综合试验结果,本文认为盐度对沙蜇有性繁殖阶段早期发育影响显著。随着沙蜇受精卵、浮浪幼虫、早期螅状的生长发育,对盐度的适应能力逐步增加。这种趋势与沙蜇的生存环境相同。沙蜇的有性繁殖发生在8—9月,降雨期使河流的径流量增大河口沿岸的低盐(30)范围扩大,给河口近海的低盐环境给沙蜇的有性繁殖提供了适宜的盐度,有性繁殖结束新的底栖螅状体生成,河流开始进入枯水期,海区盐度逐步升高,螅状体在海区越冬至第二年春季横裂或继续以螅状体形态多年存活。但是,夏季季风季节的极端降雨事件导致径流量激增,使河口近海盐度短期内显著降低,本文研究发现沙蜇受精卵发育仅需8 h,浮浪幼虫附着集中在3—4 d,那么海区短期的盐度降低可能成为沙蜇有性繁殖的不利因素。Dong等[20]通过调查发现盐度对沙蜇生物量影响显著,在6、7月平均盐度25—26年份,沙蜇生物量远低于盐度30—32年份。在沙蜇生活的海域6、7月进入雨季,8月为主汛期,9月海区盐度达到最低值。这可能是海区沙蜇减产的原因。在更早的鲁男等[50]关于辽东湾海蜇资源量锐减的原因的研究中,也指出80到90年代辽东湾海蜇的三次资源量锐减(低于上一年95%以上)与前一年双台子河和大辽河的8月平均淡水注入量过多(超100 m3/s)吻合。沙蜇分布海区生态环境复杂多变,影响其暴发的原因也是多方面的,盐度作为重要环境之一,通过对沙蜇有性繁殖阶段早期发育的影响进而影响了沙蜇对其螅状体种群的数量的补充,从而影响其水母体的资源量及暴发是值得探讨的。
