孕穗期冷水灌溉对寒地粳稻相关生理指标及叶绿体超微结构的影响
2020-08-17张盛楠瞿炤珺赵宏伟
张盛楠,贾 琰,瞿炤珺,杨 亮,李 晓,张 妍,王 喆,赵宏伟
(1.寒地粮食作物种质创新与生理生态教育部重点实验室,东北农业大学,哈尔滨 150030;2.东北农业大学 农学院,哈尔滨 150030)
水稻作为中国主要的粮食作物,对保证国内粮食安全发挥着重要作用[1]。黑龙江省是主要的粳稻产区,但其位于中国最北端,年平均气温较低,导致低温冷害现象频繁发生,对黑龙江省粮食生产造成严重影响。因此,低温冷害已经成为影响寒地粳稻高产稳产的气象灾害之一。
水稻是喜温作物,在孕穗期对温度十分敏感[2]。水稻在孕穗期遭受低温冷害胁迫会影响正常生理代谢过程,表现出结实率低,产量下降[3]。Kuk等[4]研究表明,冷胁迫显著增加水稻苗期功能叶片抗氧化酶的活性,其中CAT活性增加最为显著。朱珊等[5]研究发现,在孕穗期低温胁迫下水稻内源激素发生显著性变化。Taylor等[6]研究表明,低温胁迫会对叶绿体超微结构造成严重影响。目前大量研究集中于低温胁迫对水稻苗期生长发育及生理代谢过程等方面[7-8],而对水稻孕穗期相关生理指标、叶绿体超微结构及产量的研究还相对较少。因此本试验采取冷水灌溉方式,大田模拟自然条件下低温冷害,以粳稻品种‘东农428’和‘松粳10’为试验材料,研究孕穗期冷水灌溉对寒地粳稻耐冷相关指标、叶绿体超微结构及产量的影响,为解决寒地粳稻孕穗期低温冷害问题提供参考。
1 材料与方法
1.1 试验材料
选用经东北农业大学水稻研究所前期筛选出的生育期相近的耐冷型品种‘东农428’(‘DN428’)和冷敏感型品种‘松粳10’(‘SJ10’)为试验 材料[9]。
1.2 试验设计
试验于2018年在东北农业大学阿城实验基地(东经127.04°,北纬45.52°)进行。4月13日播种,5月20日移栽。采用裂区试验设计,以不同时间冷水灌溉处理为主区,在孕穗期(幼穗长约1 cm)进行0(正常灌溉)、5、10和15 d的冷水灌溉处理,分别记为CK、D5、D10、D15。以粳稻品种为副区,设置3次重复,每个副区面积为18 m2。插秧规格为30 cm×10 cm,行长6 m,每3株为1穴。在2018年7月12日进行冷水灌溉,采用自动冷水灌溉系统进行冷水灌溉处理,冷水灌溉温度为17 ℃(为黑龙江省孕穗期低温冷害温度)。灌水深度在20 cm[9]左右。冷水灌溉的时间为6:00-20:00。以正常灌溉为对照,处理结束后恢复正常灌溉,正常灌溉为晒水池中水(22~24 ℃)。其他同一般大田管理。采用田间气候自动测定仪测定大田气温、土壤温度(深度10 cm)、光照辐射量,具体信息见表1。
表1 孕穗期冷水灌溉期间田间试验条件Table 1 Field test conditionunder cold water stress at booting stage
1.3 取样方法
生理指标:选取各处理生长整齐一致的植株进行标记,孕穗期开始进行冷水灌溉处理,处理结束当天8:00-10:00取样,取水稻植株最上面的完全展开叶,3次重复,放在-80 ℃冰箱保存,备用。
叶绿体超微结构:选取各处理生长整齐一致的植株进行标记,孕穗期开始进行冷水灌溉处理,处理结束当天8:00-10:00取样,选取水稻植株最上面的完全展开叶进行超微结构观察,叶片剔除中间叶脉,切割成1 mm×3 mm的切片若干,放入戊二醛固定液中保存,在H-760型透射电子显微镜下观察拍照。
产量:于成熟期,各处理选取2 m2实割,3次重复,收获后自然风干,脱粒测定每公顷的实际 产量。
1.4 测定项目和方法
1.4.1 抗氧化系统 称取0.5 g水稻叶片,加入5 mL(pH=7.8)的磷酸缓冲液,冰浴研磨成匀浆,在10 000 r/min、4 ℃下离心10 min,取上清液用于抗氧化酶活性测定。超氧化物歧化酶(SOD)的活性采用氮蓝四唑法[10]测定,过氧化物酶(POD)的活性采用愈创木酚法测定[10]。过氧化氢酶(CAT)活性采用高锰酸钾滴定法[10]测定。
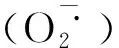
1.4.2 渗透调节物质 可溶性糖(SS)采用蒽酮-硫酸比色法[13],称取0.5 g水稻叶片,加入4 mL 80%酒精研磨成匀浆,倒入离心管中,80 ℃水浴锅中水浴30 min,在5 000 r/min离心机下离心10 min,收集上清液,其残渣加2 mL 80%酒精重复提取1次,合并上清液,上清液中加入适量活性炭,80 ℃水浴锅中脱色20 min,过滤后吸取滤液进行测定。
可溶性蛋白(SP)采用考马斯亮蓝法[14],称取0.5 g水稻叶片,加入5 mL(pH=7.8)的磷酸缓冲液,冰浴研磨,在10 000 r/min、4 ℃下离心10 min,取上清液用于可溶性蛋白含量测定。
1.4.3 内源激素 采用酶联免疫测定法[15]测定脱落酸(ABA)、吲哚乙酸(IAA)、赤霉素(GA3)、玉米素核苷(ZR)含量。
1.5 数据处理与分析
以3次重复试验的平均值为试验数据。Microsoft Excel 2013进行数据处理和作图,SPSS 22.0进行相关数据统计和方差分析,使用RStudio进行相关性分析作图。
2 结果与分析
如表2所示,所测指标处理间和品种间均差异显著。处理和品种互作间方差分析表明,CAT、GA3和产量之间差异不显著,其余指标均差异显著。
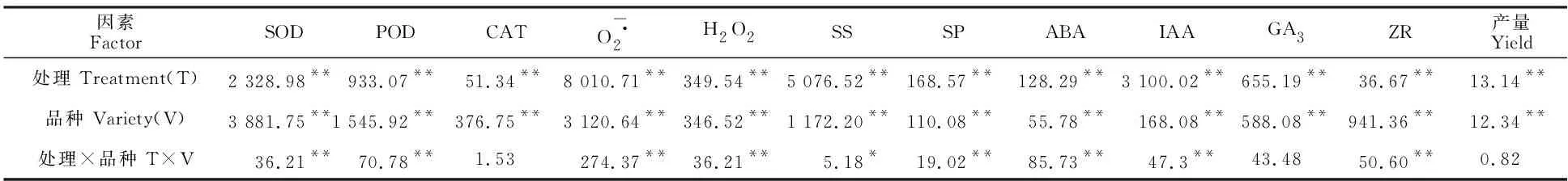
表2 处理和品种之间的方差分析Table 2 Analysis of variance between treatment and variety
2.1 孕穗期冷水灌溉对寒地粳稻抗氧化酶系统的影响
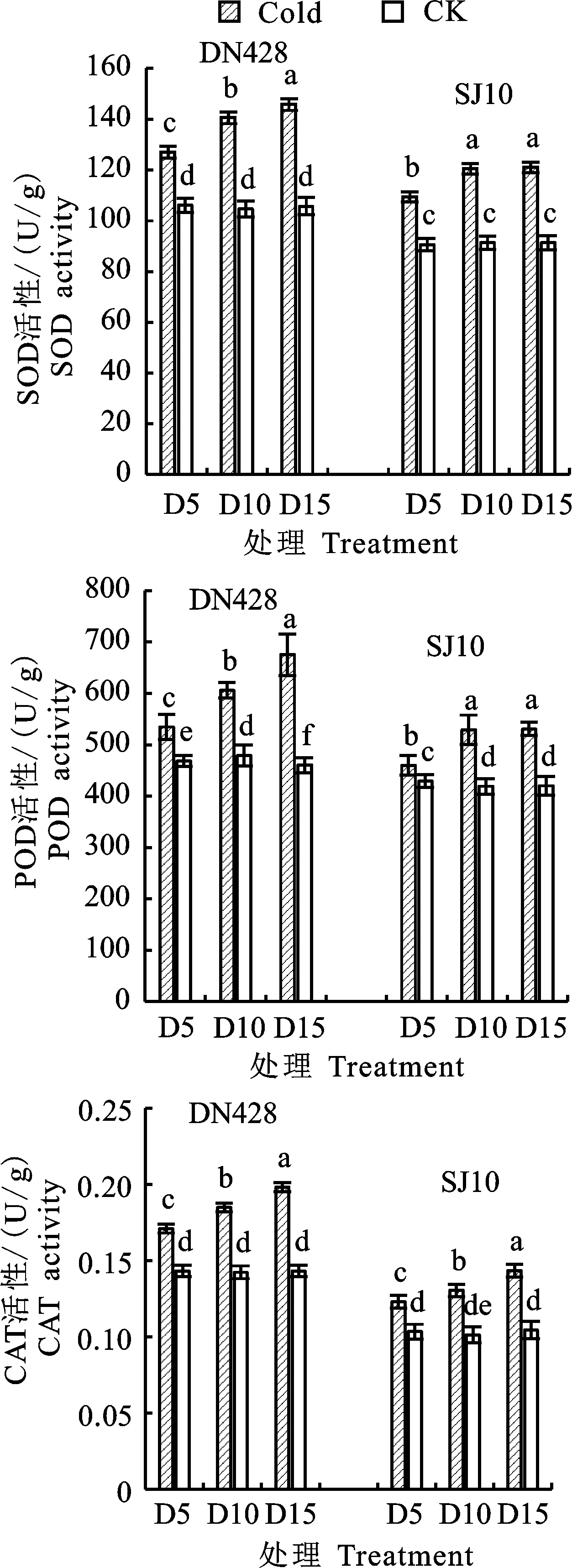
在孕穗期冷水灌溉下,水稻启动保护酶系统,通过消除体内多余的自由基来减轻冷水胁迫造成的伤害。由图1可知,与对照相比,‘DN428’和‘SJ10’在冷水灌溉下,功能叶片中SOD、POD、CAT活性显著上升,均随冷水灌溉时间增加而呈上升趋势,上升幅度也随冷水灌溉时间而增加,在D15处理下达到最大值,具体表现为D15处理下‘DN428’功能叶片SOD活性增幅为32.11%,POD活性增幅为31.81%,CAT活性增幅为 27.69%,‘SJ10’增幅分别为28.40%、20.86%、 27.15%。‘DN428’在冷水灌溉处理下,SOD、POD、CAT活性在各处理中差异显著。‘SJ10’功能叶片CAT活性在冷水灌溉各处理下差异显著,而SOD、POD活性在D10和D15处理下差异不显著。‘DN428’功能叶片在冷水灌溉处理下抗氧化酶系统增幅大于‘SJ10’,说明‘DN428’功能叶片抗氧化酶系统比‘SJ10’能更快速的响应冷水胁迫。
2.2 孕穗期冷水灌溉对寒地粳稻活性氧含量的影响
显著性分析中同一品种不同处理和对照之间一起比较,不同字母之间表示差异显著(P≤0.05)。下同。

图2 孕穗期冷水灌溉下寒地粳稻含量变化Fig.2 Changes of H2O2 contents in japonica rice under cold water irrigation at booting stagein cold region
2.3 孕穗期冷水灌溉对寒地粳稻渗透调节物质的影响
由图3可知,与对照相比,‘DN428’和‘SJ10’在冷水灌溉下,SS和SP含量显著上升。在孕穗期冷水灌溉下,‘DN428’和‘SJ10’功能叶片中SS和SP含量随着冷水灌溉时间的增加呈上升趋势,增幅也随冷水灌溉时间而增加,均在D15处理下达到最大值,具体表现为D15处理下‘DN428’功能叶片SS含量增幅为34.83%,SP含量增幅为 48.97%,‘SJ10’功能叶片SS含量增幅为44.69%,SP含量增幅为29.39%。‘DN428’功能叶片SS和SP含量在各冷水处理下差异显著,‘SJ10’功能叶片SS含量在各冷水处理下差异显著,而SP含量在D10和D15处理下无显著差异。
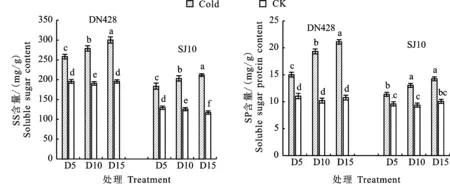
图3 孕穗期冷水灌溉下寒地粳稻SS和可SP含量变化Fig.3 Changes of soluble sugar and soluble protein content in japonica rice under cold water irrigation at booting stagein cold region
2.4 孕穗期冷水灌溉对寒地粳稻内源激素含量的影响
由图4可知,‘DN428’和‘SJ10’功能叶片中,内源激素ABA在各冷水处理下与对照相比显著上升,经过冷水灌溉后,‘SJ10’功能叶片中ABA含量随冷水灌溉时间的增加而显著上升,在D15处理下达到最大值,在D15处理下增幅为 30.96%。而‘DN428’功能叶片ABA含量在D5处理下最大,D10和D15处理差异不显著,在D5处理下增幅为24.87%。从图4中IAA、GA3、ZR含量表明,除‘DN428’的D5处理与对照差异不显著外,‘DN428’和‘SJ10’各处理中功能叶片IAA、GA3、ZR含量均显著小于对照,且随着冷水灌溉时间的增加呈下降趋势,具体表现为‘DN428’功能叶片IAA、GA3、ZR含量在D15处理下降幅为10.98%、25.64%、35.09%,‘SJ10’降幅分别为20.87%、36.49%、46.39%。两品种中功能叶片IAA含量在D10和D15处理下无显著差异。‘DN428’功能叶片中GA3和ZR含量在D10和D15处理下差异不显著,而‘SJ10’各冷水处理下差异显著。
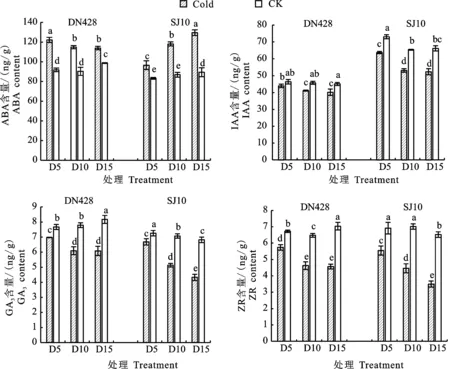
图4 孕穗期冷水灌溉下寒地粳稻内源激素含量变化Fig.4 Changes of endogenous hormones in japonica rice under cold water irrigation at booting stagein cold region
2.5 孕穗期冷水灌溉对寒地粳稻叶绿体超微结构的影响
如图5-A、5-E,在CK下,‘DN428’和‘SJ10’叶绿体结构完好,呈椭圆形或者纺锤形紧贴在细胞壁上,类囊体片层均很清晰,‘SJ10’叶绿体内部有少量的淀粉粒存在。研究表明,叶绿体在遭受低温冷害胁迫后,会表现出结构混乱,膨大,内部类囊体片层结构解体或消失[16]。在D5处理下(图5-B、5-F),‘DN428’和‘SJ10’叶绿体均出现少量的嗜锇粒,两品种中叶绿体结构与对照并未出现明显差异,‘DN428’叶绿体内部出现少量淀粉粒。在D10处理下(图5-C、5-G),‘DN428’和‘SJ10’叶绿体和类囊体肿胀或变细长,出现大量的嗜锇粒,且‘SJ10’出现质壁分离现象。在D15处理下(图5-D、5-H),叶绿体结构不规则且与细胞壁之间存在明显空隙,可观察到叶绿体和类囊体肿胀,叶绿体类囊体片层模糊,‘SJ10’叶绿体内部呈现空泡化,嗜锇粒明显增多。
2.6 孕穗期冷水灌溉对寒地粳稻产量的影响
由表3可知,与对照相比,孕穗期冷水灌溉下寒地粳稻的产量均显著下降,各处理之间差异显著,且随着冷水灌溉时间的增加下降幅度逐渐增大。‘DN428’在D5、D10、D15处理下产量较CK分别下降15.81%、25.64%、37.87%。‘SJ10’在D5、D10、D15处理下产量较CK分别下降 19.27%、33.58%、47.94%。‘DN428’各冷水处理下产量CRI为62.13%~84.19%,‘SJ10’各冷水处理下产量CRI为52.06%~80.73%。由此可见孕穗期冷水灌溉处理对‘SJ10’产量影响比‘DN428’大。
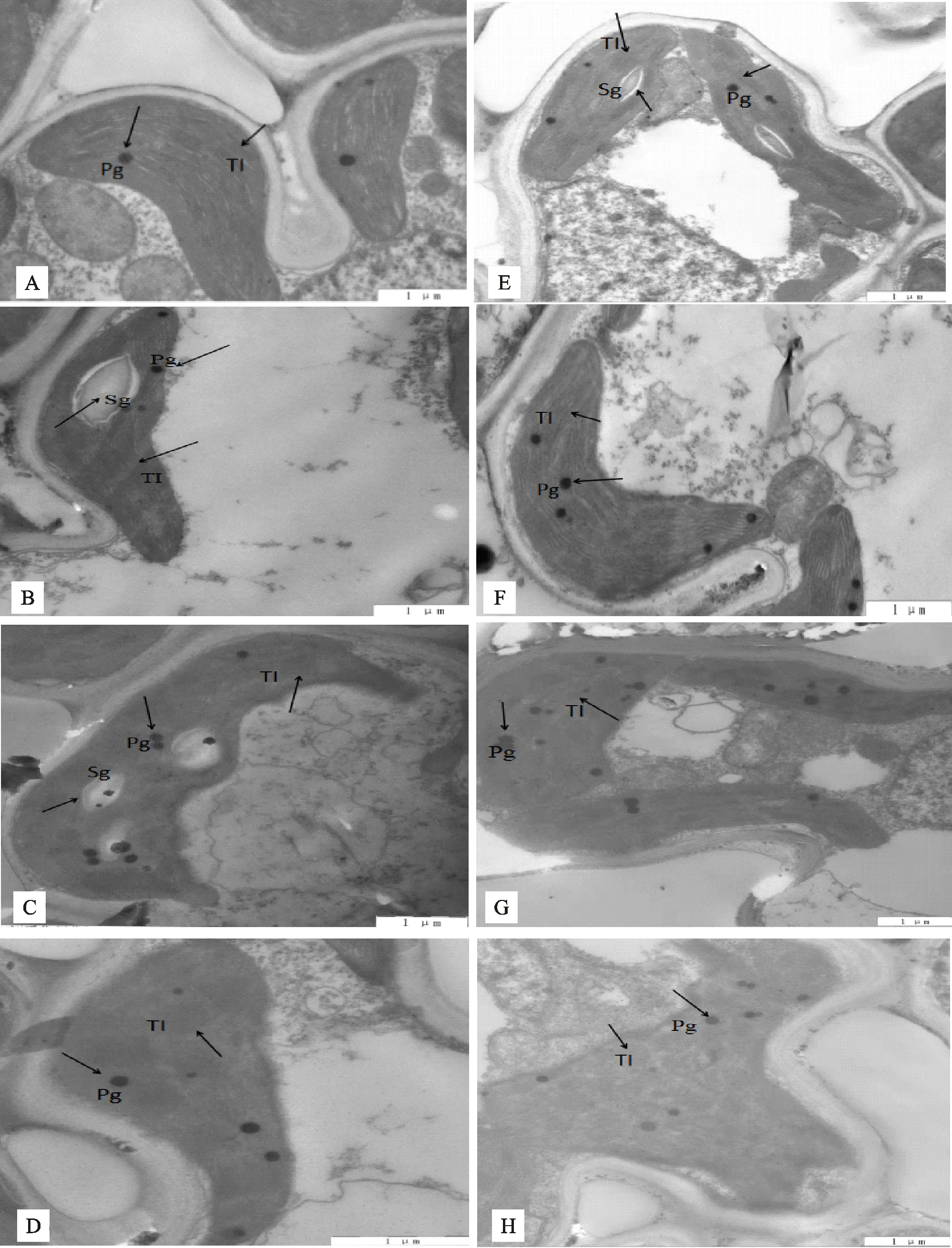
A.‘DN428’-CK;B.‘DN428’-D5;C.‘DN428’-D10;D.‘DN428’-D15;E.‘SJ10’-CK;F.‘SJ10’-D5;G.‘SJ10’-D10;H.‘SJ10’-D15
2.7 孕穗期冷水灌溉下相关生理指标之间的相关分析
由图6可知,内源激素ABA与抗氧化酶、活性氧物质和渗透调节物质呈极显著正相关关系,与其他3种内源激素呈极显著负相关关系。IAA与抗氧化酶、渗透调节物质呈极显著负相关关系。GA3与活性氧物质呈极显著负相关关系,ZR与活性氧物质、SOD、POD、渗透调节物质呈极显著负相关关系或显著负相关关系。从整体上来看,ABA是与耐冷相关生理指标相关性最大的内源激素。
2.8 孕穗期冷水灌溉下寒地粳稻相关生理指标与产量的回归分析
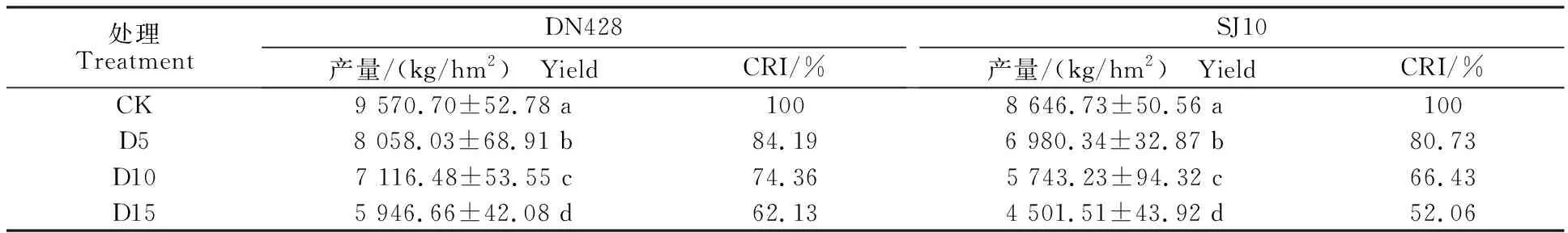
表3 孕穗期冷水灌溉后寒地粳稻的产量Table 3 Yield of japonica rice in cold region under treatment of cold water irrigation at booting stage

x,y轴代表相关生理指标,颜色代表相关系数(红色代表正相关,蓝色代表负相关,颜色越深代表相关性越高),* 表示在 0.05水平上显著相关。**表示在0.01水平上显著相关

表4 孕穗期冷水灌溉下寒地粳稻相关生理指标CRI与产量CRI的关系Table 4 Relationship between CRI and yield CRI of related physiological indexes of japonica rice in cold regionunder cold water irrigation at booting stage
3 讨 论
内源激素是一种有机化合物,在逆境胁迫下能调节植物自身生理过程[27]。Davies[28]认为,植物的内源激素具有复杂的生理效应,既有协同作用,又有拮抗作用。邵怡若等[29]研究发现当植物受到低温胁迫的情况下,ABA含量升高。这与本研究结果一致,可能原因是在冷水灌溉下,寒地粳稻功能叶片细胞膜受到破坏,导致功能叶片电子渗透率增加,ABA含量的增加可以降低水稻植株生长势,对低温胁迫做出相应的适应性反应[30]。在孕穗期冷水灌溉处理下,IAA、GA3和ZR含量下降,原因可能为促进型植物激素含量下降,可以延缓植物生长速度,维持体内代谢平衡。本试验中发现,冷敏感型水稻‘SJ10’内源激素ABA在冷水灌溉下上升幅度大于耐冷型水稻‘DN428’,而冷敏感型水稻‘SJ10’内源激素IAA、GA3和ZR含量比耐冷型水稻‘DN428’下降幅度要大,分析认为冷敏感型水稻在受到冷水胁迫后具有较强合成ABA的能力,促进植物进入休眠状态。同时促进型植物激素(IAA、GA3和ZR)含量下降,降低水稻细胞伸展性,减少因冷水灌溉带来的伤害。从抗氧化系统与内源激素的相关分析可知,内源激素ABA对抗氧化系统和渗透调节物质相关性最大。徐新娟[31]指出ABA在低温胁迫下会使植物产生较多的H2O2,促进植物气孔关闭,减少蒸腾作用,进而改变植株体内各种生理生化效应。本研究表明ABA可以促进植物抗氧化酶活性的增加,来抵御逆境胁迫。从可溶性糖和可溶性蛋白与内源激素相关分析可知,在冷水灌溉下ABA含量的增加可以提高SS和SP含量,用于维持植物细胞渗透压平衡[32]。
叶绿体是主要的光合细胞器,它含有丰富的类囊体膜。在正常灌溉下,叶绿体呈椭圆形或者纺锤型紧贴在细胞壁上,类囊体片层清晰可见[33]。在冷水灌溉下,叶绿体结构不规则且与细胞壁之间存在明显空隙,叶绿体和类囊体肿胀,类囊体片层模糊。从‘DN428’和‘SJ10’叶绿体超微结构对比发现,冷水灌溉下,‘DN428’叶绿体超微结构损伤较轻,有少量淀粉粒出现。分析认为,活性氧物质主要的产出地是叶绿体,叶绿体超微结构受损程度取决于活性氧物质含量及抗氧化酶活性高低[34]。同时‘DN428’淀粉粒的出现是对冷水胁迫做出的适应反应。由于‘DN428’在冷水灌溉下比‘SJ10’具有高的抗氧化酶活性和低的活性氧水平,因此叶绿体超微结构损伤程度较轻。
4 结 论
