miR-374b及其靶基因Myf6 调控成肌细胞增殖分化的研究
2020-08-16韩福慧栾兆进王国义贺建宁
韩福慧 ,李 倩,栾兆进,王国义,柳 楠,贺建宁,薛 明
(1.青岛农业大学动物科技学院,山东青岛 266109;2.包头市农业科学研究所,内蒙古包头 014013;3.赤峰市敖汉种羊场,内蒙古赤峰 024300;4.全国畜牧总站,北京 100125)
在骨骼肌的生长发育过程中,骨骼肌细胞需经历增殖、分化、再生等过程,除已知生肌调节因子家族(MRFs)和肌细胞增强因子家族(MEF2)起调控作用外[1],越来越多的miRNAs被发现在细胞增殖分化过程中扮演重要角色。miRNA作为重要的转录后调节因子,通过与靶基因mRNA的非编码区的种子序列碱基互补配对结合,诱导靶mRNA降解或阻遏其翻译[2]。miR-206可通过抑制ld(MyoD抑制剂)的表达促进肌细胞生成素(MyoGenin,MyoD)的表达,从而促进成肌细胞分化[3-4]。miR-24在分化的成肌细胞中高表达,TGF-β/Smad3可以通过抑制miR-24的表达来抑制成肌细胞的分化[5]。miR-374b主要在胃癌、肺癌和乳腺癌等癌症发生的过程中促进细胞增殖[6]。马志远[7]研究显示miR-374b负向调控小鼠成肌细胞的分化,促进细胞增殖。Ma等[8]研究表明,miR-374b直接靶向Myf6基因并负调节肌细胞生成。Myf6基因作为MRFs家族的成员,在肌肉细胞分化过程中起重要作用[9],并通过与卫星细胞和肌纤维的融合在肥大肌肉生长中发挥作用[10]。
目前对miR-374b的研究多集中在对癌细胞和小鼠成肌细胞增殖分化的影响,但是否对绵羊等家畜成肌细胞增殖分化有影响鲜有报道。因此,本研究通过过表达和抑制miR-374b后Myf6基因的变化及其对绵羊成肌细胞增殖分化的影响,验证在绵羊肌细胞中miR-374b与Myf6基因的关系,为肉羊分子育种提供一定理论参考。
1 材料与方法
1.1 实验动物及样品采集 选择健康70日龄的南非肉用美利奴羊1只,将其从青岛奥特种羊场连同子宫一起带回无菌操作实验室。
1.2 成肌细胞分离培养 将带回细胞间的绵羊子宫剪开,取出胎羊,迅速剪下胎羊后腿用PBS冲洗数次,移入无菌超净台中,剔除皮肤、筋膜等组织后,采用I型胶原酶和胰酶两步酶消化法分离细胞,结合差速贴壁纯化得到成肌细胞,置于培养箱培养。
1.3 诱导成肌细胞分化 细胞传代后,当细胞密度达到70%左右时,弃去完全培养基,用PBS(含双抗)清洗,加入配好的分化培养基,诱导成肌细胞分化,每天换液1次;观察记录分化第0(未加分化培养液时)、24、72、120小时细胞形态变化,并依次收集细胞,放-80℃冰箱暂存,用于总RNA提取。
1.4 不同分化时期实时荧光定量PCR采用Trizol提取不同分化时期细胞总RNA,采用核酸蛋白分析仪(Eppendorf)测量RNA浓度,Myf6、MyHC、MyoG、MyoD基因按照罗氏反转录试剂盒进行操作,取cDNA进行qRT-PCR,GAPDH基因为内参。反应程序:95℃预变性3 min,95℃变性10 s,60℃退火20 s,72℃延伸20 s,40个循环。miR-374b按照microRNA逆转录试剂盒使用说明书进行,以5sRNA为内参,反应程序:95℃预变性3 min;PCR反应95℃ 12 s,62℃ 40 s,共40个循环,62℃时采集荧光信息。采用2-ΔΔCt计算进行单因素方差分析,引物序列见表1。
1.5 细胞转染 挑选生长状态良好的细胞,细胞密度达到85%~90%时,弃去完全培养基,用PBS(无双抗)清洗,胰酶消化后加完全培养基终止消化,离心后弃去上清,清洗2次,加入纯培养基,轻轻吹打混匀,将细胞悬液加到六孔板中,再加入完全培养基至每孔最终液体为2 mL;待六孔板中的细胞生长贴壁至30%~40%时,用PBS(无双抗)清洗2遍,换纯培养基;按照micoRNA mimics、inhibitor和阴性对照说明书,对六孔板中的细胞进行转染;转染后12 h除去纯培养基,然后每孔加入2 mL分化培养基,48 h后收集细胞,放入-80℃冰箱保存,用于总RNA和蛋白提取。
1.6 转染后细胞实时荧光定量PCR提取RNA、反转录以及荧光定量PCR步骤同1.4。
1.7 Western blot检测 将收集的细胞从-80℃冰箱取出,低速离心后吸出多余残留液体,测定蛋白浓度,按蛋白浓度倍数添加裂解液稀释蛋白使各蛋白浓度在同一水平,加入裂解液后轻轻吹打混匀,静置裂解30 min;期间依次配置分离胶和浓缩胶,将Marker和所需样品缓慢加入泳道;蛋白经凝胶电泳分离后转膜于PVDF膜,加封闭液室温摇床封闭2.5~3 h;加一抗(1:1 000),4℃,过夜;PVDF膜1×TBST摇床清洗3次后加二抗(1:2 000),室温孵育1 h,注意避光;PVDF膜1×TBST摇床清洗3次,超纯水摇床清洗2次;PVDF膜上加显色液(A液:B液=1:1),覆上保鲜膜,凝胶成像仪上曝光,Image J 1.39U进行目的条带的光密度值分析。
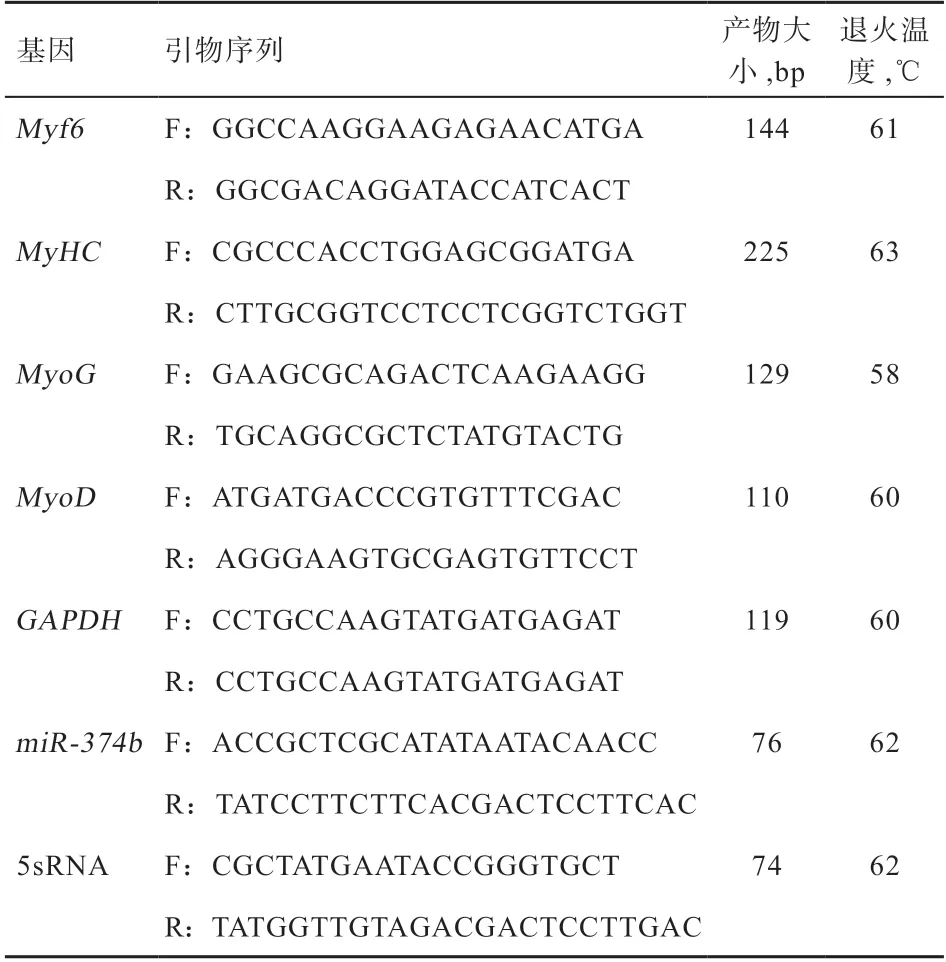
表1 引物序列
2 结果
2.1 成肌细胞的体外分化 如图1所示,细胞分化第24小时细胞形态无明显变化,诱导分化第72、120小时出现大量肌管。
2.2 不同分化时期基因表达量 如图2所示,随着细胞分化时间的增长,Myf6基因表达量也随之增长,在分化第0小时与第24小时差异不显著,第72、120小时与第0、24小时差异极显著。而miR-374b基因的表达趋势为先上升后下降,在分化第0、24、72小时呈递增趋势(P<0.01),分化第120小时miR-374b基因的表达量出现极显著下降,但仍高于生长期表达量。
由图3可知,增殖标志基因MyoD在分化24 h表达量最高,极显著高于其他3个时期;120 hMyoD表达量次之,与0 h差异不显著,显著高于72 h;分化标志基因MyHC、MyoG在细胞增殖期及分化初期表达量最低,在分化72 h和120 h时表达量呈现极显著上升。以上结果说明,绵羊成肌细胞分化成功。
2.3miR-374b调控Myf6及各标志基因的表达 如图4所示,miR-374b过表达载体(mimics)转染成肌细胞后,miR-374bmRNA表达量极显著高于阴性对照组,而Myf6基因mRNA表达量极显著低于阴性对照组,Myf6基因蛋白表达量极显著低于阴性对照组。结果表明,成肌细胞过表达miR-374b后显著抑制了Myf6基因的表达。
由图5可知,miR-374b过表达后MyHC基因mRNA表达量极显著低于阴性对照组,MyoG、MyoD基因表达量与阴性对照组差异不显著。
如图6所示,miR-374b抑制载体(inhibitor)转染成肌细胞后,miR-374b表达量极显著低于阴性对照组,而Myf6基因表达量极显著高于阴性对照组;Myf6基因蛋白表达量极显著高于阴性对照组。结果表明,成肌细胞抑制miR-374b后使得Myf6基因的表达显著上升。
由图7可知,转染miR-374b抑制载体后,MyHC、MyoD基因表达量极显著高于阴性对照组,MyoG基因表达量显著低于阴性对照组。
3 讨论
细胞在分化过程中最先被激活形成成肌细胞,成肌细胞继续增殖并沿着一定方向生长,它们平行排列并融合形成肌管,随着分化程度的加深,肌丝生长成为肌原纤维,细胞核移至周围后形成肌纤维[11]。本实验采用酶消化法分离细胞,相较于组织块分离法获得细胞更加迅速,极大地缩短了实验用时,同时采用差速贴壁纯化成肌细胞,2 h后换液,可有效排除其他贴壁细胞的干扰,保证成肌细胞的均一。在诱导成肌细胞分化时,分化第72小时细胞已明显分化融合形成肌管,在小鼠中分化第72小时可以观察到少量肌管的出现,到第96小时有大量肌管,这可能与物种差异有关[7]。
生肌调节因子(MRFs)家族及肌细胞增强因子(MEF2)家族分别在骨骼肌发育中起到至关重要的作用[12]。MRFs家族由MyoD、MyoG、Myf5和Myf64个基因组成,它们在不同的染色体位置上具有不同的表达模式[13]。Myf5和MyoD是初级的肌肉调节因子,其决定是否可以激活静息肌肉卫星细胞以成为有着分化功能的肌肉干细胞。因此,它们参与肌肉细胞的定型,并且一般在成肌细胞增殖和分化中表达[14]。MyoG和Myf6基因为次级肌源性调节因子,其在分化和融合过程中表达,主要功能是诱导成肌细胞的分化和融合成肌管[15]。因此,MyoG基因可用作细胞分化的标记基因[16]。因此,本实验通过对MyoG、MyoD、Myf6等标志基因的表达量进行分析,从而揭示miR-374b对成肌细胞增殖分化调控作用。
本研究结果表明,miR-374b表达量随着分化时间的增加呈先增长后下降趋势,这预示着miR-374b可能在成肌细胞分化的前期和后期发挥了不同的生物学功能,有待于进一步研究;同时其变化趋势与在小鼠成肌细胞分化研究中miR-374b表达量结果一致[7];Myf6及分化标志基因MyHC、MyoG表达呈持续增长趋势,MyoG基因在细胞分化初期表达量较低,在细胞分化末期大量表达,符合该基因表达模式[7]。
在本研究中,miR-374b过表达组(抑制组)相对于阴性对照组极显著上调(下调),说明转染实验成功。过表达miR-374b后,Myf6基因表达量极显著下调;抑制miR-374b后,Myf6基因出现极显著上调的现象。Myf6基因在肌肉发育中结合在肌肉特异基因的启动子上,开启这些基因的表达从而调节肌肉分化[9]。因此本实验说明在绵羊中miR-374b抑制成肌细胞分化是通过直接靶定在Myf6基因的3'UTR,参与Myf6基因的转录后调节实现的,这与在小鼠中研究结果相同[8]。同时本研究中抑制miR-374b的表达后MyoD基因表达量极显著高于阴性对照组,MyoG基因表达量显著低于阴性对照组,可能是因为MyoG基因的干扰伴随有Myf6基因的高表达,Myf6基因对MyoG有部分代偿作用,而Myf6基因的干扰并未伴随有MyoG基因的高表达,Myf6与MyoG基因的代偿作用是单向的而非相互的,这与文献中结果一致[17]。
4 结论
miR-374b表达量随着分化时间的增加呈先增长后下降趋势,可能在细胞分化前后发挥着不同的生物学功能;Myf6及各分化标志基因随着细胞分化时间增长,mRNA表达量随之增长,绵羊成肌细胞分化成功。对成肌细胞转染miR-374b的过表达及抑制载体的结果说明miR-374b负向调控MyHC、Myf6基因的表达,进而抑制成肌细胞分化。Myf6可能对MyoG基因有部分代偿作用。综上结果表明,在绵羊成肌细胞中miR-374b也可通过靶定Myf6基因,进而负向调控成肌细胞分化,抑制绵羊骨骼肌生长。
