新疆野生苹果林苹小吉丁的伴生真菌多样性*
2020-08-14周忠福赵文霞林若竹淮稳霞姚艳霞
周忠福 赵文霞 林若竹 淮稳霞 姚艳霞
(国家林业与草原局森林保护学重点实验室 中国林业科学院森林生态环境与保护研究所 北京 100091)
新疆野生苹果(Malussieversii)(蔷薇科)属第三纪孑遗植物,是现代栽培苹果品种的祖先,是我国乃至世界十分珍贵的自然种质资源,在野生种质资源利用和生物多样性保护方面具有非常重要的价值(林培钧等, 2000)。新疆野生苹果仅分布于中亚地区,是我国新疆西天山野果林最主要的建群种,占野果林总面积的90%。然而,近年来,新疆野生苹果林出现大面积死亡现象,使这种珍稀种质资源面临着严重的威胁,其中,以苹小吉丁(Agrilusmali)(鞘翅目Coleoptera: 吉丁甲科Buprestidae)为主的病虫害是一项非常重要的影响因子(崔志军, 2018; Bozorovetal., 2019)。
苹小吉丁在国外分布于俄罗斯、蒙古和朝鲜半岛,在我国分布于黑龙江、辽宁、山东、河北、陕西、四川、甘肃、青海、新疆、内蒙古、河南等十多个省(自治区)(王智勇, 2013; EPPO, 2018),以幼虫蛀食寄主树木枝干而造成危害,枝条被害处表皮呈黑褐或深褐色,稍下凹,受害枝条常常干枯,主要危害苹果属(Malus)和李属(Prunus)植物(Bozorovetal., 2019; EPPO, 2018)。由于苹小吉丁在新疆西天山野果林内暴发,已有超过80%的野生苹果树受到苹小吉丁的危害(崔志军, 2018)。根据伊犁地区林业部门的统计,截至2015年,受害严重的新源县野生苹果枯死率高达80%,局部地区100%,苹小吉丁为害被认为是引起新疆野生苹果死亡的重要生物因子(Bozorovetal., 2019;季英等, 2004; 王智勇, 2013)。
事实上,昆虫伴生的真菌类群常常加速了害虫对寄主植物的危害。伴生菌是由昆虫携带、传播到寄主植物上,并对寄主植物产生不利影响的病原真菌(叶辉,1997),也称为虫道真菌(杨群芳等, 2008)。迄今为止,昆虫伴生真菌研究最广泛的是涉及小蠹科(Scolytidae)昆虫的种类,其与小蠹科伴生关系最密切,研究最深入的真菌当属长喙壳类(吕全, 2007; Kostovciketal., 2015; Luetal., 2010; 2011; 2016)。其次,象甲科(Curculionidae)昆虫的伴生真菌也有研究报道,相比小蠹与真菌的紧密伴生关系,除了极少数能与真菌形成互惠的共生关系外, 象甲和真菌之间的伴生关系主要是松散的、随机的(Drenkhanetal., 2017; Viiri, 2004; Robertetal., 2013a; 2013b)。此外,极少数天牛种类如断眼天牛(Tetropium)据记载也与真菌伴生,主要是长喙壳类真菌(Wingfieldetal., 1987; Johnyetal., 2012)。吉丁甲科的昆虫种类鲜有伴生真菌的报道,截至目前,仅发现Agrilusgraminis是一些黄萎病真菌(Verticillium)的活性载体(Rizieroetal., 2016)。
调查、分析昆虫伴生真菌多样性,了解其群落结构有助于探讨害虫、伴生真菌与寄主植物三者的相互关系。本研究运用真菌学和群落生态学方法,对苹小吉丁伴生真菌类群及其多样性进行调查研究,旨在了解和探明苹小吉丁优势伴生菌群类群,为探讨苹小吉丁及其伴生真菌与新疆野生苹果之间的相互作用和危害过程中是否存在伴生真菌协同发生作用奠定基础。
1 材料与方法
1.1 研究区概况
新疆野生苹果在我国境内主要分布在天山西部伊犁河谷地区海拔 1 200 ~1 600 m 的阴坡、半阴坡和河谷地带,该地区属北温带大陆性半干旱气候区,气候温和,年平均气温10.4 ℃,年积温为2 700~3 500 ℃,全年日照时数2 532~2 722 h,年降水量315.6~798.3 mm,年均无霜期156天。新疆野生苹果伴生有野生樱桃李(Prunuscerasifera)、野杏(Armeniacavulgarisvar.ansu)、准噶尔山楂(Crataegussongarica)、天山桦(Betulatianschanica)等树种,林下伴生有灌木新疆忍冬(Loniceratianschanica)、黑果小檗(Berberisatrocarpa)、 腺齿蔷薇(Rosaalbertii)和覆盆子(Rubusidaeus) 等,草本植物有鸭茅(Dactylisglomerata)、新疆党参(Codonopsisclematidea)、苦苣菜(Sonchusoleraceus)、野芝麻(Lamiumbarbatum)、早熟禾(Poaspp.)、羊茅(Festucaovina)、麻叶荨麻(Urticacannabina)和大麻(Cannabissativa)等(李利平等, 2011; 马闯等, 2018; 张高, 2013)。
1.2 样地设置
新疆野生苹果在伊犁河谷地区呈散生或团状分布,伊犁哈萨克自治州的新源县、巩留县、霍城县、伊宁县均有分布,其中新源县和巩留县野生苹果分布最多且受苹小吉丁危害最重。故本研究选取目前苹小吉丁危害最严重的库尔德宁镇阿勒马赛(43.23°N,82.85°E)、阔克塔力村(43.25°N, 82.82°E)和七十二团八连(43.24°N, 82.77°E)为采样地,每个样地大小为100 m×100 m。
1.3 取样方法
1.3.1 幼虫、蛹及坑道取样方法 在苹小吉丁幼虫出蛰危害季节(5—9月),每5~7天在样地内随机选取50个带虫枝条(长50 cm,基径3~6 cm),带回实验室剥取幼虫、蛹及坑道。由于幼虫和蛹所处环境大致相同,故把二者合并在一起分析。
1.3.2 成虫取样方法 在成虫羽化高峰期使用捕虫网捕捉成虫,随后将每头成虫单独放入1.5 mL无菌离心管中待用。
1.4 真菌分离与培养
1.4.1 虫体栖息真菌分离 从受害枝条剥取活体幼虫和蛹立即置于麦芽提取物琼脂培养基(麦芽浸粉20 g、琼脂20 g,去离子水1 000 mL)中,用无菌镊子使其在培养基上滚动2圈后取出虫体,于25 ℃恒温培养箱中培养约1周后,适时挑取不同形态特征的菌落,转入PDA培养基上进行纯化培养和菌种保藏。
将活体成虫切成5 mm × 5 mm的小块,用 1.5%次氯酸钠处理1 min、无菌水漂洗3次,并置于MEA培养基中,于25 ℃恒温培养箱中培养约1周后,适时挑取不同形态特征的菌落,转入PDA培养基上进行纯化培养和菌种保藏。
1.4.2 坑道栖息真菌分离 小心切下幼虫取食部位的新鲜韧皮或蛹室木质部组织,将其剪成小块,立即接种到培养基上,并于25 ℃恒温培养箱中培养,当组织材料周围生出菌丝时,挑取形态不同的菌落,转入 PDA培养基上进行纯化培养和菌种保藏。
1.5 菌株 DNA 提取以及目标基因片段的 PCR 扩增及测序
纯化菌株在MEA培养基上培养1周后,刮取约1 g菌丝,根据CTAB 法提取纯化菌株的DNA,PCR扩增采用ITS1和 ITS4通用引物。扩增产物由上海美吉生物公司测序,序列经BioEdit等软件进行拼接处理,在NCBI基因库进行同源性比对分析,确定分类地位。
1.6 数据分析
相对多度(Pi)=某一伴生菌的分离株数/总伴生真菌的分离株数×100%。如果某一真菌类群在群体中的度Pi>1/S,则该类群为优势菌群,反之,则该类群为次要类群,S为群体中类群总数(Camargo,1992)。
选用Jaccard 指数和Bray-Curtis指数分析不同分离材料之间伴生真菌物种组成的差异性,具体计算过程通过R语言 vegan软件包中的vegdist函数完成。
另外,选取Shannon-Wiener指数Pielou均匀度和Margalef 丰富度指数来分析物种多样性。Shannon-Wiener多样性指数和Pielou均匀度指数具体计算过程通过R语言 vegan软件包中的diversity函数完成,Margalef 丰富度指数用Excel计算。
2 结果与分析
2.1 苹小吉丁伴生真菌种类及其数量
共分离得到菌株703株,隶属真菌57属(表1),其中:链格孢属(Alternaria)获得214株,镰刀菌属(Fusarium)97株,壳囊孢属(Cytospora)55株,小穴壳属(Dothiorella)49株,其余各属菌株数量在40株以下。
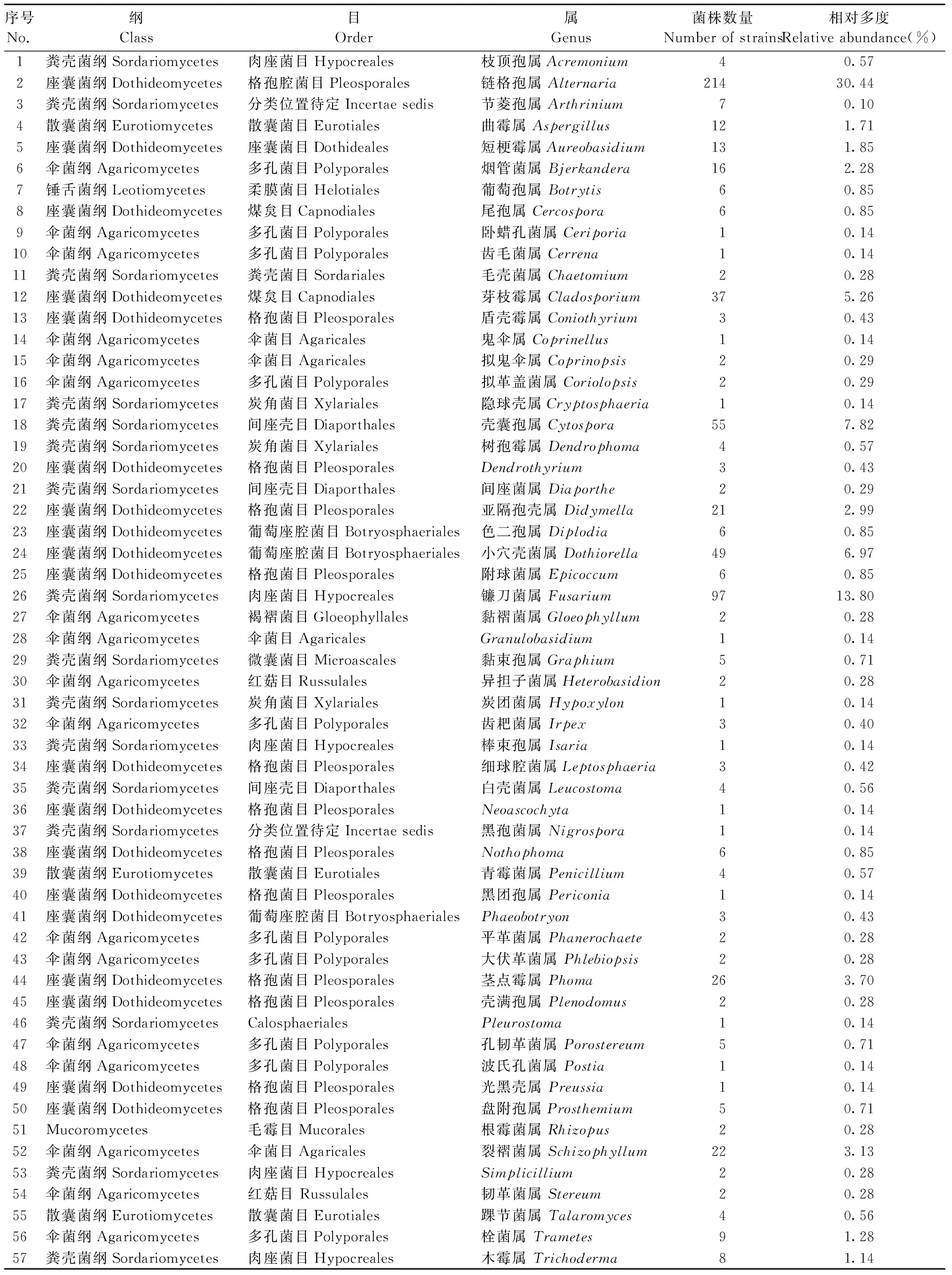
表1 苹小吉丁伴生真菌区系组成及数量①Tab.1 The genera composition and the relative abundance of associated fungi with A. mali
在这些真菌类群中,处于优势地位(相对多度Pi>1/S=1/57)的有10属(图1),所包含菌株数量占总菌株数量的78.13%。其中:链格孢属相对多度最大,其次是镰刀菌。其余8个优势属相对多度在2%~8%范围内,从大到小依次为壳囊孢属、小穴壳菌属、芽枝霉属(Cladosporium)、茎点霉属(Phoma)、裂褶菌属(Schizophyllum)、亚隔孢壳属(Didymella)、烟管菌属(Bjerkandera),短梗霉属(Aureobasidium)最小,仅为1.85%。
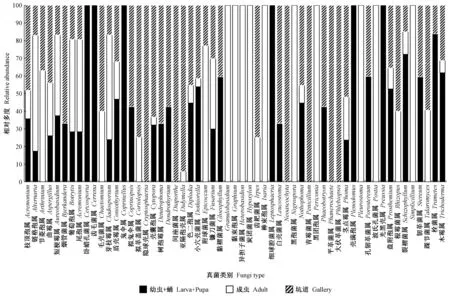
图2 不同分离材料伴生真菌的相对多度Fig.2 Relative abundance of the associated fungi from the different isolation materials
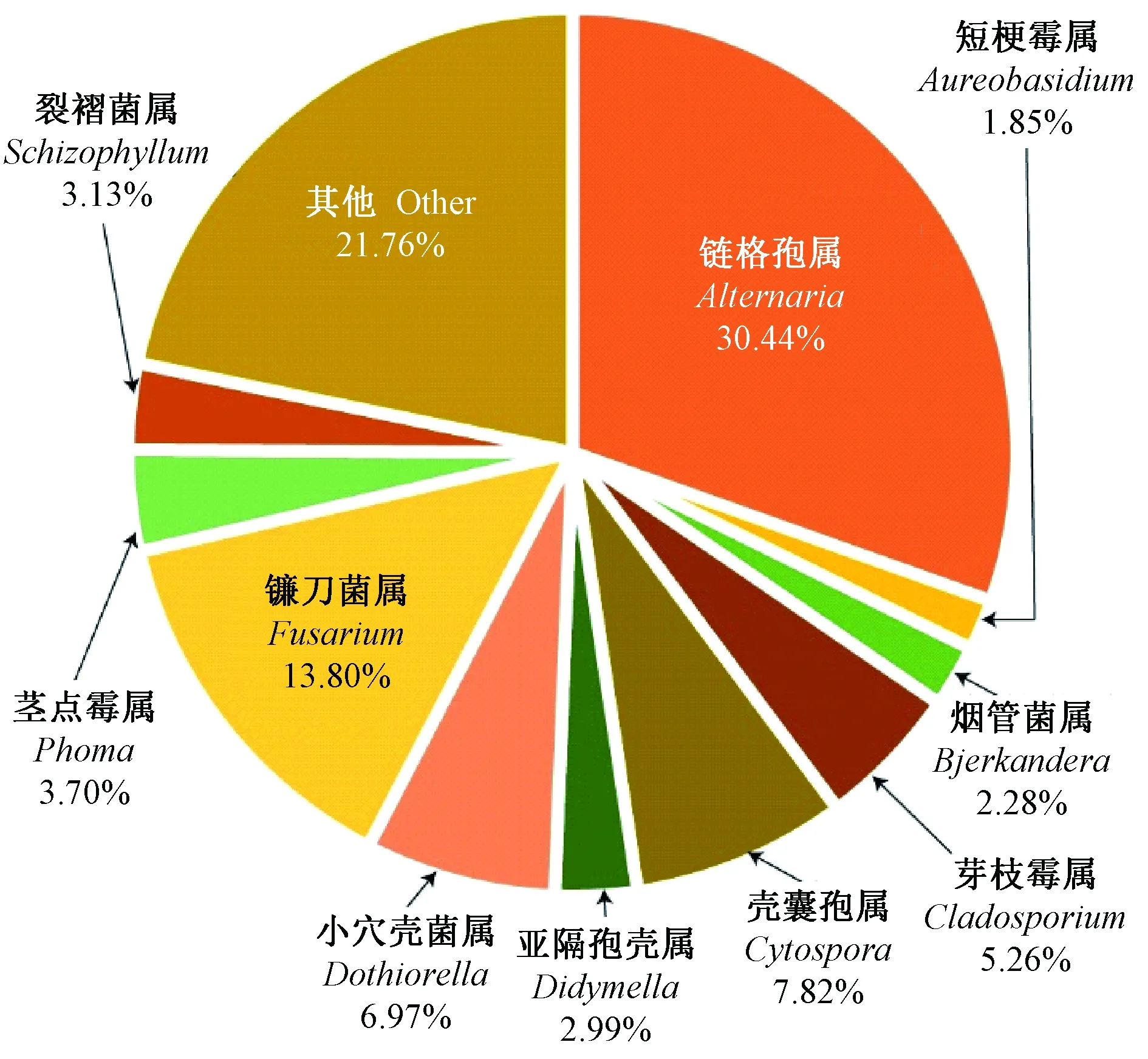
图1 苹小吉丁伴生真菌优势属的相对多度Fig.1 Relative abundance of dominant genera of associated fungi with A. mali
2.2 不同分离材料伴生真菌类群的分布
不同分离每个伴生真菌类群的相对多度如图2所示。从同一分离材料的所有伴生类群分布看,幼虫+蛹体表共分离得到菌株151株,隶属于34属,链格孢属、芽枝霉属、壳囊孢属、亚隔孢壳属、小穴壳菌属、镰刀菌属、裂褶菌属和栓菌属为优势属,小穴壳菌属相对多度最高,为13.91%,其次是链格孢属(13.21%),再为镰刀菌属(11.92%); 成虫共分离到菌株328株,隶属37属,其中链格孢属、芽枝霉属、镰刀菌属、茎点霉属为优势属,链格孢属相对多度最高,为50.30%,镰刀菌属次之(15.85%),芽枝霉属(8.23%)在镰刀菌之后; 坑道共得菌株224株,隶属40属,其中链格孢、烟管菌属、壳囊孢属、亚隔孢壳属、小穴壳菌属、镰刀菌属、茎点霉属和栓菌属为优势属,壳囊孢属相对多度最高,为16.96%,然后是链格孢(12.95%)和镰刀菌属(12.05%)。由此可见,链格孢和镰刀菌属在幼虫+蛹、成虫和坑道中均大量存在,特别是链格孢在成虫中的相对多度表现的非常突出,高达50.30%。
此外,从真菌类群来看,3种分离材料均具有各自的伴生真菌类群,如细球腔菌属(Leptosphaeria)、壳满孢属(Plenodomus)、卧蜡孔菌属(Ceriporia)等6属仅存在于幼虫+蛹的伴生真菌类群中,黏束孢属(Graphium)、异担子菌属(Heterobasidion)、炭团菌属(Hypoxylon)等10属仅存在于成虫中,隐球壳属(Cryptosphaeria)、青霉菌属(Penicillium)、平革菌属(Phanerochaete)等6属仅存在于坑道中。
2.3 不同分离部位伴生真菌组成的差异性
苹小吉丁各分离部位除具各自特异性伴生真菌外,还拥有共同伴生真菌类群,如图3所示。在获得的57属伴生真菌中,幼虫+蛹与坑道共有的伴生真菌为27属(45.76%),成虫与坑道共有的伴生真菌为26属(45.61%),幼虫+蛹和成虫共有的伴生真菌为20属(35.10%),而为上述3种分离部位所共有的伴生真菌达19属(33.33%)。
另外,Jaccard指数和Bray-Curtis指数反映出的结果是不一致的(表2),其中:幼虫+蛹与成虫的Jaccard相异性指数和Bray-Curtis相异性指数较大,而幼虫+蛹与坑道的相异性指数均较小。对于Bray-Curtis相异性指数,幼虫+蛹与成虫、成虫与坑道较为接近,都在0.6~0.7之间,但相比幼虫+蛹与坑道(0.333)差异较大;对于Jaccard指数,幼虫+蛹与坑道、成虫与坑道较为接近(0.4~0.5),但相比幼虫+蛹与成虫(0.608)差异较大。综合分析2种相异性指数发现,幼虫+蛹和成虫之间相异性较大,如果只考虑物种的有无,成虫和坑道的相异性较小(0.490),而如果不仅考虑物种的有无,而且还考虑不同物种的相对丰度时,成虫和坑道之间的相异性较大(0.609)。
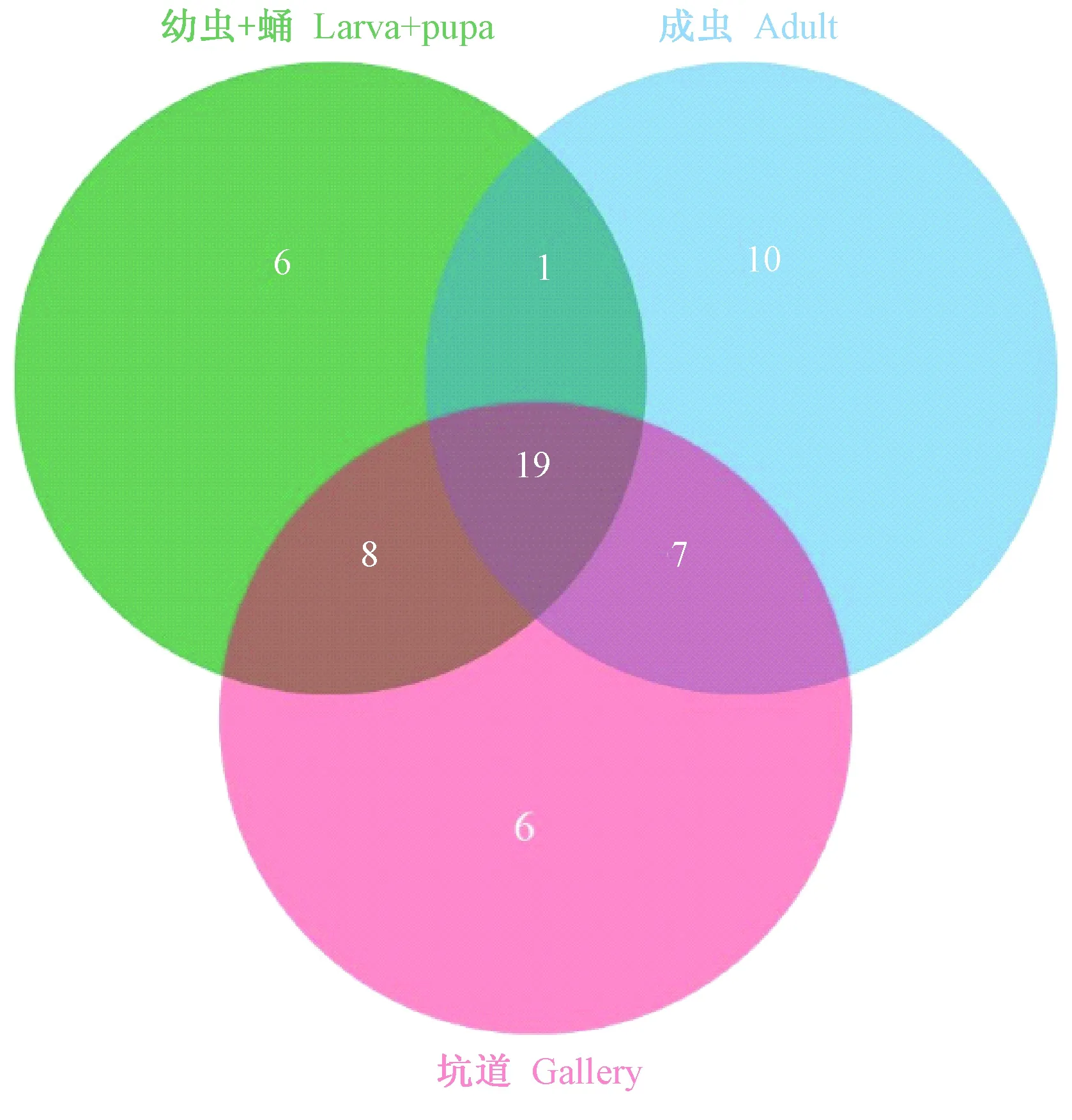
图3 不同分离材料真菌类群组成的数量关系Fig.3 Venn diagram of the fungi groups for different isolation materials
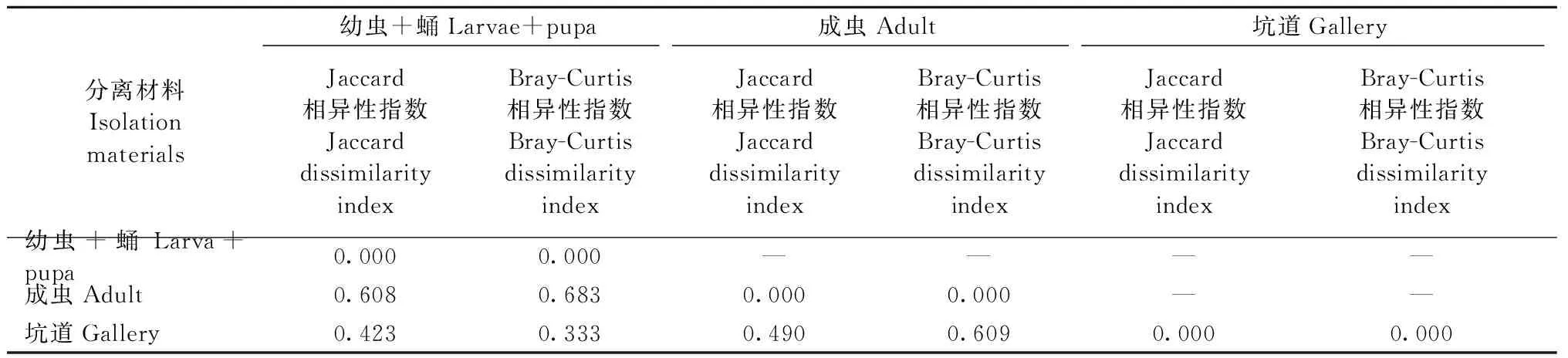
表2 不同分离材料之间伴生真菌类群相似性比较Tab.2 Paired comparison of β dissimilarity indices for different isolation materials
2.4 不同分离部位伴生真菌多样性分析
由坑道分离的伴生真菌Shannon-Wiener多样性指数最高,幼虫+蛹体表略低于坑道,成虫最低,并与前二者相差较大(图4)。幼虫+蛹体表伴生真菌类群数量不如成虫多,但Shannon-Wiener指数却大于后者,说明前者类群中个体分配上的均匀性(Evenness)较好(图4)。从Pielou 均匀度指数看,幼虫+蛹最高,坑道次之,成虫则明显低于前二者,说明幼虫+蛹伴生真菌属与属之间个体数量差别不大,而成虫伴生真菌属与属之间个体数量差别较大。而从Margalef丰富度指数来看,三者中坑道最高,成虫最低。成虫3种指数均为最低,说明成虫携带真菌种类以少量真菌类群为主。
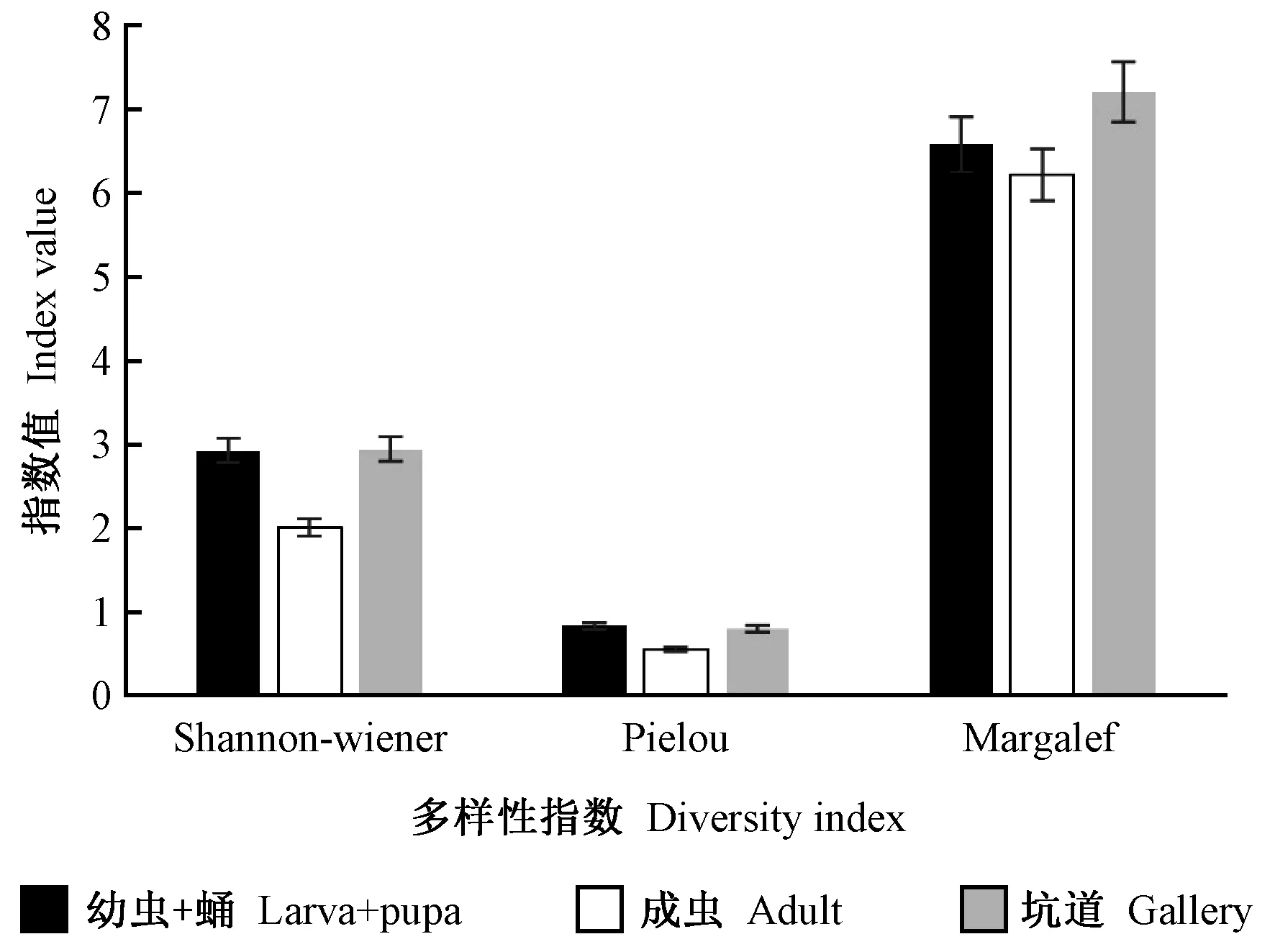
图4 不同分离材料之间伴生真菌的多样性比较Fig.4 Comparison of the diversity of the associated fungi from different isolation materials
3 讨论
本文首次系统性研究了苹小吉丁伴生真菌类群及其多样性,从苹小吉丁幼虫、蛹、成虫和坑道中共分离鉴定了57属真菌类群,隶属于6纲18目。上述57属中处于优势地位的有10属,其中链格孢属的数量最多。尽管不同分离材料伴生真菌分布不同,然而从处于优势地位的真菌类群及其相对多度来看,链格孢属在幼虫+蛹、成虫和坑道中均大量存在,特别在成虫中相对多度表现非常突出,高达50.30%,可能是因为链格孢属的分生孢子容易释放,并且通过气流传播,在空气中广泛存在,苹小吉丁成虫喜欢在天气晴朗、温度稍高的时候绕树冠飞行,这为其接触并携带链格孢提供了便利条件。链格孢主要引起寄主植物叶斑和叶枯病,也可引起果腐病,如苹果果腐和苹果果斑(王婧等, 2017; George, 2009)。由于许多链格孢属真菌是腐生真菌,仅在死亡或者正在腐烂的植物组织上生长,因而在一些衰老的叶片、花瓣和成熟的果实上最常见(George, 2009),但从苹小吉丁患病幼虫虫体上也分离到链格孢[细链格孢(Alternariatenuis)](孔婷婷等, 2017),因此,苹小吉丁和链格孢是否复合危害野果树尚待进一步研究。
镰刀菌是本研究获得的另外一个优势真菌类群,在3种分离材料中均处于优势地位。在小蠹虫伴生菌研究中,镰刀菌与小圆胸小蠹(Euwallaceaefornicatus)形成虫菌共生体方胸小蠹-镰孢菌共生体(Euwallacea-Fusarium)(Freemanetal., 2013; O’Donnelletal., 2016; Kassonetal., 2013),造成寄主机械损伤、枝干枯死和木材腐烂(Walgama, 2012)。这些危害状在受苹小吉丁危害的树体上均有表现,并且在害后发生褐变的野生苹果心材部分也发现了多种镰刀菌(Chengetal., 2019),由此推测苹小吉丁与镰刀菌在引起野生苹果枝枯的过程中均产生了影响。
此外,能引起多种植物溃疡病的重要真菌类群小穴壳菌属和壳囊孢属(张星耀等,1999; 王秀娟等, 2019; 王斌等, 2018)在苹小吉丁幼虫+蛹和坑道中大量存在,前者在幼虫+蛹中表现突出,相对多度最高(13.91%),后者在坑道中相对多度高达16.96%,因此,不能排除这些真菌类群对野果林产生的危害。与此同时,本研究从苹小吉丁成虫中分离到黏束孢属(Graphium)真菌,该属许多种类与小蠹虫形成伴生关系(Jacobsetal., 2003; Yamaokaetal., 2009); 但苹小吉丁成虫与小蠹虫生活习性完全不同,前者不进入寄主植物内部生活,仅在补充营养时取食寄主,因此,苹小吉丁成虫与该真菌的相互关系还有待进一步探讨。
从不同分离部位的伴生真菌数量来看,苹小吉丁危害的坑道中真菌类群最多,这可能与真菌本身对环境的要求有关,受到创伤的植物组织给真菌的侵入提供了有利条件(叶建仁等, 2011); 幼虫+蛹真菌类群最少可能是因为健康的虫体只允许那些对其有利的真菌类群存在。从Shannon-Wiener多样性指数分析的结果来看,由坑道分离的伴生真菌Shannon-Wiener 多样性指数最高,与坑道真菌类群最多相一致,但是幼虫+蛹体表伴生真菌Shannon-Wiener指数远远高于成虫,前者伴生真菌种类数量不如后者多,但该指数却大于后者,说明前者伴生真菌种类中个体分配上的均匀性好于后者,也就是说幼虫+蛹体表伴生真菌属与属之间个体数量差别不大,而成虫伴生真菌属与属之间个体数量差别较大,Pielou 均匀质指数的分析结果证实了这一点,同时Margalef 丰富度指数分析结果也与上述结果相一致,三者中坑道最高,成虫最低。由于上述多样性指数与地理位置、环境条件及树木微环境息息相关(王立祥等, 2017),成虫3种多样性指数均最低,而幼虫+蛹和坑道3种指数相对差异较小,因此推测可能是幼虫+蛹与坑道微环境相似而与成虫环境条件相差较大导致。此外,成虫3种指数均为最低,也说明成虫携带真菌种类以少量真菌类群为主,且主要携带真菌种类所占比例在伴生真菌类群中较大。
与此同时,苹小吉丁上述3种分离材料除拥有共同伴生真菌类群外,各自存在某些特异性较强的真菌种类。幼虫+蛹和成虫均与坑道存在共有伴生真菌,说明伴生真菌与坑道环境关系密切(Adamsetal., 2007)。另外,尽管成虫和幼虫相异性较大(Jaccard=0.608,Bray-Curtis=0.683),但二者共有伴生真菌类群也达20属,表明虽然幼虫+蛹和成虫所生活环境差别较大,但一些真菌类群和苹小吉丁关系十分密切,可能依靠苹小吉丁传播扩散。
4 结论
苹小吉丁伴生真菌种类较多,且不同分离材料的真菌类群种类和数量存在差异。在这些真菌类群当中,链格孢和镰刀菌是苹小吉丁整个生活史发育过程中的主要伴生真菌类群,而镰刀菌的种类常常引起植物枝枯和褐腐,同时,引起植物溃疡病的重要真菌类群——小穴壳菌属和壳囊孢属在幼虫+蛹和坑道中大量存在,可能存在某些与苹小吉丁关系十分密切的伴生真菌类群,说明对野果林造成危害的不仅仅是苹小吉丁,真菌对野果林的衰退也可能产生了较大的影响。因此,对退化野果林生态保育与健康调控要从多个方面考虑,把真菌所致病害和虫害综合治理相互结合起来。
