林火导致树木死亡的作用机制和影响因素的研究进展*
2020-08-14韩大校王晓红丛日征邸雪颖蔡慧颖张吉利
韩大校 韦 睿 王晓红 丛日征 邸雪颖 杨 光 蔡慧颖 张吉利
(1. 中国林业科学研究院寒温带林业研究中心 哈尔滨 150086; 2. 东北林业大学林学院 哈尔滨 150040)
林火可以通过烧伤树木的组织和器官导致其直接死亡,也可以通过影响树木水分、养分的代谢途径以及诱发病虫害等导致其延迟死亡,而这些作用往往耦合在一起,使树木的死亡机制异常复杂(Michaletzetal., 2007; 2008)。在林业工作中,树木的烧伤程度和死亡率对林火发生后开展抢救性采伐、森林火灾损失评估和火烧迹地恢复具有重要的实践意义(Hoodetal., 2010)。此外,适当强度的火烧不但可以减少积累在森林地表的可燃物,还可以增强生态系统的自我调节能力,计划烧除用火时机和强度的选择都需要火后树木死亡相关基础研究的支撑(舒立福等, 1998; 王立夫等, 2006)。本文从林火的作用途径及影响因素、火后树木死亡的原因和机制、树木死亡的判定及预测等方面进行综述,以期为今后相关的理论研究和实践工作提供科学参考。
1 林火对树木的作用途径及影响因素
林火的产生的高温是导致树木死亡的主要外因,在开放的燃烧环境中一般会产生800~1 000 ℃的高温,而阴燃产生的高温通常为600~750 ℃(Michaletzetal., 2007)。从个体角度分析,林火发生时树冠和树干处于地上的开放环境,而根系分布在土壤之中,燃烧时的热负荷及传热方式决定了其烧伤程度(Petersonetal., 1986; Michaletzetal., 2008)。例如,林火中树皮表面的温度一般为400 ℃左右,而土壤表层的温度约为150~160 ℃,并随土壤深度锐减(Swezyetal., 1991;Vanderweideetal., 2011)。在微观水平上,木质可燃物颗粒的热解始于200 ℃左右,而后热解速率逐渐提高并在300~400 ℃时达到峰值(Jonesetal., 2004)。一般认为60 ℃是植物组织细胞的致死温度(Dickinsonetal., 2004),但即使在45~60 ℃的范围内,只要持续时间足够长同样能达到致死效果(Bovaetal., 2005; Vanderweideetal., 2011)。研究发现,植物细胞的死亡速率随受热时间呈指数上升(Dickinsonetal., 2004; Butleretal., 2010):
S(t)=e-kt。
(1)
式中,S(t)为t时刻细胞的存活量,k为细胞死亡速率系数(s-1),t为受热时间(s)。但也有研究将致死温度持续1 min以上认定为植物组织死亡的参考标准(Dickinsonetal., 2004)。
1.1 林火对树冠的作用途径及影响因素
1.1.1 林火对树冠的作用途径 林火主要通过热辐射和热对流对树冠传热,而对流过程占主导地位,由于枝、叶和芽等器官形态各异、分布离散不均,对树冠传热过程模拟的难度最大(Michaletzetal., 2007)。火线强度和火焰高度常被用于反映林火能量的释放(Keeley, 2009; Alexanderetal., 2012)。火焰高度不但与树冠烧伤高度(Crown scorch height)密切相关,还可用于推算树冠烧伤体积比(Percentage crown volume scorch)(Van Wagtendonk, 1983)。当火焰高度低于树冠时,树冠承受的高温主要来源于下方火羽流的自然对流过程(Van Wagner, 1973;Cruzetal., 2006; Michaletzetal., 2007)。Peterson等(1986)将过火后死亡的树冠部分理想化为截去头部的圆锥体,则树冠的死亡体积比(ck, fraction of crown volume killed)为:
(2)
式中,hk为树冠的死亡高度(m),ht为树高(m),cl为树冠长度(m)。
在Van Wagner的经典模型中,树冠烧伤高度(hs, m)主要取决于3方面因素,即火线强度(I, kW·m-1)、风速和环境气温(Ta, ℃)(Van Wagner, 1973):
(3)
式中,k为与风速相关的经验系数,公式(3)的假设前提为叶和芽的温度与火焰或火羽流的温度一致。由于k在不同树种和燃烧环境间的变异较大(Alexanderetal., 2012),Michaletz等(2006)使用集总热容法对其进行了校正和扩展,增加了该模型在不同环境中的适用性:
(4)
式中,C为火羽流比例常数,Tc为树冠组织致死温度(℃),g为重力加速度(m·s-2),Pa为环境空气密度(kg·m-3),Cp为标准大气压下空气的比热(J·kg-1℃-1)。
当地表火的火线强度或火焰高度超过某一临界值后就会转化为树冠火,Van Wagner(1977)通过半经验模型对这一临界值(I0, kW·m-1)进行了推导:
I0=[0.01×CBH×(460+26×FMC)]1.5。
(5)
式中,CBH为树冠基部高度(m), FMC为叶片含水率(%)。
Cruz等(2006)建立的树冠引燃模型将树冠可燃物颗粒受到的净热通量(q″)表述为对流热通量(qc)、辐射热通量(qr)和该可燃物颗粒向外耗散热通量(ql):
q″=qc+qr-ql。
(6)
各部分拆分计算后对应的方程如下:
(7)

dyFdyfdxFdxf。
(8)
式中,Lf和Wf分别为可燃物颗粒的长度(m)和宽度(m),WF为火焰宽度(m),Df为火焰深度(m),Dx为火焰与可燃物颗粒的水平距离(m), ε为火焰的辐射率,TF为火焰温度(K),α为辐射吸收系数(m-1),x、y、z为三维坐标距离(m),z为垂直坐标,S为无穷小表面元之间的距离(m)。通过该模型可以精确计算林火中树冠上叶、芽和枝某一时刻热量及温度的理论值。
1.1.2 影响树冠烧伤程度的主要因素 虽然火后叶和芽的死亡因树种而异,但当火焰覆盖树冠后大部分树种会在几个月内死亡(Michaletzetal., 2007)。像美国黄松(Pinusponderosa)和杰弗里松(Pinusjeffreyi)等芽苞较大的树种,在树冠被烧伤90%以上甚至叶片全部褐化的情况下仍具有分生能力,并在次年存活下来(Hansonetal., 2009)。因此,结合火后次年的萌芽和展叶情况可以避免对树木死亡率的高估(Hoodetal., 2010)。
树冠的组成和形态主要影响传热的边界条件[例如公式(3)中的k](Michaletzetal., 2007)。如果将公式(4)中空气的比热(Cp)替换为芽或叶的平均比热则可用该公式区分芽和叶的烧伤高度(Michaletzetal., 2008)。相对而言,阔叶树种的球状树冠更易引发树冠火,但其火后不定芽的萌蘖繁殖能力更强、生长速度更快(Angersetal., 2011)。生长季布满叶片的冠层可以降低火羽的对流速度,这种效应还受叶片形状、分布密度影响(Michaletzetal., 2006)。而针叶树种较强的自然整枝能力、较厚的芽鳞,以及锥形的树冠和较高的树冠基部高度等特征都更有利于它们在火后存活(O’Brienetal., 2010)。
1.2 林火对树干的作用途径及影响因素
1.2.1 林火对树干的作用途径 树干主要通过辐射和对流2种途径受热,树皮因良好的绝热性会在林火中对形成层和木质部起到保护作用(Bovaetal., 2005; Michaletzetal., 2007)。Bova等(2009)通过逆向推导将辐射热量与对流和传导产生的热量分离,准确计算形成层吸收的总热量,但并不适用于树皮含水率较高或被严重烧伤的情况。
一般将形成层在林火中的受热过程抽象为半无限大物体的一维导热问题,并利用傅里叶传热定律对其进行解析(Martin, 1963; Petersonetal., 1986):
(9)
式中,Tc为形成层致死温度(℃),T0为树皮外表面温度(℃),T1为火焰瞬时温度(℃),x为树皮厚度(cm),α为树皮热扩散率(cm2·min-1),τ为受热时间(min),f为高斯误差积分函数。通常树皮的热扩散率在不同树种间基本恒定(0.06 cm2·min-1),约为同等密度木材的20%(Martin, 1963)。公式(9)的假设前提为外部热源和传热介质具有同质性,但实际火焰和树皮属性的异质性很大,并且树干在周向上的受热也并不均匀。例如,在热气流涡旋效应的作用下,树干背风向的烧伤程度一般更高(Gutselletal., 1996)。雷诺系数(Re)可用于反映风速与树干背风向涡旋形成的关系(Gutselletal., 1996):
(10)
式中,d为圆柱体直径(m),U为风速(m·s-1),v为空气运动黏度(m2·s-1)。Re较小时,流场中流速的扰动会因黏滞力而衰减,流体呈层流流动状态; 反之,流体呈紊流流动状态,此时背风向更容易形成涡旋。
Jones等(2004; 2006)通过模拟燃烧试验建立的一维传热模型(FireStem)对形成层温度的模拟精度可达75%~80%,但其缺陷在于对不同树种的预测敏感性差异较大。Chatziefstratiou等(2013)在此基础上建立的二维模型(FireStem2D)将从圆周向圆心的传热过程进行离散化的数值模拟:
(11)
式中,t为受热时间(s),ρ为木材或树皮的密度(kg·m-3),k为热导率(W·m-1K-1),cp为木材或树皮的比热(J·kg-1K-1),T为温度(K),θs:θn为周向积分;rw:re为径向积分。但热量在轴向上的传热速率受树干液流、木质部导管(或管胞)形态结构的影响未被纳入FireStem2D,这些问题解决后即可对树干的传热过程进行三维模拟,目前FireStem2D已在美国林务局的推动下应用,并在我国的部分树种中得到验证(Weietal., 2019)。
1.2.2 影响树干烧伤程度的主要因素 胸径较大的树木可以降低环状烧伤的概率,并且因其叶片和芽苞所处的位置也相对更高,对地表火的抗性更强(Brandoetal., 2012)。而树皮较薄的桉树(Eucalyptusspp.)会分配更多的生物量在树干的生长上,并使处于高位的潜伏芽(epicomic bud)在火后尽快萌生出新的枝条和叶片(lawesetal., 2011)。但对美国黄松的研究发现,由于大树基部积累了更多的凋落物和腐殖质,导致火焰的驻留时间更长、火强度更高,因此成、过熟林过火后比中、幼龄林的死亡率更高(Kolbetal., 2007; Hoodetal., 2010)。
树皮的厚度相对于含水率、密度和表面纹理结构等因素对树皮绝热性能的影响作用更大,是评估树木火后能否存活的关键参数(Lawesetal., 2011; Weietal., 2019)。有研究表明,树干形成层在林火中受热达到峰值温度的时间与树皮厚度的平方呈显著的线性相关关系(Hoodetal., 2008),而树皮厚度与胸径又具有较好的线性关系,因此可将胸径作为树木耐火能力的指示变量(Petersonetal., 1986; Vanderweideetal., 2011)。把树皮厚度与含水率、抽提物变量综合分析,可以为耐火树种的筛选提供依据并制定相应的经营措施(田晓瑞等, 2001; 李世友等, 2009)。树皮的相对厚度(树皮厚度与胸径的比值)因种而异,在同一树种的不同生长阶段也随会因分配策略不同而变化,相对厚度可以通过反映异速生长关系间接指示树木的抗火性(Lawesetal., 2014)。
但在讨论树皮的绝热性时,有必要区分内树皮和外树皮的作用。内树皮的含水率较高,因此可以吸纳更多的热量,对形成层传热起到缓冲作用,但火头过后同样会增加散热时间(Lawesetal., 2011; Vanderweideetal., 2011; Odhiamboetal., 2014)。有研究认为,含水率较高的内树皮在总树皮厚度中占有更大的比例可能是小径级树木在火后存活的重要原因(Van Mantgemetal., 2003; Schaferetal., 2015)。外树皮由脱落的死细胞积累构成,含水率一般仅为内树皮的20%,多数研究者仍认为外树皮的绝热性能更强,但较厚的外树皮也会造成阴燃,从而增加形成层的受热时间和强度(Harrington, 2013)。
1.3 林火对根系的作用途径及影响因素
1.3.1 林火对根系的作用途径 不同于树冠和树干,通过土壤进行热传导是林火向根系传热的主要途径(Michaletzetal., 2007)。地表可燃物和土壤在结构组成上较强的异质性导致很难确定根系受热的边界条件(Massmanetal., 2010)。目前多用细根生物量及活性的变化间接反映林火对树木根系的影响(Hartetal., 2005;楚旭等, 2013)。例如,在对脂松(Pinusresinosa)细根进行不同温度和处理时间的热胁迫后发现,细根1周内死亡率的持续变化主要受胁迫时间的影响而非温度,造成细根死亡的最低温度为52.5 ℃(Zelezniketal., 2004)。根部的韧皮部比树干更薄,因而其形成层对温度的敏感性更高(Kolbetal., 2007)。同叶片类似,在研究中一般假设细根(<2 mm)可以在外界温度变化时瞬间达到热平衡(Michaletzetal., 2007)。在对美国白杨(Populustremuloides)的研究中发现,其细根暴露于49 ℃下持续75 min后,韧皮部细胞几乎全部死亡(Dickinsonetal., 2004)。
土壤中含有大量的水分因而也具有较大的热容,对美国黄松的研究发现,计划火烧后0~5 cm的土层温度高于60 ℃的状态仍可持续5 h以上,此后1~5个月内细根的生物量会减少50%~75%(Swezyetal., 1991)。在分析林火中土壤温度变化时要分别考虑持续受热阶段和散热阶段,因为处于散热阶段的土壤仍能保持较高的温度。Enninful等(2008)将这2个阶段表述如下:
(12)
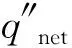
1.3.2 影响根系烧伤程度的主要因素 根系的烧伤程度主要取决于可燃物属性和根系分布的特征(Zelezniketal., 2004; Wrightetal., 2008)。粗木质残体、有机土壤等可燃物的阴燃可以持续几小时甚至几天,这种半开放环境中的燃烧会导致树木细根的大量死亡(Varneretal., 2009)。当阴燃发生时,潮湿的腐殖质层由隔热介质转化为潜在的热源(Butleretal., 2010)。但低强度的地表火和控制火烧并不会造成腐殖质和土壤的阴燃,也不会导致土壤的斥水性(Ryanetal., 1991; Swezyetal., 1991; Varneretal., 2009)。在多数地表火中,5~10 cm以下的土壤很少能达到60 ℃,但大部分细根分布在腐殖质层及浅层土壤中(Wrightetal., 2008),并且越接近土壤表面的细根生命周期越短,也更容易在林火中死亡(Zelezniketal., 2004)。根系的受热还与土壤的含水率、质地、孔隙度和密度等属性密切相关(Busseetal., 2010),持有足够水分的土壤可以有效缓冲林火向深层传递的热量(Wrightetal., 2008)。Busse等(2010)研究表明,当土壤含水率为20%或更高时,超过致死温度的土壤深度可控制在2.5 cm以内,而当土壤含水率在5%左右时,相应的深度会超过10 cm。
2 树木过火后的死亡机制及影响因素
林火发生时,急剧升高的温度会对过火树木产生一系列的影响,而树木的响应会从物质交换和代谢产物等方面表现出来(Michaletzetal., 2007; Butleretal., 2010)。通过对这些响应机制的研究有助于对火后树木直接死亡和延迟死亡的深入理解和预测。
2.1 分生组织和叶片
如果树木过火后树干的韧皮部和形成层全部死亡,树木不仅会失去径向生长能力,光合作用合成的有机物质也无法通过筛管输送到根部(Ryanetal., 1988; Michaletzetal., 2012)。根部失去养分供给后只能利用存储的可溶性糖和淀粉等物质维持生存,存储养分耗尽后细根对水分和无机盐的吸收能力逐渐丧失,最终导致树木死亡(Michaletzetal., 2008; Odhiamboetal., 2014)。有研究发现,只要树干在周向上形成层存活的比例高于15%~20%,树木就可以维持养分输送(Ducreyetal., 1996),留存的形成层细胞会继续分化并在烧伤部位形成火疤(Gutselletal., 1996)。
林火发生时树冠的蒸腾作用强度会急剧增加,饱和水气压差(VPD,vapour pressure deficit)的瞬时升高会对叶片组织造成不可逆的伤害(O’Brienetal., 2010)。虽然叶片是进行光合作用的主要器官,但火后树木死亡与否主要取决于芽而非叶,如果顶芽和侧芽全部死亡即代表树木失去潜在的分生活力(Petersonetal., 1986)。
2.2 气体交换和光合速率
林火中气孔的关闭可以缓解蒸腾作用造成的水分流失,降低气穴(Cavitation)现象的形成概率,但也会限制CO2通过气体交换参与光合作用,从而造成光合速率下降(Ducreyetal., 1996; Kavanaghetal., 2010)。Kavanagh等(2010)认为,环状烧伤后因树干液流减弱引起的气孔关闭相对于韧皮部养分传输能力下降对树木的致死作用更强,前者会使树木在5个月内死亡。林火的矿化作用会增加叶片的养分供应,一些阔叶树种的每日最大光合速率会增加30%~50%,但仅是暂时性的(Reichetal., 1990)。Carlo等(2016)也发现,由于火后栎树(Quercusspp.)林下的氮素含量较高,林内乔木的光合作用速率也会比火前增强。树冠上层的光合能力通常较中、下层更强,因此即使在林火中失去中、下层75%的树冠,树木仍能存活(Fowleretal., 2010)。还有研究发现,火后斜叶桉(Eucalyptusobliqua)萌生新枝的叶片单位面积蒸腾速率约是对照组的3.4倍,而2 m以下萌生新枝条的叶片要比树冠上层的老枝叶片具有更高的气孔导度和光合速率(Nolanetal., 2014)。
气孔的导度和调节能力决定了叶片对VPD变化的响应速度,而这在不同树种叶片间的差异却很大(Reichetal., 1990; Westetal., 2016)。总体上被子植物对叶片气孔的调控过程比裸子植物更加复杂,并且裸子植物叶片多覆被蜡质以防止水分流失,在林火中需要更多的热量融解蜡质,因此被子植物更容易在林火中形成气穴栓塞(Kavanaghetal., 2010)。美国黄松叶片的气孔对环境温度和土壤干旱的响应具有较高的敏感性,这种特性不但能减少气穴栓塞的形成,还会在火后提高单位叶面积的光合速率(Salaetal., 2005)。
2.3 木质部水分传导能力
通常认为树干环状烧伤所导致的养分胁迫是火后树木死亡最主要的直接原因,但近年研究发现过火后木质部的水分传导会降低40%~80%,在短期内对树木的致死作用更强(Michaletzetal., 2012; Westetal., 2016; Lodgeetal., 2018)。树干液流密度可以作为预判火后树木死亡的依据,地中海松(Pinushalepensis)在环状烧伤1周后树干液流密度会下降为轻度烧伤和对照组的60%~70%(Ducreyetal., 1996)。O’Brien等(2010)在对长叶松(Pinuspalustris)的研究中发现,林火发生后树干液流会持续下降直至根系死亡生物量达到过火前的30%~40%,超过这一阈值后剩余根系的水分输送能力便难以维持生存。
在火羽流的作用下,空气的VPD会很快超过6 kPa的临界值,最高可达270 kPa,而叶片的气孔却不能及时关闭,蒸腾作用产生强大的负压使木质部水分的张力骤然增大,外界空气从导管(或管胞)壁渗入形成气穴栓塞从而阻碍水分经由木质部向上传导(Kavanaghetal., 2010; Michaletzetal., 2012)。如果林火发生时土壤中的水分充足(例如树木生长在河岸),气穴栓塞形成的机率会大幅降低(Kavanaghetal., 2010)。叶片、侧枝和根系形成气穴栓塞的概率要大于主干,这些位处远端的组织相对于主干的传热速率更快,在高温中承受的负压也会更强,此外导管或管胞壁结构更利于气穴栓塞形成(Kavanaghetal., 2010; Westetal., 2016; Lodgeetal., 2018)。West等(2016)发现,火后气穴栓塞会更多地形成于树木远端的芽和叶柄中,以保护主干和枝条的水分传导功能,而气孔栓塞的形成概率主要受木质部导管或管胞、叶片表皮细胞及气孔的热敏感性影响。同理,下层灌木木质部形成气穴栓塞的可能性更大,而在土壤水分恒定的前提下,上层乔木的水分利用率会因此增加,这种现象会在树干液流通量的变化上得到印证(Kavanaghetal., 2010;Carloetal., 2016)。Battipaglia等(2016)研究表明,温度在50~300 ℃范围内、高度不超过1.5 m的火焰不但不会对意大利伞松(Pinuspinea)的气孔导度、木质部结构等造成不利影响,还可以借此清除林下共生的草本和灌木,提高该上层乔木的水分利用率。
还有研究发现,当树干和枝条内部温度升至60~90 ℃时,木质部导管的木质素会发生玻璃化转变,导管壁软化、形变并失去支撑和固定能力,在树液张力的作用下导管会萎缩甚至崩裂(Michaletzetal., 2012; Westetal., 2016; Bäretal., 2018)。这种不可逆的形变同样会降低树干水分的传导能力,是气穴栓塞理论之外另一关于木质部水分传导率下降的重要假说。但细胞壁形变假说的验证建立在水浴试验的基础上,在真实的林火环境中能否成立仍待进一步研究(Bäretal., 2018; Hoodetal., 2018)。
干旱、土壤冻融和昼夜变化都可能造成少量气穴栓塞的形成,但会随时间推移被自身修复,并不会对树木整体的水分输导产生明显的影响(Kavanaghetal., 2010; Westetal., 2016; Hoodetal., 2018; Lodgeetal., 2018)。因此,对枝条和叶片水势的测定最好在黎明前进行,以便尽可能让根系和土壤的水势处于平衡态(Lodgeetal., 2018)。另外,由于树干基部积累的可燃物一般更多,即使低强度地表火也可能烧伤根颈部从而导致根系发生严重的气穴栓塞(Lodgeetal., 2018)。
2.4 生物化学响应
林火产生的高温不仅会破坏树木的组织结构,还会影响其生理代谢过程。通过对火后树木代谢产物的研究可以深入认识树木的烧伤和死亡机制。
2.4.1 活性氧及酶代谢 林火对植物组织的热胁迫可被视为一种氧化胁迫过程(Alexouetal., 2014)。高温胁迫后活性氧会大量累积并在1周左右达到峰值,细胞膜流动性和渗透性的损伤体现为丙二醛(MDA)含量的显著升高(王荣等, 2012; Alexouetal., 2014; Escandnetal., 2016)。为了减少活性氧造成的伤害,植物细胞会通过调节合成超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)等中和活性氧类物质,这些酶类物质可以在过火树木的叶片中被大量检测到。
2.4.2 植物激素代谢 植物激素信号的传导是树木受到高温胁迫后生理响应的起点。Escandn等(2016)对辐射松(Pinusradiata)的研究发现,脱落酸(ABA)和水杨酸(SA)的含量在过火后的3天内持续增加并达到峰值,它们主要用于调节气孔的关闭以及合成丙二醛; 而吲哚乙酸(IAA)和细胞分裂素(CKs)等主要在树木过火后的恢复期合成,对调节气孔张开、提升光合速率方面作用更明显。抗坏血酸(AsA)可以提高树木组织的抗氧化能力,在火烧处理2 h后就可在叶片中检测到抗坏血酸浓度的显著升高(Alexouetal., 2014)。
2.4.3 萜烯类化合物代谢 萜类(单萜类、倍半萜类和二萜类等)是构成树脂的主要成分,可以将其浓度变化作为树木受高温胁迫后短期内的指示变量(Alessioetal., 2004)。Valor等(2017)发现:地中海松和欧洲赤松(Pinussylvestris)过火前后单萜类物质质量分数变化不显著, 但耐火性更强的欧洲黑松(Pinusnigra)火烧处理24 h之后的单萜类物质质量分数显著低于火烧前,而分别对过火后24 h和1年松针的取样分析都表明,所有的单萜和二萜类物质质量分数都随树冠烧伤体积百分比的增加而升高。Lavoir等(2013)发现,反复火烧会显著促进南欧黑松(Pinuslaricio)萜烯类物质的合成,而单次火烧对萜类物质含量的影响并不明显。Kelsey等(2017a)研究表明,α蒎烯作为火后美国黄松释放的化学信息素能够吸引大量的红脂大小蠹(Dendroctonusvalens),其在被蛀食的韧皮组织和边材中的质量分数分别为164.2 μg·g-1和71.5 μg·g-1,分别是未蛀树干一侧的4.9倍和1.6倍。
2.4.4 酚类化合物代谢 Alonso等(2002)研究发现,75%的海岸松(Pinuspinaster)树冠受到火烧处理2个月后,针叶中浓缩单宁酸、叶绿素和类胡萝卜素等多酚类物质的含量会明显升高。对南欧黑松的研究也发现,在控制火烧3个月后其针叶中总酚含量会显著增加,而单酚含量的变化并不明显(Cannacetal., 2009)。因此总酚类物质也可被视为树木过火后短期代谢的响应指标,例如在春季计划烧除后的总酚含量要高于秋季,这间接说明在秋季实施计划烧除可以减轻对树木的不利影响(Cannacetal., 2009)。同样,南欧黑松在过火3个月后针叶中总黄酮类物质含量增加60%,并在约1年后恢复到过火前的水平(Cannacetal., 2011)。而酚类化合物含量的增加可能源于高温胁迫后苯丙氨酸解氨酶活性的提高,是细胞抗氧化作用的体现(Lavoiretal., 2013)。
2.4.5 乙醇代谢 当植物组织温度升至30~40 ℃时,增强的呼吸作用会形成低氧胁迫环境; 当温度升至40~50 ℃时,细胞膜和线粒体膜的选择透过性降低,电解质开始外渗,丙酮酸脱羧化酶和乙醇脱氢酶活性的增加会导致乙醇大量合成积累(Kelseyetal., 2017b)。对美国黄松的研究发现,乙醇同样是诱发大小蠹(Dendroctonusspp.)虫害的化学信息素,过火2周后边材和韧皮部合成乙醇的浓度显著升高,并与树干和树冠的烧伤程度正相关(Kelseyetal., 2003)。火后被蠹虫蛀食树皮中的乙醇的质量分数是对照组的几百甚至几千倍,仅少量可以通过树干液流和自身代谢消耗,而大部分被释放到空气中(Kelseyetal., 2017a; 2017b)。
3 影响火后树木死亡的其他因素
3.1 过火季节
过火季节对树木死亡率的影响主要体现在树木、虫害的生理节律和潜在的燃烧环境2个方面。有研究认为,秋季地表积累的可燃物更多,相应火强度更高,因而树木死亡率更高; 也有研究认为,树木储存的养分在春季处于较低水平,因而春季和早夏的林火对树木幼芽、幼叶和细根的伤害更大,也更容易引发虫害,导致树木的死亡率比秋季高(Swezyetal., 1991; Schwilketal., 2006)。Harrington(1993)发现,在树冠烧伤程度相近的前提下,美国黄松在春、夏季过火后的死亡率约是秋季的2.5倍,但这种差异主要体现在小树上,而胸径大于30 cm树木的死亡率差异并不显著。Mchugh等(2003)发现,秋季过火后树木感染虫害的比率最低(11%)、夏季次之(19%)而春季最高(41%)。这可能是因为春季正处树木代谢旺盛时期,过火后会在低氧胁迫环境中大量合成乙醇和蒎烯类化合物(Kelseyetal., 2017a; 2017b)。也有研究发现,在相近的火烧强度下,美国黄松在晚季林火后的死亡率更高; 冷杉属(Abiesspp.)的小树在早季林火过后更容易受到大小蠹的蛀食,但总体上早、晚季的火后死亡率差异并不显著(Schwilketal., 2006)。
3.2 群落组成与结构
林分密度可以从潜在的火行为和资源竞争2方面影响火后树木的死亡过程(Hoodetal., 2018)。草本、灌木和乔木等不同群落的空间分布直接影响着可燃物载量、分布、含水率等属性和林内的气象条件(Stephensetal., 2012)。此外,密度更高的林分也更容易在火后遭受虫害的侵染从而提高树木的延迟死亡率(Schwilketal., 2006)。经历中、低强度林火后,更新的针叶实生苗与邻近的阔叶根蘖苗在生长过程中是竞争还是协同关系决定群落未来的空间格局(Owenetal., 2017)。而生长速率是衡量树木活性、环境胁迫与资源竞争的综合参数,在过火强度相近的前提下,在竞争中处于劣势、生长相对缓慢的树种死亡率更高(van Mantgemetal., 2018)。
3.3 树脂的分泌
大部分针叶树种都会在过火后分泌更多树脂,油状树脂不但可以在物理上抵御虫害的蛀食,还能对大小蠹等虫害及寄生真菌起到化学毒杀作用。树脂在夏季的生长高峰期流动性最强,其分泌量、树脂道的尺寸与树木的胸径、生长速度呈正相关(Hoodetal., 2015)。树脂道的面积和分布密度也是衡量树脂分泌能力的重要参数,当树木受到虫害攻击或干旱胁迫时会把计划分配给径向生长的养分转投入到促进树脂合成的相关代谢中(Kaneetal., 2010)。研究发现,低强度的火干扰会促使美国黄松的树脂道组织更加发达,这可以有效降低其在火后由病虫害所致的延迟死亡率(Hoodetal., 2015)。
4 火后树木死亡的预测
目前,对火后树木死亡率的调查仍以表观变量为主,叶片、树皮等可燃物的过火痕迹主要源于纤维素、半纤维素和木质素等聚合物在热解和燃烧阶段的变色、炭化和龟裂,这些痕迹能独立反映火行为特征和树木组织的烧伤情况,也可联合起来建立树木死亡率函数模型(Woolleyetal., 2012)。
4.1 表观经验预测
4.1.1 独立指示变量 反映烧伤程度的独立指示变量可以分为2类: 一类反映火焰高度、火强度等火行为参数,主要包括地表炭化码(ground char)、树干炭化码(bole char)、树干熏黑高度(bole scorch height)和树冠烧伤高度等; 另一类反映树木组织烧伤程度,包括树冠烧伤体积(crown scorch volume)、树冠死亡体积(crown consumption volume)以及其他针对形成层和细根的观测变量,前者适用于计划烧除方案的制定,而后者更适用于过火后树木死亡率的预测(Fowleretal., 2004)。形成层死亡等级(CKR, cambium kill rating)是指在胸径(1.4 m)处截面按坡向或方向划分的4个象限中的形成层已经死亡的象限数,主要通过形成层位置的颜色、湿润度和柔韧性判定(Ryanetal., 1988)。CKR是被广泛使用的火后树木死亡评级标准,但取样过程仍比较复杂,Hood等(2008)在CKR的基础上提出了在野外更具操作性的树皮炭化码(bark char codes),分为未烧伤、轻度、中度和重度烧伤4级(Hoodetal., 2008)。树皮炭化码更适用于树皮较薄的树种,而对树皮较厚的树种适用性较差(Hoodetal., 2008)。
4.1.2 综合评级法 分级检索表是美国进行火后树木死亡率评估的常用工具,首先检索过火树木的树种和径级,再根据过火前的立地条件和过火后树木的烧伤程度对死亡率综合评级,预期死亡率较高的树木会被尽早采伐以降低损失,而检索表的编制依赖于常见树种火后死亡率预测模型的确立(Scottetal., 2002)。
4.2 统计经验模型
目前,二项分布Logistic模型是最重要的火后树木死亡率统计模型(Woolleyetal., 2012)。Fowler等(2010)使用分段Logistic模型确定了美国黄松的死亡阈值,即当树冠的烧伤体积比大于85%,或树冠死亡体积比大于40%,或死亡体积比为5%~40%且烧伤体积比大于50%时,可以判定树木死亡。虽然树木的死亡多发生在火后次年,但多数研究会使用火后3~4年内的死亡率调查数据建模,以保证延迟死亡的树木也能被纳入样本(Harrington, 1993; Fowleretal., 2004)。有研究发现,针叶树种多在火后的第一年内死亡,而大部分(约82%)阔叶树种的死亡发生在过火一年之后(Catryetal., 2010)。
与树冠相关的指标变量是入选Logistic模型最多的一类(Hoodetal., 2010),其中又以树冠烧伤体积比对模型的解释能力最强,树干熏黑高度次之(Keyseretal., 2006)。Ganio等(2017)也发现,由树冠烧伤体积比和CKR建立的Logistic模型对于火后美国花旗松(Pseudotsugamenziesii)死亡率的预测精度高于80%。在使用非火后指示变量建立的Logistic模型中,树皮厚度通常对模型的贡献率最高(Ryanetal., 1988; Vanderweideetal., 2011; Brandoetal., 2012)。
4.3 半物理模型
统计模型无法解释过火树木的死亡机制,在建模样本对应的总体中预测精度较高,但受树种和燃烧环境等因素的约束也很大,Peterson等(1986)结合树冠和树干的烧伤程度提出了半物理模型:
(13)
式中,Pm为死亡率;ck为树冠死亡体积比(0≤ck≤1);τc为形成层受热致死所需时间(min);τL持续燃烧时间(min)。ck可由公式2推算,τc可由公式(9)中的τ推算,τL可由下式计算(Petersonetal., 1986):
(14)
式中,wi为第i个径级的可燃物载量(g·cm-2);cfi第i个径级的可燃物燃烧消耗比例。在地表火中,当τc≤ 0.5τL时,表明会发生树木环状烧伤并死亡; 而当τc≥4.5τL时,代表形成层未被烧伤,树木的死亡仅取决于树冠的烧伤程度(Petersonetal., 1986)。τL和cfi除受可燃物颗粒的含水率和表面积体积比影响外还受风速影响,同等条件下风速越大形成层的烧伤程度和树干内部的气穴栓塞程度越高,反之树冠叶片的烧伤更严重(Kavanaghetal., 2010)。
4.4 生物过程模型
生物过程模型包含林火的传热过程和生理变量的参数化,因而比统计模型和半物理模型的普适性更强(Michaletzetal., 2008; Butleretal., 2010),但模型构建过程更复杂,这在一定程度上限制了过程模型的应用(Michaletzetal., 2007;Woolleyaetal., 2012)。由于目前养分、水分胁迫和病虫害等原因导致的树木死亡过程仍不清楚,过程模型仅被用于树木直接死亡率的预测(Michaletzetal., 2008)。Michaletz等(2008)通过傅利叶传热定律将高温与树木的侧生分生组织(公式9)和顶端分生组织(公式4)的死亡联立并将死亡率Pm表示为:
(15)
式中,Nn、Nt分别是芽的死亡数量和总数量;xn、xba分别是树皮组织的死亡深度和总厚度。当xn/xba<1时,U(xn/xba)=0, 反之,U(xn/xba)=1。该过程模型没有考虑树皮和腐殖质的阴燃并且仍局限于树干受热的均一性假设。过程模型的建立首先要明确树木组织受热的边界条件,如树干、根系或土壤受热时表面的温度或热量分布情况,这些数据可以通过热电偶和热流计直接测得,也可以通过热力学公式估算(Butleretal., 2010; Chatziefstratiouetal., 2013)。
5 结论与展望
林火是森林生态系统中的一个关键生物物理学过程,深入的认识这一过程对于我国的天然林资源保护和林火管理工作至关重要。国外对该领域的研究综合了气象学、传热学和植物生理学等多个学科,并且开展了大量的试验和监测(Bäretal., 2018)。但相关研究与我国实际情况存在较大差距,例如,国外的研究对象多为成、过熟林,而我国多为采伐和火干扰后密度较高的中、幼龄林; 国外研究对象多为美国黄松、花旗松等针叶树种,而我国多为温带阔叶树种; 国外研究多建立在计划烧除试验基础之上,而我国很少开展林内的计划烧除试验。
综上所述,对火行为和热量传递的量化是科学评估和预测火后树木死亡率的基本前提,而分生组织的烧伤程度是决定火后树木死亡率的关键,与树冠烧伤程度相关的变量指示性较强,结合火后次年的树木萌芽、展叶情况可以更准确的估计树木死亡率。Logistic模型和分级检索表是目前预测和评估火后树木死亡率最常用的方法,而过程模型是未来的研究趋势。光合速率下降和韧皮部传导功能的减弱共同造成了养分胁迫,而木质部的气穴栓塞和软化形变是导致水分传输失败的潜在原因,并且后者在短期内的致死作用更强。激素、酚类、萜烯类和乙醇等代谢产物是反映树木烧伤程度的关键生化指标。此外,过火季节、群落结构和树脂的分泌也是影响树木死亡率的重要因素。
最后,对本领域仍存在的部分问题总结并提出以下建议: 现有火行为模型的空间尺度大、预测结果分辨率低,无法与树木个体水平上的过火痕迹变量相匹配,而这些痕迹变量同样难以与树木内部的生理变化建立对应联系(O’Brienetal., 2010),这类联系的建立有赖于对火行为、树木组织的热物理属性和异速生长关系的协同研究(Michaletzetal., 2008)。只有量化热量的传递才能明确叶、芽、形成层和根系等死亡的关键参数,并解释树木过火后在水分、养分利用和生物化学方面的响应机制(Kremensetal., 2010; O’Brienetal., 2018)。建议通过野外原位观测和室内风洞模拟建立可靠的小尺度火行为模型,根据树冠、树干和根系不同的结构特征和边界条件采用几何近似法分别建立或改进传热模型,以实现对树木个体死亡率的定量预测。
Logistic模型的实用性较强,建议根据立地、树种、径级和林火发生时期分别建立试验数据库和死亡率模型,并将较易调查的虫害变量耦合到Logistic模型中,以便不断进行模型校验和改进,为我国火后树木死亡率检索表的编制提供依据。未来应联合多个学科建立能够反映树木死亡机制的半物理模型和过程模型,充分利用不同类型模型的优势(Hoodetal., 2018)。
建议研究我国主要树种的树冠、树干和根系过火后在响应途径和敏感程度方面的差异。利用现有的生化检测、水势测定、光合测定和同位素标记等技术手段,尽量统一取样和测定标准,以便得到更多可比较、能重复的科学数据和研究成果。
由于树木对林火的抗性在种间及种内都存在差异,可以先获取林分中不同树种和径级的比率,然后利用差分归一化燃烧率将单木死亡率模型进行尺度放大(Travis Beloteetal., 2015)。虽然林火中通过热辐射释放的能量占释放总能量的比例不到30%,但该比例基本恒定,并且产生的长波辐射可以穿透烟气,通过机载红外相机获得的高分辨率数据可以推算火场的辐射能量密度,并结合所占总能量的比例关系反演火场热能的三维分布情况(Smithetal., 2017)。而机载激光雷达已经可以分析可燃物在小尺度水平上的空间异质性(Kremensetal., 2010)。这些监测数据的优势在于它们都是有度量单位的连续变量,因而可以与净光合速率等连续变量建立函数联系(Smithetal., 2016)。
