补喂亮氨酸对伊犁马血浆氨基酸及其主要相关代谢物浓度的影响
2020-08-11艾毕肯赖晓鹤张文杰张仕琦郑文祥欧阳文祁居中杨开伦
艾毕肯, 赖晓鹤, 张文杰, 韩 明, 刘 律,张仕琦, 郑文祥, 欧阳文, 祁居中, 杨开伦*
(1.新疆农业大学动物科学学院,新疆肉乳用草食动物营养重点实验室,新疆乌鲁木齐 830052;2.新疆伊犁哈萨克自治州昭苏县种马场,新疆昭苏 835000)
亮氨酸(Leu),又称为白氨酸,是必需氨基酸之一,动物机体自身不能合成,只能在日粮中获取(Lee等,2019)。Leu在肌肉中在支链氨基酸氨基转移酶的催化下能发生可逆反应,生成α-酮异己酸(KIC),然后进入肝脏,在肝脏细胞溶胶中被KIC二氧化酶催化生成β-羟基-β-甲基丁酸(HMB)(Arazi等,2018)。 Van 和 Nissen(1992)研究表明,Leu的代谢产物KIC和HMB同Leu一样具有调控蛋白质代谢的作用,抑制蛋白质降解主要通过KIC刺激胰岛素的分泌以及HMB抑制泛素-蛋白酶体系统来进行。
研究表明,食物缺乏的大鼠口服Leu可以刺激其骨骼肌蛋白质的合成,这不依赖于循环中胰岛素的浓度,而是通过促进eIF4F的形成来进行的(Anthony 等,2000);Boutry 等(2013)的研究结果表明,给新生猪肠外输注Leu能增加肌肉蛋白质的合成;Suryawan 等(2012)为新生猪以 148 μmol/kg的输注率从颈静脉滴注Leu 10 min,然后再以400 μmol/kg·h持续滴注1 h,结果表明Leu通过诱导mTORC 1及其下游通路的激活来促进新生猪mRNA的翻译,使肌肉蛋白质合成;Bro·jer等(2012)给马匹服用葡萄糖和Leu时,马匹运动后6 h内的血浆胰岛素浓度明显高于给马匹注射生理盐水时的血浆胰岛素浓度,但是,对马匹体内糖原合成速率没有影响;而在人类补充Leu的研究中,独木舟运动员通过6周的饮食补充Leu,显著提高了上半身力量和耐力(Crowe等,2006)。本试验选择年龄为1岁的伊犁马为研究对象,探究通过在日粮添加不同水平的Leu对伊犁马血浆氨基酸浓度以及Leu及其代谢产物KIC、HMB吸收规律的影响,为Leu在伊犁马饲养中的应用及Leu需要量的确定提供科学依据。
1 材料与方法
1.1 试验时间及地点 本试验于2019年9月18日至2019年10月8日在新疆伊犁哈萨克自治州昭苏县种马场进行。
1.2 试验设计及饲粮组成 试验采用单因素多水平试验设计方法。挑选体重为(321.68±25.7)kg、年龄为1岁的伊犁马20匹,分为四组,设为对照组、试验Ⅰ组、试验Ⅱ组、试验Ⅲ组,每组5匹(3♀+2♂),试验马匹日粮组成及营养水平见表1,精料补充料及干牧草中氨基酸含量见表2。粗饲料每匹(7.00±0.80)kg/d,精料补充料 3 kg/d。 在此基础上,试验Ⅰ、Ⅱ、Ⅲ组分别补喂 17、34、51 g Leu(Leu购自乌鲁木齐市新华锐饲料有限公司,纯度98%,含量≥90%)。玉米和次粉以1:1的比例混合均匀,再与Leu以1:1的比例制作成颗粒的形式与精料补充料混合饲喂。所有试验马匹均在单独的马厩饲养,自由饮水。试验期21 d。每天定时打扫马厩,更换干净的垫料。
1.3 样品的采集及指标测定
1.3.1 样品的采集 在试验的第21天进行样品的采集。分别在饲喂前,饲喂后1、2、4 h用5 mL肝素钠抗凝管于试验马匹进行颈静脉采血。3500 r/min离心10 min,收集上清血浆,分装于2 mL冻存管中,-20℃冷冻保存。
1.3.2 指标测定 饲喂后2 h的血浆样品指标:20种氨基酸、3-甲基组氨酸(3-MH);饲喂前、饲喂后 1、2、4 h 血浆样品指标:KIC、HMB。 以上样品均送至天津普瑞华盛医学检验有限公司测定。均采用高效液相色谱串联质谱联用法测定。
1.4 数据处理 试验数据采用Excel 2010进行初步整理,再用SPSS 22.0软件中单因素方差分析(one-way ANOVA)数据的差异显著性,同时使用Duncan’s法进行多重比较。试验结果以“平均值±标准误”表示。采用正交多项式对比,检验日粮中Leu浓度增加的线性和二次效应。以P<0.05表示差异显著水平,P<0.01表示差异极显著水平。

表1 精料补充料及干牧草的营养水平(干物质基础) %

表2 精料补充料及干牧草中氨基酸含量(干物质基础)%
2 结果与分析
2.1 补喂Leu对伊犁马血浆支链氨基酸浓度的影响 由图1可知,血浆中Leu浓度随着Leu补喂水平的增加而呈上升的趋势 (拟合方程为y=4.128x+102.84,R2=0.9492)。各试验组伊犁马血浆Leu浓度均高于对照组。

图1 日粮亮氨酸补喂水平与伊犁马血浆亮氨酸浓度的关系
由表3可知,与对照组相比,试验Ⅲ组血浆亮氨酸浓度显著升高90.53%(P<0.05);试验Ⅰ组、试验Ⅱ组、试验Ⅲ组血浆异亮氨酸浓度分别降 低 32.18% (P < 0.05)、50.53% (P < 0.01)、39.27%(P<0.05);试验Ⅱ组血浆缬氨酸浓度有下降趋势(P=0.044)。
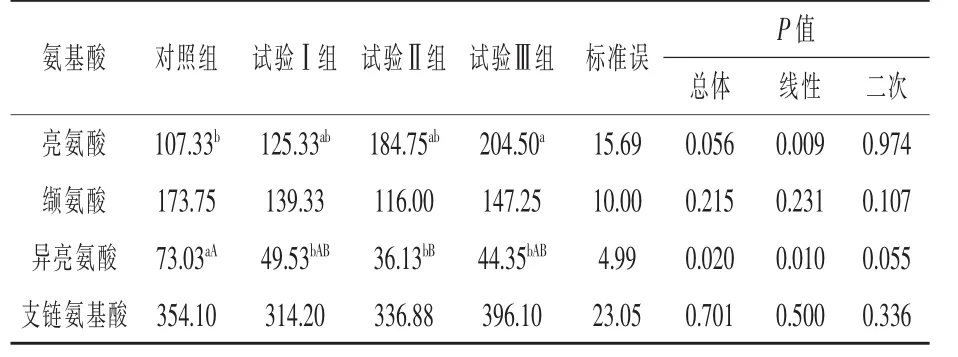
表3 补喂亮氨酸对伊犁马血浆支链氨基酸浓度的影响 μmol/L
2.2 补喂Leu对伊犁马血浆KIC、HMB浓度的影响
2.2.1 补喂Leu对伊犁马血浆KIC浓度的影响由图2可知,血浆KIC随着Leu补喂水平的增加而呈上升的趋势。不补喂Leu时,血浆KIC浓度最低,Leu补喂水平为每匹马每顿8.5~17 g时血浆KIC浓度趋于平缓,超过17 g时,血浆KIC浓度有持续升高的趋势。
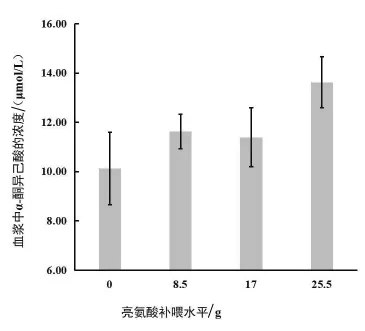
图2 日粮亮氨酸补喂水平与伊犁马血浆中α-酮异己酸浓度的关系
由表4可知,在饲喂前,与对照组相比,试验Ⅰ组、试验Ⅱ组血浆KIC浓度差异不显著 (P>0.05),试验Ⅲ组血浆KIC浓度显著升高 (P<0.05),升高了30.56%。与试验Ⅱ组相比,试验Ⅲ组血浆KIC浓度显著升高 (P<0.05),升高了35.52%。饲喂后1 h各组血浆KIC浓度差异不显著(P>0.05)。在饲喂后2 h,试验Ⅲ组血浆KIC浓度比对照组显著升高 (P<0.05),升高了43.54%。在饲喂后4 h,与对照组相比,试验Ⅰ组、试验Ⅱ组血浆KIC浓度差异不显著 (P>0.05),试验Ⅲ组血浆KIC浓度极显著升高 (P<0.01),升高了53.91%。
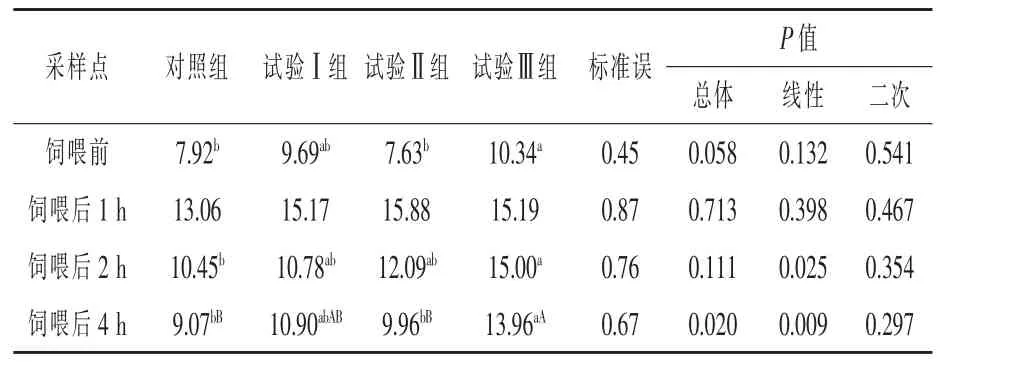
表4 补喂亮氨酸对伊犁马血浆α-酮异己酸浓度的影响μmol/L
2.2.2 补喂Leu对伊犁马血浆HMB浓度的影响由图3可知,不补喂Leu时伊犁马血浆HMB浓度最高,其随着Leu补喂水平的增加而呈下降的趋势,当每匹马每顿Leu补喂水平>17 g时,血浆HMB浓度趋于平缓。

图3 日粮亮氨酸补喂水平与伊犁马血浆中β-羟基-β-甲基丁酸浓度的关系
由表5可知,补喂Leu对伊犁马血浆HMB浓度无显著影响(P>0.05)。对照组、试验Ⅰ组血浆HMB浓度在饲喂后1 h达到峰值,试验Ⅱ组、试验Ⅲ组血浆HMB浓度在饲喂后4 h达到峰值。

表5 补喂亮氨酸对伊犁马血浆β-羟基-β-甲基丁酸浓度的影响 μmol/L
2.3 补喂Leu对血浆中非支链氨基酸、3-甲基组氨酸浓度的影响 由表6可知,与对照组相比,试验Ⅰ组血浆Glu浓度极显著降低37.07%(P<0.01),试验Ⅲ组显著降低 26.52%(P < 0.05)。
与对照组相比,试验Ⅲ组血浆3-MH浓度有下降趋势(P=0.055),血浆半胱氨酸浓度有下降的趋势(P=0.060)。与试验Ⅰ组相比,试验Ⅲ组血浆3-MH浓度有下降趋势(P=0.051)。
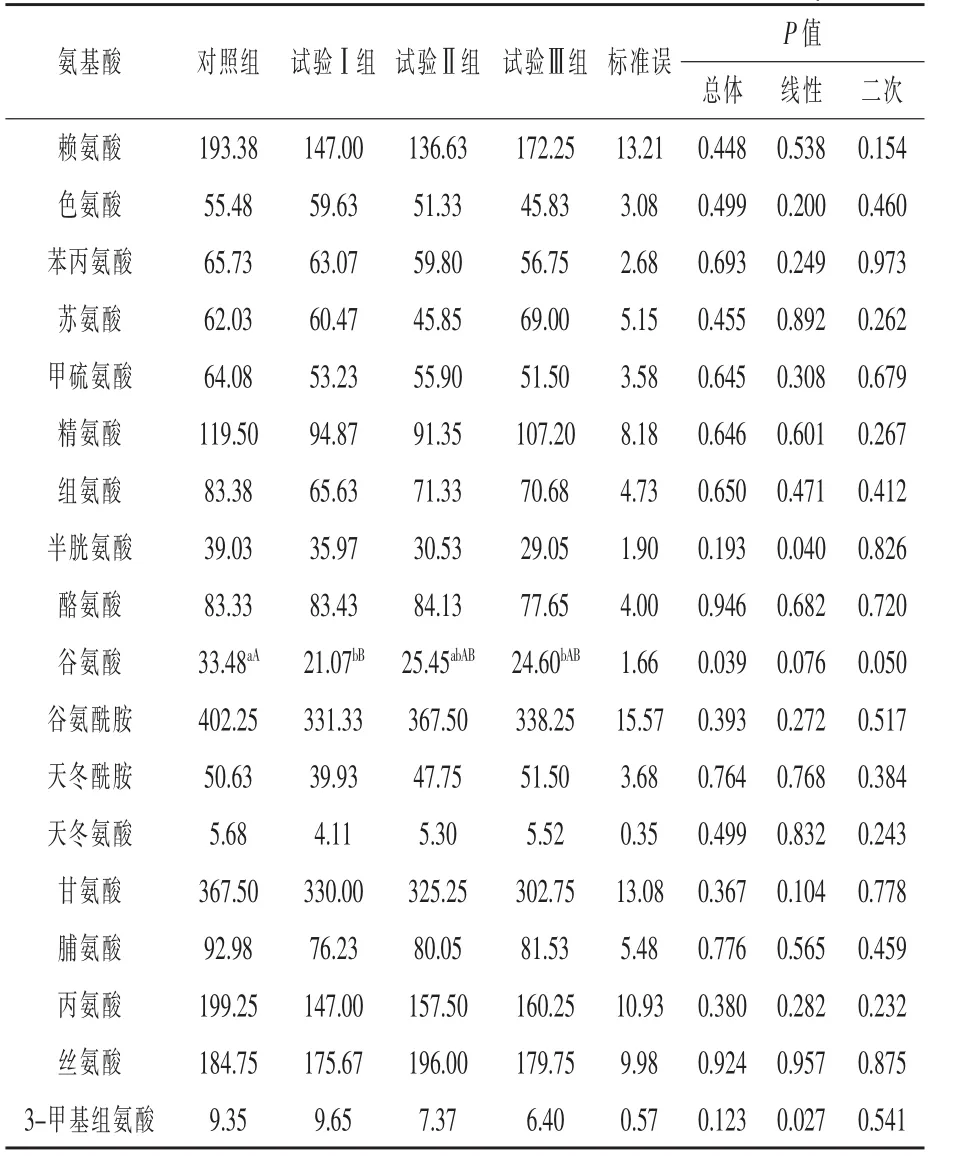
表6 补喂亮氨酸对伊犁马血浆非支链氨基酸浓度的影响 μmol/L
3 讨论
3.1 补喂Leu对伊犁马血浆氨基酸浓度的影响BCAA能促进肠道发育,促进肠道氨基酸运输和黏液蛋白的产生,调节免疫反应,刺激骨骼肌蛋白合成,研究表明,BCAA中最有效的是Leu(Escobar等,2010)。Leu具有刺激骨骼肌中蛋白质合成和抑制蛋白质降解的能力,这种促进蛋白质合成主要通过上调雷帕霉素靶蛋白复合物1(mTORC1)的活性,而抑制降解主要在组织中通过泛素-蛋白酶体途径发生 (Norton和Layman,2006)。
本试验中,补喂Leu显著提高了伊犁马血浆Leu浓度,极显著降低血浆Ile浓度,血浆Val浓度有下降的趋势,对BCAA浓度无显著影响。孙玉丽(2015)为7日龄的正常新生哺乳仔猪及宫内生长受限新生哺乳仔猪连续灌服14 d的Leu,研究结果表明,与对照组相比,给正常新生仔猪补喂Leu极显著提高了血浆中Leu浓度,极显著降低了血浆中Val和Ile的浓度,为宫内生长受限新生哺乳仔猪补喂Leu极显著升高了血浆中Leu浓度,血浆中Val及Ile浓度随着补喂的Leu水平升高有下降的趋势;Wiltafsky等(2010)为去势猪补喂Leu 34 d,共6个处理组,L100组Leu水平为11.5 g 标准化回肠可消化 Leu/kg,L150、L175、L200组的Leu水平为 L100组的 150%、175%、200%,在大于200%的组中,Leu水平为L100组的一倍,该研究的阳性对照组日粮组成与L100组相同,但Ile水平为6.3 g标准化回肠可消化Ile/kg,研究结果表明补喂Leu增加一倍的组,血浆Leu增加64%,这与本研究结果一致。这是因为Leu与其他BCAA竞争抑制。BCAA家族及Trp是由中性氨基酸转移载体 (LAT)家族中的一员LAT1转运的,这是因为含有支链和苯环侧链大分子的BCAA与其亲和力较强 (Neinast等,2019)。Leu竞争性的结合LAT1,抑制了Ile及Val在血浆 中 的 量 (Wolfe 和 Robert,2017;Louard 等 ,1990)。Leu血浆浓度的升高激活了所有BCAA的氧化代谢途径,结果导致血浆中Ile和Val降低,当单独使用Leu时,Ile和Val的可用性可能会限制肌肉蛋白质合成的速度(董千铜等,2014)。Zeitz等(2019)将450只成年雄性肉鸡平均分为三个试验组,对照组饲喂超过推荐剂量15%的Leu,L35组饲喂超过推荐剂量35%的Leu,L60组饲喂超过推荐水平60%的Leu,进行为期5周的试验,结果显示L60组的Leu既不会影响肉鸡蛋白质合成和降解的分子途径,也不会影响5周内的生产性能。随着Leu摄入量增加到所需水平,Ile和Val的浓度下降,可能是由于这些氨基酸现在正以越来越多的量用于蛋白质合成。然而,Val、Leu、Ile的比例几乎总是接近于1.6:2.2:1.0(Louard等,1990),这也可能是在本研究中Ile浓度低的原因。
本试验中,试验Ⅰ组血浆谷氨酸的浓度极显著低于对照组,试验Ⅲ组显著低于对照组。当动物体内氨基酸降解速度过高,体内产生一些过量且必须排出的氨时,氨基酸降解速度增高的信号会使转氨反应加速,从而会引起谷氨酸浓度的增高(Schachter和 David,2008)。 而本试验中血浆谷氨酸浓度降低,可能是由于马匹血浆氨基酸降解的速度下降引起氨的量降低,导致谷氨酸浓度降低。
在谷氨酰胺合成酶的催化下,谷氨酸转化成谷氨酰胺,动物体内产生的过多氨与谷氨酸结合,生成谷氨酰胺(Erich,2008),而本试验中血浆谷氨酰胺的浓度差异不显著,可能是由于该反应是可逆的,处于动态平衡中。
3-MH可用于识别蛋白质的代谢。3-MH浓度的输出用作肌肉蛋白质降解程度的指标,3-MH在肌肉降解期间释放到循环中,然后在尿中定量、不可逆地排出而不被代谢(David和Pearlstone,1996)。肌动蛋白和肌球蛋白分解过程中的组氨酸甲基化是3-MH的唯一来源(Nagasawa和Takashi,1996)。马匹是草食动物,体内的3-MH来源于饲粮中摄入的蛋白质,而蛋白质摄入对肌肉代谢的影响,导致3-MH不同程度的释放到血液中(Kochlik等,2018)。也有研究表明,马匹血浆中的3-MH浓度会受到季节的影响,主要是热应激引起的,在7~9月马匹血浆中3-MH的浓度显著增加,在此之后降低(Glade等,1990)。本试验中试验Ⅲ组3-MH浓度有下降的趋势,随着Leu补喂水平的升高,3-MH的排出下降,这可能是补喂Leu有抑制蛋白质降解的作用,还需要进一步研究。
3.2 补喂Leu对伊犁马血浆KIC、HMB浓度的影响 Leu促进蛋白质合成的作用,可能是通过其代谢产物KIC和HMB的作用产生的(Wilkinson等,2013),然而,支持这种观点的证据鲜有报道。KIC作为Leu的直接代谢产物,且在分解代谢过程中刺激蛋白质合成的能力已被研究了60多年(Kang 等,1986),研究表明,KIC 能够在不增加细胞内Leu浓度的情况下促进大鼠心肌中蛋白质合成并抑制蛋白质降解(Tischler等,1982)。HMB不仅能够提高肌肉功能,而且能够提高脂肪代谢速率并降低蛋白质分解速率,促进组织细胞修复,减轻肌肉损伤并延缓肌肉疲劳(He等,2016;Korna-sio 等,2009)。
本试验中,血浆KIC的浓度随着Leu的补喂水平呈升高趋势,在饲喂前,与对照组相比,试验Ⅲ组血浆KIC浓度显著升高。饲喂后1 h各组血浆KIC浓度差异不显著,各组都达到峰值,在饲喂后2 h各组血浆KIC浓度开始下降,与对照组相比,试验Ⅲ组血浆KIC浓度显著升高,饲喂后4 h,试验Ⅲ组血浆KIC浓度极显著升高;Wiltafsky等(2010)研究表明,为猪补喂不同水平的Leu,比Leu水平为11.5 g标准化回肠可消化Leu/kg多一倍的Leu组,在试验第34天试验动物禁食2.5 h后血浆中的KIC浓度增加了45%。这与本研究结果一致。在本试验中,在饲喂后2 h,与对照组相比,试验Ⅲ组血浆中KIC浓度升高43.54%,在饲喂后4 h血浆中KIC的浓度升高了53.91%,这说明补喂Leu伊犁马血浆中的KIC浓度呈剂量依赖性增加。补喂Leu对伊犁马血浆HMB浓度差异不显著。饲粮中摄入的Leu可以绕过肝脏,使补喂后血浆Leu水平急剧升高,并在周围组织中激活Leu信号传导,随后Leu在肌肉中被支链氨基酸氨基转移酶的催化下发生可逆反应,生成KIC,然后KIC可以重新合成为Leu或被进一步氧化(Duan 等 ,2016;Shimomura 等 ,2001)。 大 多 数KIC氧化发生在肝脏中肝线粒体内,约90%的KIC被氧化为乙酰辅酶A的类似物异戊酰CoA,剩余5%~10%的KIC在肝脏细胞溶胶中被氧化为HMB,其中10%~40%随尿液排出,其余的转化为胆固醇和乙酰辅酶A(Arazi等,2018)。随后,仅由Leu产生的HMB被肾脏清除或代谢为β-羟基 -β - 甲 基 戊 二 酰 -CoA (Girón 等 ,2016)。Wilkinson等 (2013) 研究表明, 向年龄为 (21±0.3),体重相近的7名健康男性青年受试者提供含有3.42 g Leu的400 mL水,采集2.5 h内的血液样品,结果表明血浆中的HMB浓度一直保持不变,直到 150 min 后,HMB 浓度才从(3.2±0.6)μ mol适度上升到(10.3±3.5)μmol。 而在本研究中,补喂Leu对伊犁马血浆HMB浓度无显著影响,这可能是由于物种的不同和补喂Leu水平的不同引起的Leu代谢差异,也有可能是不同物种机体内Leu代谢途径中相关酶的活性表达不同。
KIC的半衰期极短,血液中KIC的代谢很快(Harper和 Benjamin,1984), 可能是因为 KIC 的氧化速率很快,这就解释了各组血浆中KIC的浓度在饲喂后1 h达到峰值,然后下降的原因。但随着时间的延长,试验Ⅲ组血浆KIC的浓度趋于平缓,可能是由于补喂的Leu水平足够,在KIC氧化的同时,Leu也在不断地转化为KIC(Moghei等,2016)。在Leu代谢的路径中由于只有极少的Leu才能转化为HMB,所以机体的HMB浓度也极少(Silva等,2017),这可能是本研究中补喂Leu对伊犁马血浆HMB浓度的影响不显著的原因。
4 结论
在本试验条件下,补喂亮氨酸能提高伊犁马血浆中亮氨酸的浓度,且随着补喂水平的增加而升高;补喂亮氨酸降低了血浆异亮氨酸浓度,血浆3-甲基组氨酸浓度有下降的趋势。补喂亮氨酸能提高伊犁马血浆中α-酮异己酸浓度,但对血浆β-羟基-β-甲基丁酸的浓度无显著影响。
