塔河油田奥陶系生物扰动碳酸盐岩储集层微观孔隙结构的数字化表征与连通性分析*
2020-08-10牛永斌徐资璐刘圣鑫钟建华赵佳如王培俊
牛永斌 徐资璐 刘圣鑫 钟建华 赵佳如 王培俊
1 河南理工大学资源环境学院,河南焦作 454003 2 中国地质科学院地质力学研究所,北京 100081 3 中国石油大学(华东)地球科学与技术学院,山东青岛 266580 4 中国石油塔里木油田分公司开发事业部,新疆库尔勒 841000
塔河油田是中国第1个古生界海相亿吨级大油田(韩革华等,2006)。传统上认为该奥陶系油藏是典型的“缝—洞型”油藏,溶洞、裂缝、小型溶蚀孔洞、礁滩粒间孔等是该油气藏的主要储集空间(顾家裕等,2001;翟晓先等,2002;邓小江等,2007;郑和荣等,2009;Lietal., 2016;Tianetal., 2016),基质对碳酸盐岩储集体的储集和渗透能力基本无贡献(鲁新便,2014)。然而,从目前开发实践看这种认识存在严重不足,因为有2个“简单”的科学事实无法回避: (1)塔河油田奥陶系碳酸盐岩油藏80%以上的井需要压裂才能求产;(2)有不少“功勋”井实际产出的石油量比缝洞系统本身体积要大得多,如S48井、T401井,至2011年11月,分别累计产油已超过150×104it和65×104it,这说明有的石油来自“缝—洞”系统之外。通过对塔河油田奥陶系鹰山组和一间房组岩心观察发现,泥晶灰岩中的白云石砂屑团块普遍含油,由此推测有相当大一部分油气来源于碳酸盐岩基质中的这些“白云石砂屑团块”。钟建华等(2010)、牛永斌等(2010,2018)认为,如果把塔河油田奥陶系这样的油藏比拟为一个“有机体”,“白云石砂屑团块”犹如这个“有机体”的细胞,缝合线则如毛细血管,(大)裂缝相当大动脉,而溶洞则是“心脏”,这样一个四级存储单元构成了塔河油田奥陶系油藏复杂的储集系统,其中“白云石砂屑团块”中的油气储集量不可忽视。具有同类储集层的加拿大阿尔伯特省Pine Creek油气田就是一个典型的例证(Baniaketal., 2013),该油田单井油气开发已超过40年,而塔河油田单井油气开发超过10年的寥寥无几,这也间接说明目前对塔河油田奥陶系储集层的认识还远远不够。目前普遍认为,这些“白云石砂屑团块”同“生物扰动”和“白云石化的不均匀”关系极为密切(贾振远和马淑媛,1984;郭建华等,1994;陈曦等,2011;郝毅等,2012;Maoetal., 2014;牛永斌等,2017,2018),因此,研究碳酸盐岩基质中“白云石砂屑团块”的发育特征、孔隙结构、储集能力和分布规律,对加深碳酸盐岩油气藏的勘探具有十分重要的意义,将成为开启碳酸盐岩型岩性油气藏的一把关键“钥匙”。
生物扰动(Bioturbation)为生物在生命活动过程中对生存环境周围的沉积物颗粒进行搅动、混合和破坏后所形成的各类生物潜穴、足迹和遗迹(牛永斌等,2019)。石油地质学家对生物扰动作用感兴趣,最初因为它可提供有关沉积环境的线索(Baniaketal., 2013;Abdel-Fattahetal., 2016;Bayet-Golletal., 2017;Rodríguez-Tovaretal., 2017;Zhengetal., 2018);随后又惊奇地发现它可降低或改善储集层的岩石物性,从而影响储集层质量、储量计算和流体流动特征(Gingrasetal., 2004a,2014;Pemberton and Gingras,2005;Knaust,2009,2014;Gordonetal., 2010;Tonkinetal., 2010;Bednarz and McIlroy,2012,2015;Greeneetal., 2012;La Croixetal., 2012,2013,2017;Qietal., 2012;Baniaketal., 2013;Ben-Awuah and Eswaran,2015;Hsiehetal., 2015,2017;Bayet-Golletal., 2017;Dey and Souvik,2017;Friesenetal., 2017;Golabetal., 2017a,2017b;Martiniusetal., 2017;Adametal., 2018;Quayeetal., 2019;Knaustetal., 2020)。生物扰动对碳酸盐岩组构和物性的改造尤为显著(Zenger,1992,1996;Gingrasetal., 2004a,2014;Pemberton and Gingras,2005;Cunninghametal., 2006;Rameil,2008;Knaust,2009,2014;Greeneetal., 2012;Jinetal., 2012;La Croixetal., 2012,2013,2017;Baniaketal., 2013,2014;Hsiehetal., 2015,2017;Friesenetal., 2017;Golabetal., 2017a,2017b;Adametal., 2018;Quayeetal., 2019;Knaustetal., 2020),碳酸盐岩基质沉积物由于原始物性较差,常常被认为很难形成有效的储集层(Rashidetal., 2015),但若生物扰动非常强烈,那么在适宜的温压条件、良好的时空物质匹配、充足的作用时间、适量的白云化流体供应和后期溶蚀作用下,多期次的生物潜穴叠加可形成横向连片、垂向连通的大规模生物扰动碳酸盐岩储集层(Zenger, 1992, 1996; Gingrasetal., 2004a, 2004b, 2005, 2014; Pemberton and Gingras, 2005; Rameil, 2008; Cunninghametal., 2009; Baniaketal., 2012,2013, 2014; Corlett and Jones, 2012; La Croixetal., 2013;Hsiehetal., 2017;Golabetal., 2017b;Martiniusetal., 2017;Adametal., 2018;Liuetal., 2019;Knaustetal., 2020)。而由于生物扰动常造成潜穴与围岩基质的组构差异,故这类储集层呈现极强的非均质性(李峰峰等,2019;沈瑛楚等,2019)。近年来,国外学者在生物扰动增强储集层储集性能的研究上取得了较为丰硕的成果(Gingrasetal., 2004a, 2004b, 2014;Pemberton and Gingras, 2005; Knaust, 2009, 2014; Tonkinetal., 2010; Bednarz and McIlroy, 2012, 2015; La Croixetal., 2012, 2013, 2017; Baniak, 2013; Baniaketal., 2014; Ben-Awuah and Eswaran, 2015; Bayet-Golletal., 2017; Dey and Souvik, 2017; Friesenetal., 2017; Quayeetal., 2019; Knaustetal., 2020), 而国内这方面的研究起步较晚,且主要关注在生物扰动对碎屑岩储集层的影响上(纪友亮等,1990;尹燕义等,1996;齐永安等,1998;林世国等,2012;Qietal., 2012)。对于受生物扰动更为强烈的碳酸盐岩,由于油气藏勘探起步较晚、前期关注重点不同等各种因素影响,致使相关研究成果较少。
生物扰动碳酸盐岩储集层的微观孔隙结构特征与分布规律决定了这种储层的储集能力、渗流能力以及渗流机理与渗流规律,微观孔隙结构的差异也将直接导致生产效果的差异(Gingrasetal., 2005,2014;Pemberton and Gingras,2005;Baniaketal., 2012,2014;La Croixetal., 2012,2013;Cunninghametal., 2009;Lilian,2016;Andreaetal., 2017;Golabetal., 2017a,2017b;Liuetal., 2019)。国内外众多这类储集层的开发实践表明,常见的注水“突窜”就主要与微观孔隙结构密切相关(Gingrasetal., 2005,2014;Pemberton and Gingras,2005;Cunninghametal., 2009;Lilian,2016;Golabetal., 2017a,2017b;陈明江等,2019)。因此,只有深化生物扰动碳酸盐岩储集层的微观孔隙结构研究,才能更准确地描述和分析微观孔隙结构中的流体分布形态及流动状态等(Raeinietal., 2019)。目前,能否正确认识这类储集层的微观孔隙结构特征和分布规律,是制约这类油气藏开发的关键问题,特别是微观孔隙结构定量表征和连通性分析是其中的难点问题(Hollisetal., 2010;Maoetal., 2014;Njiekaketal., 2018;Weietal., 2019),因为传统碳酸盐岩储集层孔隙结构研究方法对于这种微纳米孔隙尺度为主的储集空间分析具有一定局限性(王晨晨等,2013;赵新伟和许红,2016)。传统储集层孔隙结构研究方法主要包括 2类: 一类是依靠光学显微镜和电子显微镜来分析孔隙特征;另一类是压汞法和氮气吸附法,主要用于测试孔隙度等参数(赵新伟和许红,2016)。前者仅能观测到面的特征,后者则不够直观。X-CT射线层析扫描技术是近些年发展起来的一种利用X射线对岩石进行扫描成像、进而三维重构出孔隙空间展布特征的分析技术,其以快速、无损和三维可视化显示为特征,兼具传统方法的优点,同时又可弥补传统方法的不足,在生物扰动碳酸盐储层微观孔隙结构的直观刻画、定量表征和连通性分析方面具有明显优势(盛军等,2019)。
笔者在岩心观测和详细描述的基础上,利用偏光显微镜和阴极发光显微镜对优选样品的磨制薄片进行了观测和分析,详细记录了潜穴充填物和围岩基质的镜下特征,确定了生物潜穴充填的矿物成分和微观孔隙类型;然后利用X射线显微镜对精选的不同尺度柱塞样品进行了扫描与分析,对塔河油田奥陶系生物扰动碳酸盐岩储集层孔隙结构进行数字化表征与连通性分析。该成果真实、直观、准确刻画了生物扰动碳酸盐岩储集层微观孔隙结构特征,可为后续这类生物扰动碳酸盐岩储集层的微观渗流模拟研究提供所需的特征参数;未来将这类生物扰动碳酸盐岩储集层微观尺度上的孔隙结构数字化表征和渗流模拟结果与宏观分布规律相结合并应用到实际储集层评价中,可为这类储集层的油气资源量估算、开发方案优选、产能预测和提高采收率提供指导。
1 地质背景
塔河油田位于新疆塔里木盆地北缘、天山南麓的戈壁荒漠内,地处新疆维吾尔自治区的库车县和轮台县境内(图 1-a),面积为2400ikm2。根据油藏特征,可细分为12个小区(图 1-b)(Tianetal., 2016)。在早—中奥陶世(鹰山组和一间房组沉积期)的海侵、海退过程中,广泛发育的碳酸盐岩台地沉积了巨厚碳酸盐岩,厚度高达300im(图 1-c);同期适宜的生存环境使(古)生物非常繁荣,大量(古)生物在生命活动过程中在碳酸盐沉积物表面或内部建造了大量形态各异的潜穴,这些潜穴充填物在后期的成岩改造过程中(主要为白云化作用和溶蚀作用)形成了厚度极大的生物扰动碳酸盐岩储集体(牛永斌等,2018)。
2 孔隙结构的定量表征
碳酸盐岩储集层的孔隙结构主要为孔隙和喉道的几何形态、大小、分布、相互连通情况以及孔隙与喉道的配置关系,它们控制了储层流体的流动和分布,是储集岩的微观物理性质(邓虎成等,2014;孙亮等,2016;郑剑锋等,2016)。基于研究区奥陶系生物扰动发育的16口取心井的岩心观测,优选样品磨制127块岩石薄片,在河南省生物遗迹与成矿过程重点实验室利用偏光显微镜和阴极发光显微镜观测对这些岩石薄片进行观测和分析,详细记录潜穴充填物和围岩基质的镜下特征,确定生物潜穴充填的矿物成分和微观孔隙类型;然后精选生物扰动发育岩心样品,钻取不同尺度柱塞6个,在中国地质科学院地质力学研究所自然资源部古地磁与古构造重点实验室完成了X射线显微镜扫描与分析,完成塔河油田奥陶系生物扰动碳酸盐岩储集层孔隙结构的数字化表征,表征尺度集中在纳—微米级。
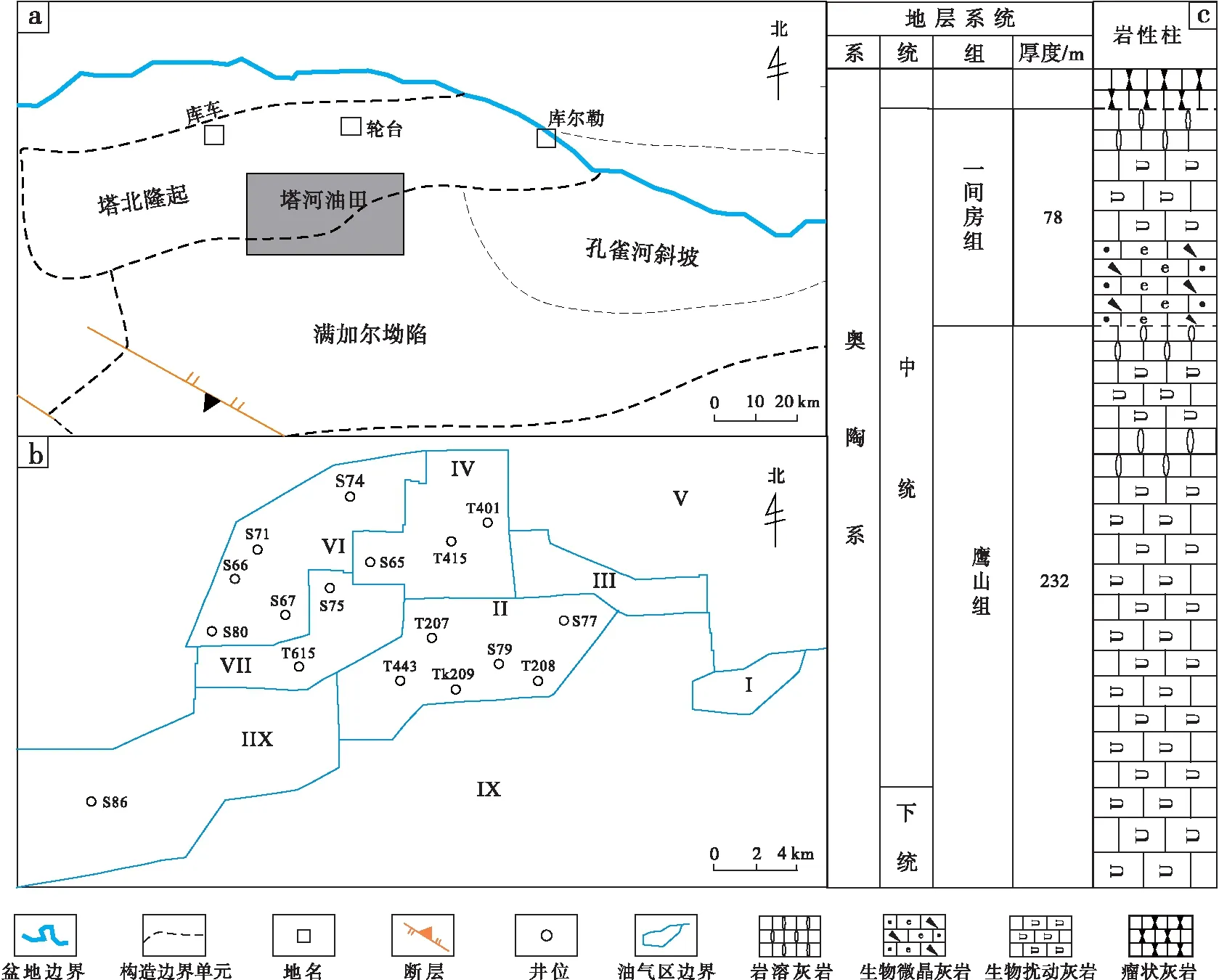
a—塔河油田构造位置;b—塔河油田区块划分;c—塔河油田中下奥陶统地层柱状图图 1 塔河油田地理位置、区域划分和中—下奥陶统地层柱状图(据牛永斌等,2018,有修改)Fig.1 Location,regionalism and the Lower-Middle Ordovician stratigraphic column of Tahe Oilfield(modified from Niu et al., 2018)
2.1 显微特征
岩心上生物扰动区域多为沥青质重油充填,呈暗褐色或灰黑色的不规则绸带状或离散的斑状;围岩基质很少被油斑浸染,颜色也相对较浅。生物扰动区边界微裂隙或缝合线常常比较发育,裂缝或缝合线内部也多被沥青充填,造成生物潜穴充填物与围岩基质岩石组构和颜色差异较大,非常容易识别。有时不同生物扰动区又常被裂缝切穿(图 2-a)。由油斑充填分布特征可知生物潜穴充填物孔隙较围岩基质明显发育。
通过偏光显微镜与阴极发光显微镜观察发现:生物潜穴充填物主要由白云石晶体组成,白云石晶体以自形、半自形晶型为主,他形白云石晶体较为少见,晶体之间普遍可见未白云化的残留灰泥;白云石晶体粒径普遍为100~200iμm,少量大于200iμm,以细晶结构为主,少见粉晶结构(图 2-b至2-e)。阴极发光显微镜下白云石颗粒呈亮红色的光,见明显的“亮边雾心”菱面体(图 2-c,2-e),表明这些白云石颗粒主要是在后期深埋藏过程中白云化作用形成的。有时生物潜穴充填物和围岩基质间可见清晰可辨的暗色“晕带”,推测为沥青浸染的边界。偏光显微镜下生物潜穴宿主围岩基质粒度非常细,多为微晶结构;阴极发光显微镜下它们发光极其微弱或不发光,揭示其矿物成分主要为微晶方解石,中间离散的亮蓝色斑点可能为极少量的钾长石(有可能为所含灰泥矿物蚀变残留的一部分)。生物潜穴充填物内部的白云石晶间孔发育,是这类储集层的主要储集空间,除此之外还发育生物铸模孔,但围岩基质储集空间不发育(图 2-c,2-e)。
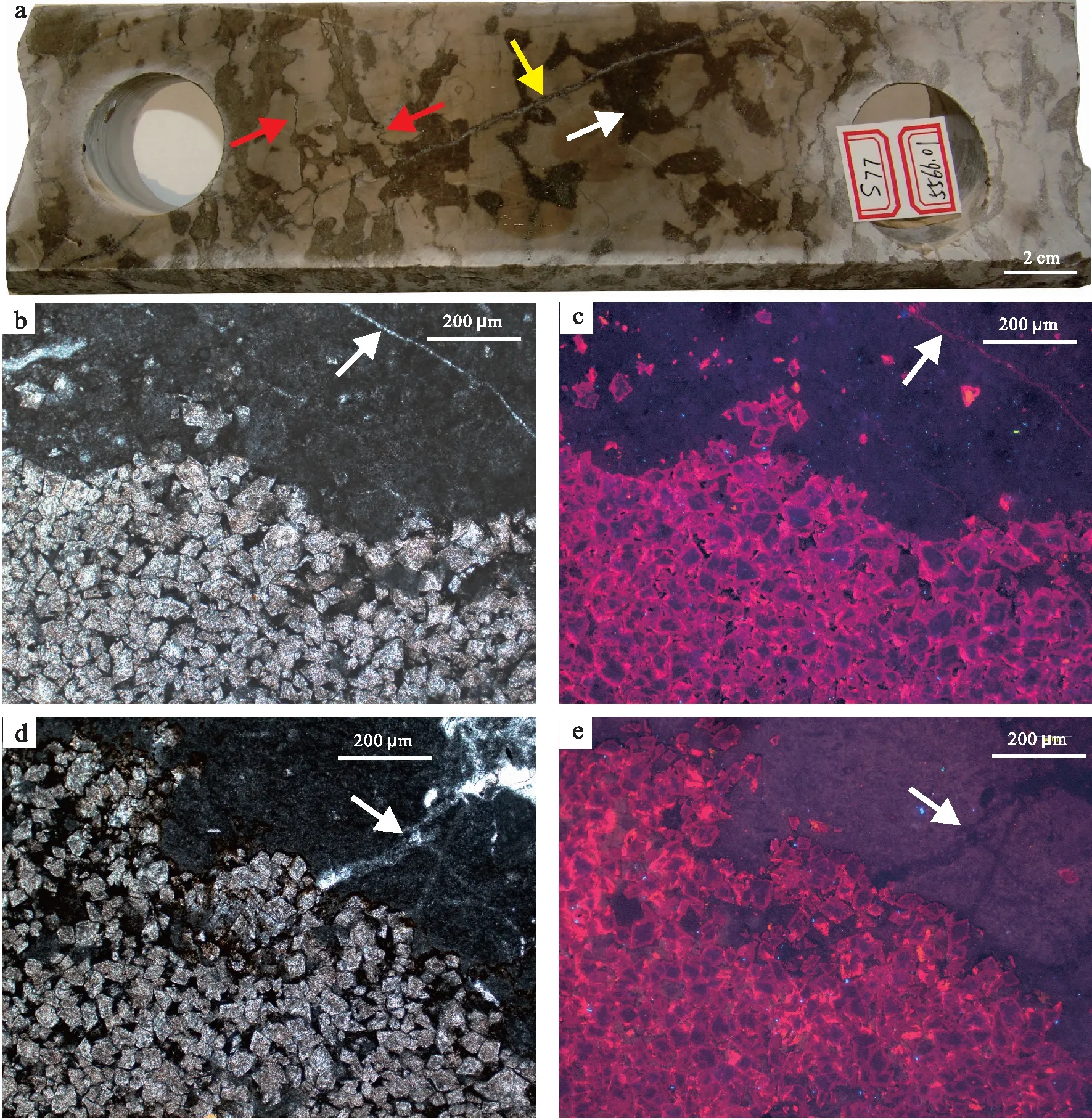
a—塔河油田,奥陶系,S77井,生物扰动含油岩心,岩性为生物扰动灰岩,白色箭头指示黑色斑块为重油浸染的遗迹潜穴(主要为Thalassinoides-like潜穴);中间黄色箭头指示为一被方解石半充填裂缝,红色箭头指示为缝合线或微裂缝;b—塔河油田,奥陶系,S77井,5578.93im,生物扰动灰岩薄片,下部为生物扰动区(主要为Thalassinoides-like潜穴),潜穴内充填物为白云石颗粒,晶间孔发育;上部为围岩基质发育区,主要为微晶方解石,右上角白色箭头所指为1条方解石充填裂缝;c—图b同视域的阴极发光显微镜照片,下部发红色的为生物扰动区域,上部为围岩基质中的亮蓝色的斑点为钾长石,右上角白色箭头所指为1条方解石充填的裂缝;d—塔河油田,奥陶系,S77井,5603.37im,生物扰动灰岩照片,左下部为生物潜穴,充填物为白云石颗粒,可见“亮心雾边”现象,内部可见沥青充填的微裂缝;右上部为围岩基质,主要为微晶方解石,内部白色箭头指示为1条方解石充填裂缝;e—图d同视域的阴极发光照片,左下部为发红色的为生物扰动区域,右上部发暗棕色或不发光的为围岩基质,矿物成分为微晶方解石,白色箭头指示为1条方解石充填裂缝,通过裂缝的 交切关系,可判定裂缝形成的先后次序图 2 塔河油田奥陶系典型岩心和薄片显微镜下生物扰动照片Fig.2 Bioturbation of the Ordivician in core from Tahe Oilfield and microphotographs under polarizing and cathodoluminescence microscope
2.2 定量表征
正如前文所述,光学显微镜观测范围有限,仅能观测某些面的孔隙结构特征,无法完整观测生物扰动碳酸盐岩储集层内部三维空间上的孔隙结构特征,故笔者重点利用X射线显微镜对所选取的生物扰动区域岩石柱塞样品进行扫描获取三维数据体。三维数据获取步骤如下:(1)通过对获取三维数据体的切片分析,进行孔隙发育区的图像分割;(2)通过对图像分割切片的三维重构,获取分析样品的三维孔隙结构栅格模型;(3)对重构后的三维孔隙结构栅格模型进行参数提取和建立球棍模型,实现生物扰动碳酸盐岩储集层孔隙结构的定量表征(图 3)。

图 3 塔河油田奥陶系典型岩心样品孔隙结构定量表征工作流程Fig.3 Quantitative characterization workflow of microcosmic pore structure of the typiacal Ordivician samples in core from Tahe Oilfield
笔者以生物扰动碳酸盐岩储集层非常发育的S77井(5567.03im)的1个岩石柱塞样(主体为潜穴充填物)为例说明定量表征过程。该样品在中国地质科学院地质力学研究所自然资源部古地磁与古构造重建重点实验室采用Carl Zeiss AG公司推出的X-microCT(Xradia 510 Versa)进行扫描,得到了985张高分辨率的岩石柱塞样品的连续二维CT切片,扫描的精度为0.8197iμm。由于扫描图像清晰度不一定为最佳效果,可能会影响后续孔隙分割,所以获取三维数据体(图 4-a)后首先对图像进行后期处理,使其在减弱原始数据图像噪点的同时较好地保留图像的细节特征及边缘信息(图 4-b); 继而再对处理过后的图像进行分割(即二值化处理)(图 4-c),三维重构二值化分割后所获的该岩样孔隙空间的栅格模型(图 4-d);最后从获取三维柱塞数据体中选取不同尺度更微观的数据体进行纳—微米尺度的孔隙结构分析。本次共选取了5种不同尺度的数据体进行微观孔隙结构分析与表征(表 1)。
通过对数字岩心孔隙结构中边长体素和孔隙度的关系分析发现,选取边长体素为450×450×450的立方体可作为分析样品的最优表征单元体REV(Representative Elementary Volume)。接着对选取的REV中分割的孔隙几何、孔隙网络模型(PNM)生成相应孔隙参数进行定量表征,如孔隙体积、表面积、孔的等效半径、喉道的等效半径、喉道的长度。最后利用所表征单元体导出的孔隙参数绘制各种孔隙/喉交汇图(图 5)。
通过分析表征单元体内孔隙结构特征可知: 孔隙体积集中于10~100iμm3,占总孔隙数量的近67%;孔隙表面积为10~100iμm2的数量最多,约占孔隙总数的59%;孔隙的等效半径集中在0~5iμm,约占总孔隙数量的80%;喉道长度则多集中在0~40iμm,约占喉道总数的91%,其中20~40iμm的最多,约占59%;喉道等效半径多在0~10iμm之间,约占喉道总数量的84%;孔喉等效半径分布具有偏态分布特征,主体分布在0~4iμm之间,其中孔隙等效半径以0~2iμm最多,喉道等效半径2~4iμm最多(图 5)。

a—岩心柱塞三维数据体;b—柱塞样品的三维切片;c—孔隙的阈值分割;d—三维重构的孔隙结构栅格模型图 4 塔河油田奥陶系典型岩心样品扫描三维数据体、Micro-CT图片和三维孔隙栅格模型Fig.4 Scan 3D data volume,micro-CT images and 3D pore raster model of the typiacal Ordivician samples in core from Tahe Oilfield
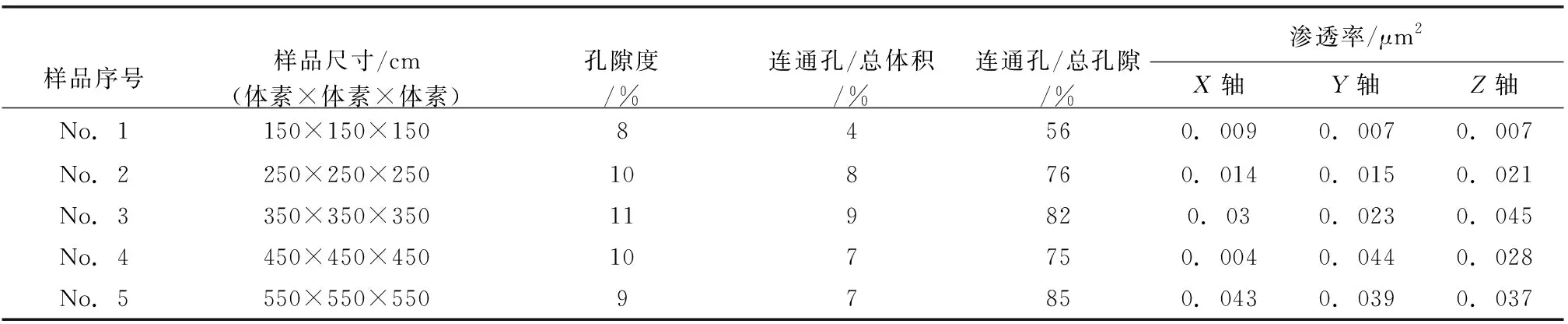
表1 塔河油田奥陶系典型岩心样品样品体素大小、孔隙度、连通孔占比和渗透率统计Table 1 Statisticas of pixel size,porosity,proportion of connected pore and permeability of the typical Ordivician samples in core from Tahe Oilfield
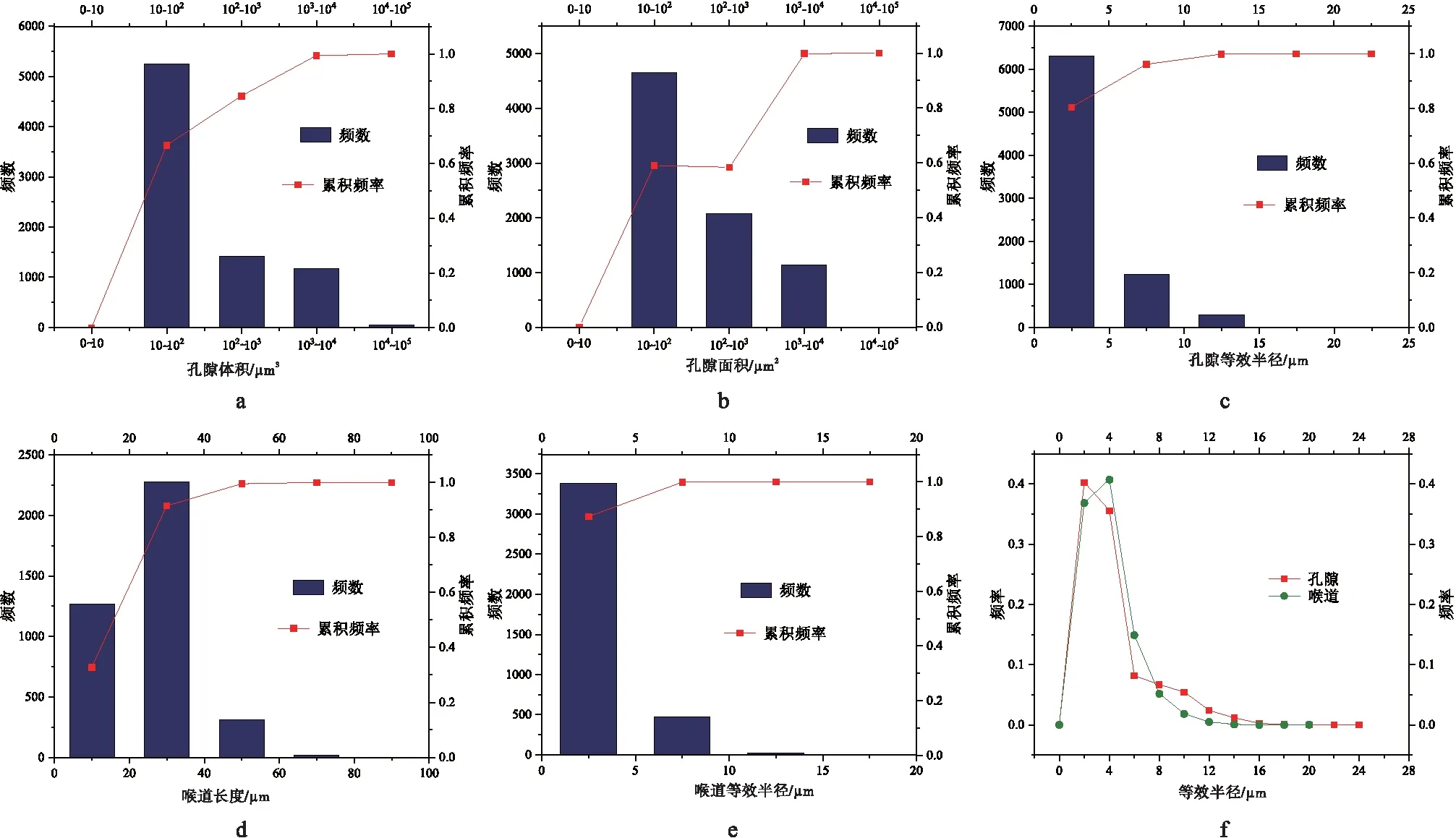
a—孔隙体积频数和累积频率直方图;b—孔隙面积频数和累积频率直方图;c—孔隙等效半径频数和累积频率直方图;d—喉道长度频数和累积频率直方图;e—喉道等效半径频数和累积频率直方图;f—孔隙和喉道频率折线图图 5 塔河油田奥陶系典型岩心样品孔隙/喉道的直方图和折线图Fig.5 Histograms and line graph of pores and throats of the typiacal Ordivician samples in core from Tahe Oilfield
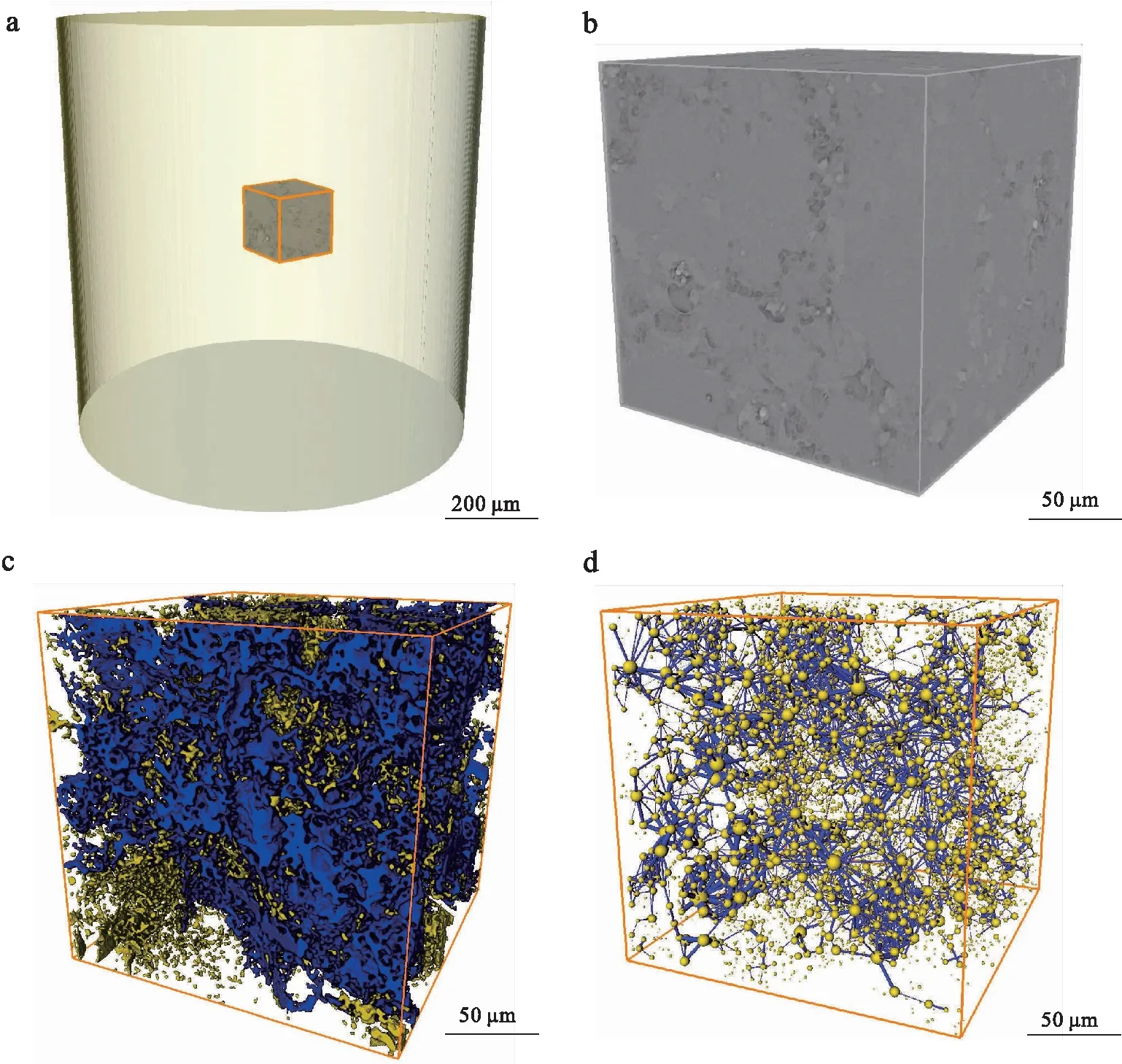
a—扫描3D数据体中的REV选取;b—REV的灰度模型;c—孔隙结构的三维栅格模型(蓝色部分为连通孔隙,黄色部分为不连通孔隙);d—孔隙网络结构的球棍模型图 6 塔河油田奥陶系典型岩心样品表征单元体与孔隙网络结构模型Fig.6 REV and pore structure network models of the typical Ordivician samples in core from Tahe Oilfield

图 7 塔河油田奥陶系No.5样品中孔隙的配位数频数直方图和累积频率折线图Fig.7 Coordination number frequency histogram and cumulative frequency line graph of the pores from No.5 sample of the Ordivician from Tahe Oilfield
3 孔隙的连通性分析
储集层连通性分析也是多孔储层介质中孔隙空间形态学研究的重要内容,不仅和有效储集空间有所关联,还与有效渗透率有直接相关;孔隙空间的连通性定量评价可为孔隙网络模型提供量化参数,并可应用到储层的渗透性分析中。储集空间连通性评价的一个重要手段就是借用数字岩心技术,该技术是描述岩石微观结构、建立数字模型、定量分析岩石孔隙结构特征、岩石物理数值模拟等技术的统称(孙亮等,2016)。
本次选取塔河油田奥陶系生物扰动型碳酸盐岩储集层连通性分析的岩样与前文孔隙结构定量表征的样品一致(图 6-a)。本次连通性分析实例仍然选取体素为450×450×450立方体为最优表征单元体REV(图 6-b)。从选取的表征单元体中提取孔隙并进行分割,得到该表征单元体REV孔隙结构的三维栅格模型(图 6-c),其中黄色部分为孤立的空隙,蓝色部分为连通孔隙。在表征单元体孔隙结构三维栅格模型的基础上,对所有孔隙利用分离算法进行进一步的分割,此过程通过计算表征单元体REV中各体素灰度梯度模量对孔隙进行分割,以此来确定孔隙的最佳轮廓。最后建立该样品的孔隙网格模型(PNM)(图 6-d),三维可视化地展示孔隙与吼道的空间分布特征和表征单元内各孔隙的连通性: 模型中黄色圆球表示的是被分割的每一个孔隙的等效球体,孔隙尺度大小按照球体的等效半径来定义,被分割出的孔隙等效半径越大,球就越大;蓝色棍棒表示连接孔隙的喉道,其大小也同样按照喉道的等效半径来定义,喉道的等效半径越大,蓝色棍棒就越粗。
根据 表 1 中各样品的孔隙度、渗透率和连通性状况分析可知,当选取的样品孔隙度在10%左右时,连通孔体积的占比在7%左右,连通孔隙约占总孔隙度的80%。其中No.1样品的连通孔隙度最大,连通孔隙度为9%,占总孔隙的82%;No.5样品的连通孔隙度为7%,占总孔隙的85%,在总孔隙的占比中最高。由该样品中的相邻孔隙配位数频数直方图和累积频率折线图可知: 配位数为0的孔隙占孔隙总数的79%,孔隙的最大配位数为21,在配位数1~21的孔隙之间,配位数1~4的孔隙约占孔隙总数的90%,其中配位数为2、3的孔隙最多(图 7)。此外,在该表征单元体REV中,绝大多数孔隙(79%)为孤立孔隙(配位数为0),但连通孔隙的体积在总孔隙的占比相对比较大。由此可知,虽然该表征样品整体的孔隙度不高(7%),但是其孔隙拓扑结构非常复杂,孔隙连通性很好。基于上述所选择表征样品的尺度限制,为获得柱塞岩样的连通状况,通过以2为倍数逐步放大表征单元体REV的边长体素进行尺度粗化升级分析,发现这类生物扰动型碳酸盐岩储层的孔隙局部连通性较好,而整体连通性相对较差。
4 研究意义
由于对塔河油田奥陶系生物扰动碳酸盐岩储集层微观孔隙结构的数字化表征与连通性分析是建立在对真实柱塞样品的高分辨率成像和无损检测的基础上, 因此建立的孔隙结构模型(PNM)真实反映了研究区这类储集层的微观性质, 能够准确、 直观地刻画生物扰动型碳酸盐岩储集层的微观孔隙结构特征。所获取的孔隙结构模型与孔喉的连通状况(孔隙与喉道的几何尺度及其拓扑关系等), 可为后续这类生物扰动碳酸盐岩储集层的微观渗流模拟研究提供所需的特征参数。而在本次所建立的孔隙网络模型上应用现代渗流力学理论进行微观尺度上多场作用下多相流体的渗流模拟, 可确定各种流体在这类储集层中的绝对渗透率和相对渗透率及其分布规律。若将这类生物扰动碳酸盐岩储集层微观尺度上的孔隙结构数字化表征和渗流模拟结果与宏观分布规律相结合并应用到实际储集层评价中, 可为将来这类生物扰动碳酸盐岩储集层的油气资源量估算、 开发方案优选、 产能预测和提高采收率提供指导。
5 结论
1)塔河油田奥陶系鹰山组和一间房组生物扰动碳酸盐岩储集层生物扰动区域多为沥青质重油浸染,生物潜穴充填物主要由白云石晶体组成,白云石晶体以自形、半自形为主,他形晶少见,白云石晶体间普遍可见残留灰泥成分。生物潜穴充填物内部白云石晶间孔隙发育,是这类储集层的主要储集空间,除此之外还有生物铸模孔;孔隙体积大多集中于10~100iμm3,喉道等效半径一般小于10iμm,喉道一般小于40iμm;围岩基质储集空间不发育。
2)塔河油田奥陶系鹰山组和一间房组生物扰动储集层孔隙结构模型揭示生物扰动充填物孔隙度介于7%~10%之间,孤立孔隙虽然数量占比较大,但连通孔隙体积在总孔隙体积中占比较大,且连通孔隙拓扑结构非常复杂,孔隙连通性很好;通过对表征单元体(REV)尺度粗化升级分析发现这类储集层的孔隙局部连通性较好,而整体连通性相对较差。
3)笔者建立的孔隙结构模型(PNM)真实反映了研究区生物扰动碳酸盐岩储集层的微观性质,可为后续这类生物扰动碳酸盐岩储集层的微观渗流模拟研究提供所需的特征参数。未来将这类生物扰动碳酸盐岩储集层微观尺度上的孔隙结构数字化表征和渗流模拟结果与宏观分布规律相结合并应用到实际储集层评价中,可为这类储集层的油气资源量估算、开发方案优选、产能预测和提高采收率提供指导。
