辣椒亲本间SRAP遗传距离与配合力、杂种优势的关系研究
2020-08-06吴立东
吴立东


摘 要:以11个辣椒亲本为材料按不完全双列杂交配置30个杂交组合,利用SRAP标记分析亲本间的遗传距离,探究其与配合力、杂种优势间的相关关系。结果表明:辣椒亲本间的遗传距离与亲本间的一般配合力(GCA)、组合间的特殊配合力(SCA)以及杂种优势均无显著的相关性,各性状的杂种优势与组合间SCA的相关性均不显著,只有果长的杂种优势与亲本间GCA呈极显著正相关。以上结果说明,利用遗传距离难以预测配合力以及后代的杂种优势,GCA和SCA也不能完全预测杂种优势。因此,遗传距离和配合力、杂种优势之间的关系还有待于进一步深入的研究。
关键词:辣椒;遗传距离;一般配合力;特殊配合力;杂种优势
中图分类号:S641.3 文献标识码:A
Abstract: The SRAP was used to analyze the correlation between the geneic distance with combining ability and heterosis of various agronomic traits in pepper. The materials consisted of 11 parent peppers and 30 F1 hybrids based on half diallel methods. The results showed that there was no significant correlation between the genetic distance and general combining ability (GCA), special combining ability (SCA) and heterosis of pepper parents. There was no significant correlation between SCA and heterosis of all traits, but only significant positive correlation between heterosis of fruit length and GCA. It is difficult to predict the combining ability and heterosis of the offsprings by the genetic distance, GCA and SCA could not be used to predict heterosis completely. Therefore, the relationship between the genetic distance, combining ability and heterosis needs to be further studied.
Keywords: pepper; genetic distance; general combining ability; specific combining ability; heterosis
DOI: 10.3969/j.issn.1000-2561.2020.04.005
辣椒原產于中南美洲热带地区的墨西哥、秘鲁等地,是人类种植最古老的作物之一,其栽培历史悠久,种质资源极其丰富[1]。不同辣椒品种或类型之间存在着明显的杂种优势[2-3],利用杂种优势是进行辣椒新品种的选育的主要途径之一;20世纪60年代江苏省农业科学院蔬菜所育成我国第1个辣椒杂交品种‘早丰一号,产量比一般品种增产30%~40%[4]。到80年代以后,我国辣椒育种迅速发展,杂种优势利用效率得到大幅度的提高,选育出大量早熟优质高产的辣椒杂交种在生产上大面积应用推广,获得巨大经济效益,如中椒、湘研、茂椒和苏椒等一系列辣椒新品种。
在品种选育过程中,选择优良亲本配制强优势杂交组合是杂种优势育种的关键。目前,主要通过配合力测定和遗传距离分析来进行亲本的选配和杂种优势的预测。国内外学者已经开展了大量的关于配合力和杂种优势方面的研究工作,发现配合力可以预测杂种优势[5-7],杂种优势受亲本一般配合力和杂交组合间特殊配合力的影响[8-10]。近年来,随着分子生物学的迅速发展,DNA分子标记技术被广泛应用于作物的遗传距离和杂种优势的相关研究[11-14],但研究结果不尽相同。有的研究发现,遗传距离与杂种优势间存在不同程度的相关性,可以进行杂种优势预测,如水稻[11]、甘蓝[12]等;也有研究显示,遗传距离与杂种优势之间的相关性较弱或不存在相关性,如玉米[13]、小麦等[14]。因此,进一步深入研究遗传距离与配合力、杂种优势的相关关系,可为今后杂种优势有效利用提供科学依据。
本研究选取11个辣椒亲本材料,按5×6不完全双列进行杂交,利用SRAP标记测定亲本间的遗传距离,分析辣椒亲本间遗传距离和配合力、杂种优势的相关性,以期为亲本选配和杂种优势预测提供有效的理论依据。
1 材料与方法
1.1 材料
供试材料母本(P1)为m35、m118、m62、m254、m119,父本(P2)为f65、f15、f93、f243、f65、f188,均为经过多代自交分离的纯合自交系材料,所有材料均由三明市农业科学研究院蔬菜所提供。
1.2 方法
1.2.1 试验设计 2018春季(4—7月)上述亲本材料按5×6 NCⅡ配制30个杂交组合;2018年秋季(8—11月),将11个亲本材料和30个杂交组合种植于沙县西霞村蔬菜育种基地。试验采用随机区组设计,重复3次,每小区种植株距40 cm,行距50 cm,双行种植,共种植30株,田间管理同一般大田生产管理。
1.2.2 數据收集 在辣椒生育期内,每小区随机选定10株,参照农业行业标准《植物新品种特异性、一致性和稳定性测试指南 辣椒》(NY/T 2234-2012)[15]定期考查亲本及其杂种F1的始花节位、果长、果宽、果柄长、果肉厚、单果重、结果数和单株产量等表型性状。
1.2.3 SRAP标记 采用改良的CTAB法提取全基因组DNA,用1%琼脂糖凝胶电泳检测DNA质量。根据Li等[16]公布的SRAP引物(包括13条上游引物和9条下游引物),共合成117对引物组合,从中筛选出多态性好、条带清晰的22对引物组合用于遗传距离的分析。PCR扩增反应体系(20 ?L)为:模板DNA 50 ng,10 ?mol/L dNTPs 0.4 ?L,Taq聚合酶0.2 U,上下游引物各50 ng,10×buffer 2 ?L,用ddH2O补充至20 ?L。PCR扩增反应程序为:94 ℃预变性5 min;94 ℃变性1 min,35 ℃复性1 min,72 ℃延伸1 min,5个循环;94 ℃变性1 min,50 ℃复性1 min,72 ℃延伸1 min,35个循环;72 ℃延伸10 min,4 ℃保存备用。采用6%聚丙烯酰胺凝胶进行电泳,用银染法检测。
1.3 数据处理
采用Excel 2007软件对田间数据进行统计分析,中亲杂种优势按公式:H=[F1(P1+P2)/2]/[(P1+ P2)/2]×100%。一般配合力(general combining ability, GCA)、特殊配合力(special combining ability, SCA)采用DPS 7.05软件进行分析。根据SRAP引物在11个亲本材料中PCR扩增的结果,在相同迁移位置上有带的记为“1”,无带的记为“0”,并利用NTSYS-pc软件计算遗传距离(GD)。采用SPSS 19.0软件对各组合的亲本遗传距离、配合力的效应值和杂种优势之间进行相关性分析。
2 结果与分析
2.1 SRAP遗传距离分析
本研究选取条带清晰且多态性好的28对SRAP引物对11个辣椒亲本材料进行PCR扩增,共扩增出189条带,每对引物扩增出的4~9条不等,其中152条具有多态性,多态性比率为80.4%。利用SRAP标记对11个辣椒亲本材料进行遗传距离计算(表2),结果发现遗传距离变异幅度为0.2149~ 0.5605,平均遗传距离为0.3130,其中A5×B3的遗传距离最大(0.5605),A5×B6的遗传距离最小(0.2149)。统计结果表明,30个遗传距离中,其中有13个遗传距离大于平均距离(0.3130),表明这11个亲本材料间的遗传差异较大。
2.2 杂种优势表现
由表3可看出,辣椒30个杂交组合间各性状的杂种优势差异较大,其中单果重的杂种优势变异幅度最大,为36.31%~86.06%。A3×B5组合在果长上杂种优势最高,为58.49%。辣椒各个组合在始花节位上的杂种优势均为负值,表现为负向优势,其中A3×B2的杂种优势最大,为1.60%。A2×B3组合在果宽上的杂种优势最高,为38.75%,但在果柄长上杂种优势最小,为31.81%。A1×B2组合在果柄长上杂种优势最高,为47.92%。A1× B1组合在果肉厚上杂种优势上最大,但在结果数上最小,为47.19%。A2×B3组合在单果重和单株产量上杂种优势最高,分别为86.06%和85.90%。
2.3 配合力分析
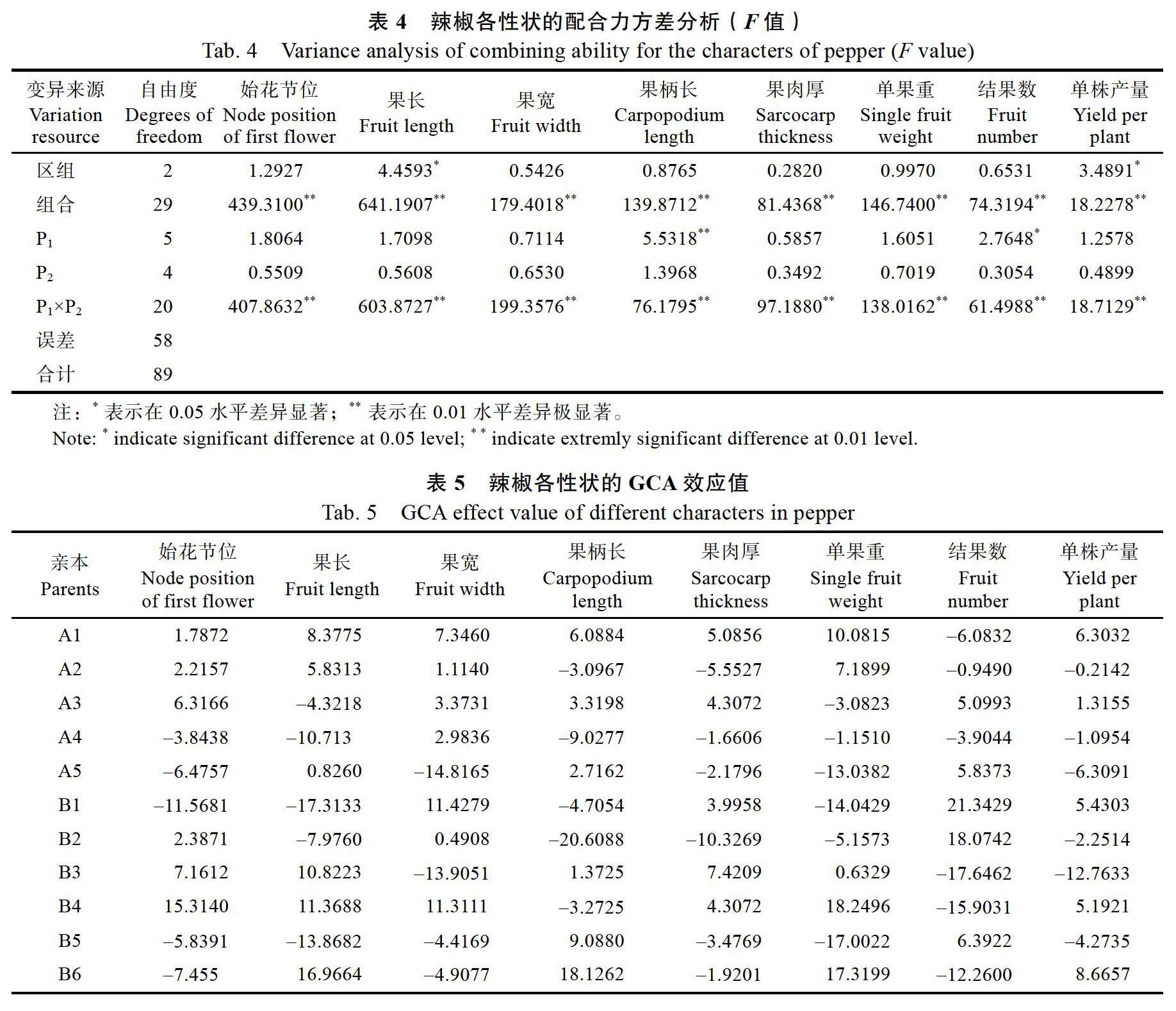
辣椒30个组合8个性状的配合力方差分析结果见表4。由表4可见,区组间除果长和单株产量差异显著外,其余性状的区组间均无显著差异,而组合间的各个性状均达到极显著差异水平,表明11个亲本材料间存在显著的遗传差异,可进一步做GCA和SCA分析。
对亲本间的GCA以及杂交组合间的SCA方差分析表明,P1中果柄长和结果数的GCA达到了极显著水平,P2中各性状的GCA均未达到显著水平,而P1×P2的SCA方差中各性状均达到了极显著水平。由此可见,果柄长和结果数主要受P1的一般配合力和组合间的特殊配合力作用,其遗传主要受加性效应和非加性效应共同影响的;而始花节位、果长、果宽、果肉厚、单果重和单株产量主要受组合间的特殊配合力作用,其遗传主要受非加性效应影响较大。
11个辣椒亲本的GCA效应值见表5,在母本中,A3在始花节位上的GCA效应值最高,A2次之;A1在果长、果宽、果柄长、果肉厚、单果重以及单株产量上的GCA效应值最高;A5在结果数上的GCA效应值最高,A3次之。在父本中,B4在始花节位和单果重上的GCA效应值最高;B6在果长、果柄长和单株产量上的GCA效应值最高;B1在果宽和结果数上的GCA效应值最高;B3在果肉厚上的GCA效应值最高,B4次之。
从表6可看出,11个辣椒亲本材料组配的30个杂交组合间的SCA差异较大,且同一亲本所杂交的不同组合间的SCA效应值差异也较大。始花节位中,以A4×B3、A3×B4、A5×B2的SCA为较高;果长以A2×B4、A1×B6、A4×B2以及A3×B5的SCA为较好;果宽以A1×B1、A3×B6、A3×B3的SCA为较高;果柄长以A3×B4的SCA为最高,A3×B5的SCA为最低;果肉厚以A1×B1的SCA为最高,A2×B3的SCA为最低;单果重以A2×B2和A1×B1的SCA为较高;结果数以A3×B1的SCA为最高;单株产量以A4×B2、A5×B4、A3×B1的SCA为较高。
结合表5、表6分析可见,在某些杂交组合中,亲本的GCA效应值较高,用其配制的杂交组合的SCA效应值也较高,如A3和B4在始花节位上的GCA效应值均较高,用它们配制的杂交组合A3×B4的SCA效应值也较高。但有些组合中,亲本具有较高的GCA效应值,所配制杂交组合的SCA不一定高,例如A1和B6在果柄长上的GCA效应值均较高,且都为正值,但其杂交组合A1×B6的SCA效应值为负值。
從表7可看出,始花节位、果长、果柄长、单果重、结果数以及单株产量等性状的P1的GCA基因型方差大于P2,表明这些性状受母本影响较大;辣椒各性状组合间的SCA基因型方差较大,表明这些性状受SCA影响较大。始花节位、果长、果宽、果肉厚、单果重、结果数以及单株产量等性状的SCA方差占总方差的比例较大,表明这些性状主要受SCA影响。辣椒8个性状的广义遗传力均达到了85%以上,表明各个性状的表型变异主要受基因型控制,受环境影响较小,遗传力较高,可以对这些性状进行早期选择。辣椒各性状的广义遗传力与狭义遗传力相差较大,表明这些性状主要受非加性基因控制,受环境影响较大。
2.4 遗传距离与配合力、杂种优势的相关性
本研究对辣椒各性状杂种优势、配合力与亲本间的遗传距离进行相关性分析,结果见表8,辣椒亲本间的遗传距离与GCA、SCA以及杂种优势均无显著的相关关系,且相关程度均较弱,说明利用SRAP分子标记估算的亲本间遗传距离难以预测到配合力以及后代的杂种优势。在辣椒8个性状中,只有果长的杂种优势与GCA呈极显著正相关关系,相关系数为0.765,其余7个性状的杂种优势与GCA的相关性均不显著,而始花节位的杂种优势与GCA呈负相关关系。辣椒各性状的杂种优势与SCA的相关性均不显著,且相关程度均较弱,其中单果重的杂种优势与SCA呈负相关关系。
3 讨论
关于辣椒的杂种优势,前人已做了大量研究。段晓铨等[17]研究表明,辣椒F1代中初花节位、果长、果宽、果肉厚、单株挂果数、单果重量以及单株产量的超中优势平均值均表现为正向优势,其中以单株产量的杂种优势最为明显。何建文等[18]也得出了类似的结果,不同的是,果肉厚的超中优势平均值表现为负向优势。而本研究发现,辣椒的始花节位、果柄长和果肉厚的杂种优势平均值表现为负向优势,其余5个性状均为正向优势,其中单株产量的杂种优势最为明显。
杂种优势是植物界中普遍存在的一种现象,2个亲本间存在的遗传差异是产生杂种优势的基础,人们通常用配合力测定、遗传距离等方法进行杂种优势预测。在遗传距离方面,国内外学者已做了大量关于利用分子标记技术预测杂种优势的研究,但其结果却不尽相同[19-21]。赵庆勇等[19]研究认为,分子标记可用于水稻杂种优势群的划分和遗传多样性分析,但相关程度还不足以预测产量杂种优势,而姚艳梅等[12]研究显示,利用基于SRAP标记的遗传距离有可能用于预测油菜的杂种优势。本研究结果表明,SRAP分子标记遗传距离与辣椒各性状的杂种优势之间的相关系数都较小,难以用来准确预测杂种优势;同时,SRAP分子标记遗传距离与亲本的GCA、组合间的SCA之间的相关程度都较弱,说明遗传距离对亲本间组配的指导作用不大。王玉平等[22]、张晓梅等[23]也得出了类似的结果,表明遗传距离和配合力之间的关系还有待于进一步的研究。在配合力方面,大多数研究者认为只有亲本的GCA高,利用该亲本组配的SCA高的杂交组合,才可能获得较强的杂种优势[24-27],这与本研究的结果不一致。如亲本材料A1、A2、B4、B6在果长上的GCA均较高,其配制的杂交组合A1×B6、A2×B4的SCA也都很高,但它们的杂种优势均不明显,这在其他性状上也得到了类似的结果。因此,要想获得强优势组合,并不能仅仅依据亲本的GCA、杂交组合的SCA效应值的高低来决定,还要结合辣椒各性状的综合表现进行选择。此外,本研究在研究配合力和杂种优势的相关性时发现,杂种优势仅与果长的GCA呈极显著正相关,与其他性状的GCA、SCA均有一定的相关性,但不显著,说明GCA、SCA只能在一定程度上对杂种优势有指导作用,但不能完全预测杂种优势。
参考文献
[1] 邹学校. 中国辣椒[M]. 北京: 中国农业出版社, 2002.
[2] 黄国伟, 苏晓华, 黄秦军. 植物杂种优势机理研究进展[J]. 世界林业研究, 2012, 25(1): 13-18.
[3] 袁俊水. 辣椒杂种优势利用途径分析[J]. 辣椒杂志, 2009, 7(4): 24-25.
[4] 陈桂英, 刘汉宇, 李尽朝. 辣椒杂种优势利用的研究进展[J]. 内蒙古农业科技, 2003(6): 30-32.
[5] Ahangar L, Ranjbar G A, Nouroozi M. Estimation of combining ability for yield and yield components in rice (Oryza sative L.) cultivars using diallel cross[J]. Pakistan Journal of Biological Sciences, 2008, 11(9): 1278-1281.
[6] Gopal J, Kumar V, Luthra S K. Top-cross vs. poly-cross as alternative to test-cross for estimating the general combining ability in potato[J]. Plant Breeding, 2008, 127(5): 441-445.
[7] Huang M, Chen L Y, Chen Z Q. Diallel analysis of combining ability and heterosis for yield and yield components in rice by using positive loci[J]. Euphytica, 2015, 205(1): 37-50.
[8] 赵庆勇, 朱 镇, 张亚东, 等. 杂交粳稻品质性状的配合力和杂种优势分析[J]. 江苏农业学报, 2008, 24(4): 387-393.
[9] 陈发波, 龙永昌, 姚启伦, 等. 茎瘤芥(榨菜)主要性状配合力及杂种优势分析[J]. 河南农业科学, 2017, 46(1): 93-99.
[10] 杨鹏鸣, 蔡祖国, 李新峥, 等. 南瓜自交系主要农艺性状配合力及杂种优势研究[J]. 湖北农业科学, 2006, 45(4): 481-483
[11] 蔡 健, 兰 伟. AFLP标记与水稻杂种产量及产量杂种优势的预测[J]. 中国农学通报, 2005, 2l(4): 39-43, 121.
[12] 姚艳梅, 聂 平, 杜德志. 不同生态类型甘蓝型油菜的SRAP分析及其遗传距离与杂种优势的关系[J]. 西北农业学报, 2012, 21(1): 80-87.
[13] 谭 静, 番兴明, 杨峻芸, 等. 玉米分子标记遗传距离与产量杂种优势关系的研究[J]. 西南农业学报, 2004, 17(3): 278-281.
[14] Dreisigacker S, Melchinger A E, Zhang P, et al. Hybrid performance and heterosis in spring bread wheat, and their relations to SSR-based genetic distances and coefficients of parentage[J]. Euphytica, 2005, 144(1-2): 51-59.
[15] 全国植物新品种测试标准化技术委员会. 植物新品种特异性、一致性和稳定性测试指南 辣椒: NY/T2234-2012 [S]. 北京: 中国农业出版社, 2012.
[16] Li G, Quiros C F. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica[J]. Theoretical and Applied Genetics, 2001, 103(2-3): 455-461.
[17] 段曉铨, 周书栋, 马艳青. 辣椒杂交F1代主要农艺性状杂种优势分析[J]. 湖南农业科学, 2013(19): 7-10.
[18] 何建文, 姜 虹, 赖 卫, 等. 辣椒杂交F1代主要性状杂种优势分析[J]. 长江蔬菜, 2012(14): 19-22.
[19] 赵庆勇, 朱 镇, 张亚东, 等. SSR标记遗传距离与粳稻杂种优势的相关性分析[J]. 中国水稻科学, 2009, 23(2): 141-147.
[20] Reif J C, Melchinger A E, Xia X C, et al. Use of SSRs for establishing heterotic groups in subtropical maize[J]. Theoretical and Applied Genetics, 2003, 107(5): 947-957.
[21] Lanza L L B, Souza C L, Ottoboni L M M, et al. Genetic distance of inbred lines and prediction of maize single-cross performance using RAPD markers[J]. Theoretical and Applied Genetics, 1997, 94(8): 1023-1030.
[22] 王玉平, 郭明星, 王 峰, 等. SSR标记遗传距离与杂交稻农艺性状一般配合力的相关性分析[J]. 分子植物育种, 2013, 11(3): 317-325.
[23] 张晓梅, 苗增建, 杜德志. 新型早熟甘蓝型春油菜恢复系遗传距离及遗传成分与配合力关系[J]. 西北农业学报, 2013, 22(1): 68-72.
[24] Makanda I, Tongoona P, Derera J. Combining ability and heterosis of sorghum germplasm for stem sugar traits under off-season conditions in tropical lowland environ-ments[J]. Field Crops Research, 2009, 114(2): 272-279.
[25] 刘金波, 宋兆强, 王宝祥, 等. 几个新选杂交水稻不育系和恢复系的主要农艺性状的配合力分析[J]. 西南农业学报, 2016, 29(2): 209-213.
[26] 王利民, 张建平, 党 照, 等. 胡麻两系杂交亲本的配合力及杂种优势分析[J]. 中国农业科学, 2016, 49(6): 1047-1059.
[27] 陈海荣, 王加红, 韦祝山, 等. 辣椒的新品种保护和DUS测试[J]. 辣椒杂志, 2006(4): 15-18.
