春季南黄海、东海浮游植物群落结构与环境因子
2020-08-05郭术津姚庆祯魏传杰孙晓霞
郭术津姚庆祯魏传杰孙晓霞
(1.中国科学院 海洋研究所胶州湾海洋生态系统国家野外科学观测研究站,山东 青岛266071;2.中国科学院 海洋研究所海洋生态与环境科学重点实验室,山东 青岛266071;3.中国科学院 海洋大科学研究中心,山东 青岛266071;4.海洋化学理论与工程技术教育部重点实验室,山东 青岛266100;5.中国海洋大学 化学化工学院,山东 青岛266100;6.中国科学院 海洋研究所海洋环境工程中心,山东 青岛266071;7.中国科学院大学,北京100049)
浮游植物是海洋中的初级生产者,通过光合作用将CO2转化为有机碳,启动海洋中的物质循环和能量流动,并支持渔业生产[1]。另外,浮游植物通过“生物泵”将碳向海底运输,形成碳汇,参与调节大气CO2浓度,进而影响全球气候[2]。
陆架边缘海仅占全球海洋总面积的7.6%,却提供全球生物总生产量的26%和世界渔业总产量的67%~75%[1]。另外,陆架边缘海是一个复杂的活跃体系,生产力高,在碳的海洋生物地球化学循环中的作用也十分重要。因此研究陆架边缘海的浮游植物群落结构,有利于我们深入认识和了解陆架边缘海的生物资源状况及其碳循环机制。
黄、东海总面积约1.23×106km2,70%为陆架区,水深一般不超过200 m,是中国重要的陆架边缘海[3]。早在20世纪50年代中国研究人员便开展了黄、东海浮游植物的相关调查研究[4-9]。早期研究以网采方法为主,不能获得浮游植物在各水层的垂直分布信息;后来以水采方法为主的调查研究也相继展开[10-13],并积累了大量的宝贵资料,不过多数研究限于黄海或东海海域,将黄、东海作为一个整体区域进行调查研究的报道较少。
为研究目前黄、东海海域浮游植物的群落现状及与环境因子的关系,我们使用水采的方法分析了2017年春季南黄海、东海海域的浮游植物群落;同时,对调查海域的温度、盐度和营养盐等理化参数也进行测定,对环境数据和浮游植物群落数据进行相关性分析。希望为南黄海、东海浮游植物相关的生态学研究提供借鉴和参考。
1 材料和方法
1.1 研究区域
搭乘“科学三号”调查船于2017年春季(2017-04-20—05-09)在南黄海和东海海域(119°30′~125°00′E,26°00′~36°00′N)设置的67个站位进行水文、化学和生物的综合调查(具体研究区域和站位设置见图1,虚线为水深)。从北向南分别选择A,B,C,D,E和F六条断面,来展示调查区域理化因子和浮游植物的垂直分布趋势。另外,鉴于黄、东海生态系统的差异较大,为将调查区2个海域的浮游植物群落特征区别分析,根据黄、东海的分界线以长江口北侧启东角与济州岛之间连线为分界线的准则,将站位3200-03和3200-05以北的区域划为南黄海区域,2个站位及其以南的区域划为东海区域。
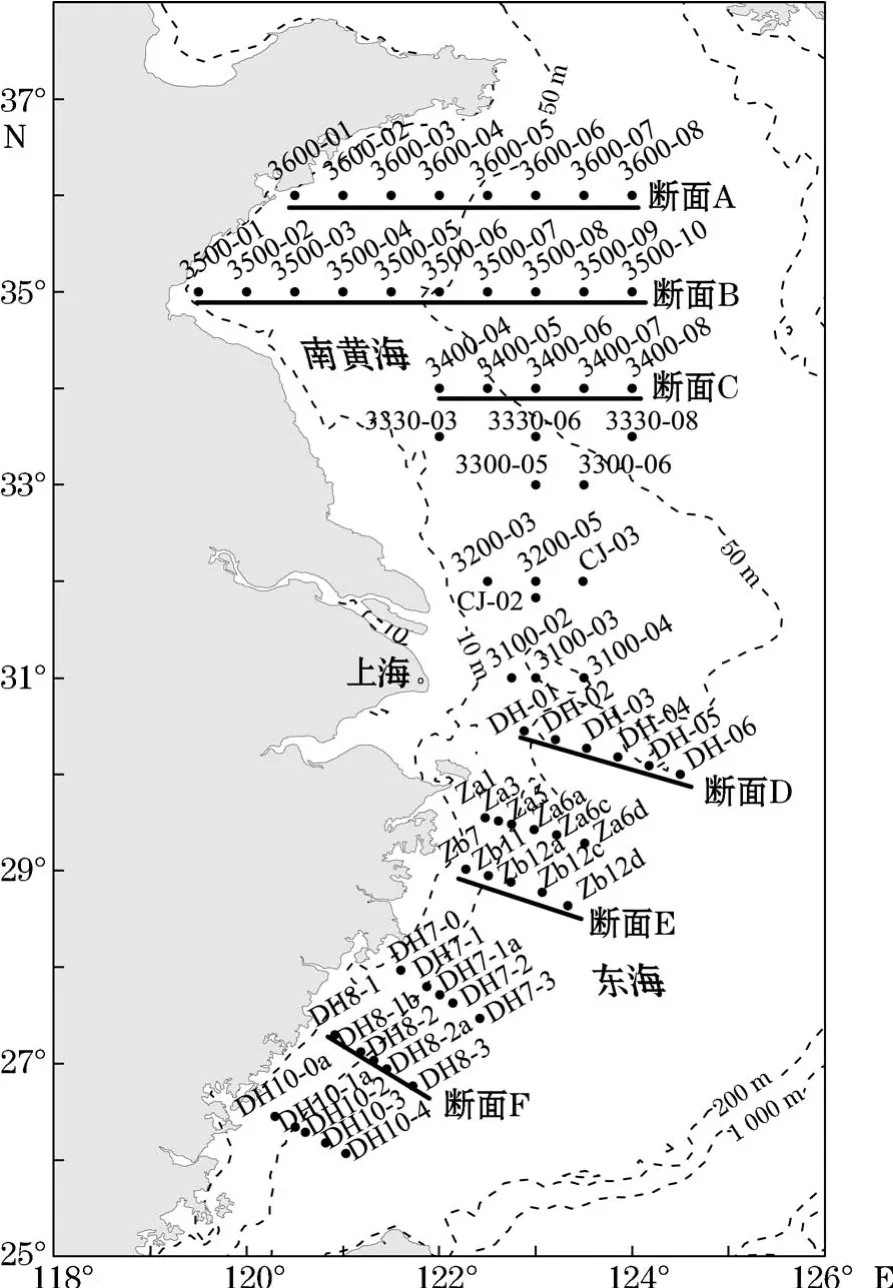
图1 2017年春季南黄海、东海调查区域和站位 ()Fig.1 Sampling stations()in the southern Yellow Sea and East China Sea in spring 2017
1.2 样品采集及分析
现场使用便携式美国Seabird 917Plus CTD的Rosette采水器,在不同水层采集水样,CTD同时测量温度、盐度和深度等参数。采水深度的设置为水深0,10,20,30,50 m和底层。取500 m L水样置于大口PE样品瓶中,加甲醛溶液固定(终浓度体积分数1%)。样品遮光置于阴凉处保存。航次结束后2个月内,在陆上中国科学院海洋研究所胶州湾海洋生态系统国家野外科学观测研究站实验室完成对浮游植物样品的分析工作。实验室内浮游植物样品按照Utermöhl方法进行分析[14]。取25 m L浮游植物亚样品置于Hydro-bios的Utermöhl计数框内,静置沉降24 h后,用日本Olympus CX-31倒置显微镜在200倍或400倍下进行物种鉴定和细胞计数。
营养盐样品采集后立即用浓度为1 mol/L的HCl溶液处理过的0.45μm醋酸纤维素膜过滤,于-20℃冷冻保存,用于测定硝酸盐(NO3-N)、亚硝酸盐(NO2-N)、磷酸盐(PO4-P)、硅酸盐(SiO3-Si)和铵盐(NH4-N)。营养盐的室内分析采用德国Bran-Luebbe公司生产的Quaatro营养盐自动分析仪完成,各营养盐的检出限分别为NO3-N浓度0.015μmol/L,NO2-N浓度0.003μmol/L,PO4-P浓度0.024μmol/L,SiO3-Si浓度0.030μmol/L,NH4-N浓度0.04μmol/L。
1.3 数据分析
对调查区浮游植物群落结构的香侬-威纳多样性指数(Shannon-Wiener diversity index,H′)、Pielou均匀度指数(Pielou′s evenness index,J)和各物种的优势度指数(Y)进行了计算[11-12]:

式中Pi是样品中第i种的细胞丰度与该样品总细胞丰度的比值,S为该样品总物种数。

式中S为样品物种数。

式中ni是第i个物种在所有样品中的细胞丰度之和,N为所有样品的细胞丰度之和,fi为该物种在调查区各站位出现的频率。
浮游植物细胞丰度与环境因子的关系采用Pearson相关性分析,浮游植物细胞丰度数据与环境参数经过lg(x+1)转换后,应用SPSS14.0软件[15]计算其相关系数以及显著性水平。
2 结 果
2.1 海区环境特征
调查海域表层温度、盐度和营养盐浓度分布见图2。调查区表层温度为10.64~24.39℃,平均值为16.36℃;表层盐度为24.23~34.56,平均值为31.4,盐度最低值出现在长江口邻近的3100-03站位(图2b)。NO3-N浓度为0.08~24.51μmol/L,平均值为5.70μmol/L,高值区集中在长江口邻近的外侧站位和闽浙沿岸区域(图2c)。PO4-P浓度为0.03~0.80μmol/L,平均值为0.14μmol/L,高值区集中在闽浙沿岸区域(图2d)。SiO3-Si浓度为0.30~25.19μmol/L,平均值为6.12μmol/L,高值区集中在长江口外侧及闽浙沿岸区域,另外南黄海中南部也有高值区存在(图2e)。
调查区断面温度、盐度和硝酸盐浓度分布(图3)显示,在南黄海海域温度从表层到底层基本呈逐渐降低趋势,在断面A中部的3600-04站位中下层有冷舌现象;盐度则从近岸到外海呈逐渐升高趋势。断面A和B的营养盐分布趋势表明南黄海的近岸站位NO3-N浓度较高;南黄海中部的10~30 m水层存在NO3-N的涌升现象。在东海海域,除了断面D温度呈现从近岸到外海逐渐降低的趋势,断面E和F的温度均从近岸到外海呈逐渐升高的趋势。3条断面盐度均呈从近岸到外海、从表层到底层逐渐升高的趋势。东海海域的3条断面NO3-N浓度高值均出现在近岸区域,且从近岸到外海呈逐渐降低趋势。
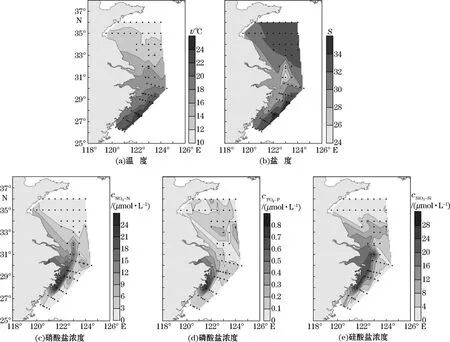
图2 调查区域表层温度、盐度、硝酸盐、磷酸盐和硅酸盐分布Fig.2 Distributions of temperature,salinity,nitrate,phosphate and silicate in the surface layer in the study area
2.2 物种组成
通过对306个浮游植物样品的分析,共鉴定浮游植物4门55属146种,其中硅藻门38属97种,甲藻门15属45种,蓝藻门2属3种,金藻门1属1种。硅藻和甲藻是调查区的主要浮游植物功能群,其中硅藻门中角毛藻属(Chaetoceros)、圆筛藻属(Coscinodiscus)和根管藻属(Rhizosolenia)物种最多,甲藻门中角藻属(Ceratium)和原多甲藻属(Protoperidinium)物种最多。生态类型方面,调查区浮游植物物种以温带近岸性和广布性物种为主,也包含部分暖水性或大洋性物种,如太阳漂流藻(Planktoniellasol)、秘鲁角毛藻(Chaetocerosperuvianus)、刺尖甲藻(Oxytoxumscolopax)和单刺足甲藻(Podolampasspinifera)等。
调查区浮游植物优势物种组成(表1):南黄海海域优势物种以链状硅藻为主,也包含部分甲藻,排名靠前的优势物种为诺氏海链藻(Thalassiosiranordenskioldii)、太平洋海链藻(Thalassiosirapacifica)、微小原甲藻(Prorocentrumminimum)和具槽帕拉藻(Paraliasulcata)等;东海海域优势物种同样以链状硅藻为主,包含部分甲藻物种,排名靠前的优势物种为柔弱伪菱形藻(Pseudo-nitzschiadelicatissima)、具槽帕拉藻(Paraliasulcata)、柔弱角毛藻(Chaetocerosdebilis)和具齿原甲藻(Prorocentrumdentatum)(同物异名:东海原甲藻Prorocentrumdonghaiense)等。可以看出,南黄海和东海海域浮游植物优势物种均以链状硅藻为主,并且2个海域前2位优势种的优势度要明显高于其他优势种;另外,相较于硅藻,甲藻在2个海域的优势度较低。

图3 调查区域断面A,B,C,D,E和F的温度、盐度和硝酸盐浓度分布Fig.3 Distributios of temperature,salinity and nitrate concentration along section A,B,C,D,E and F in the study area
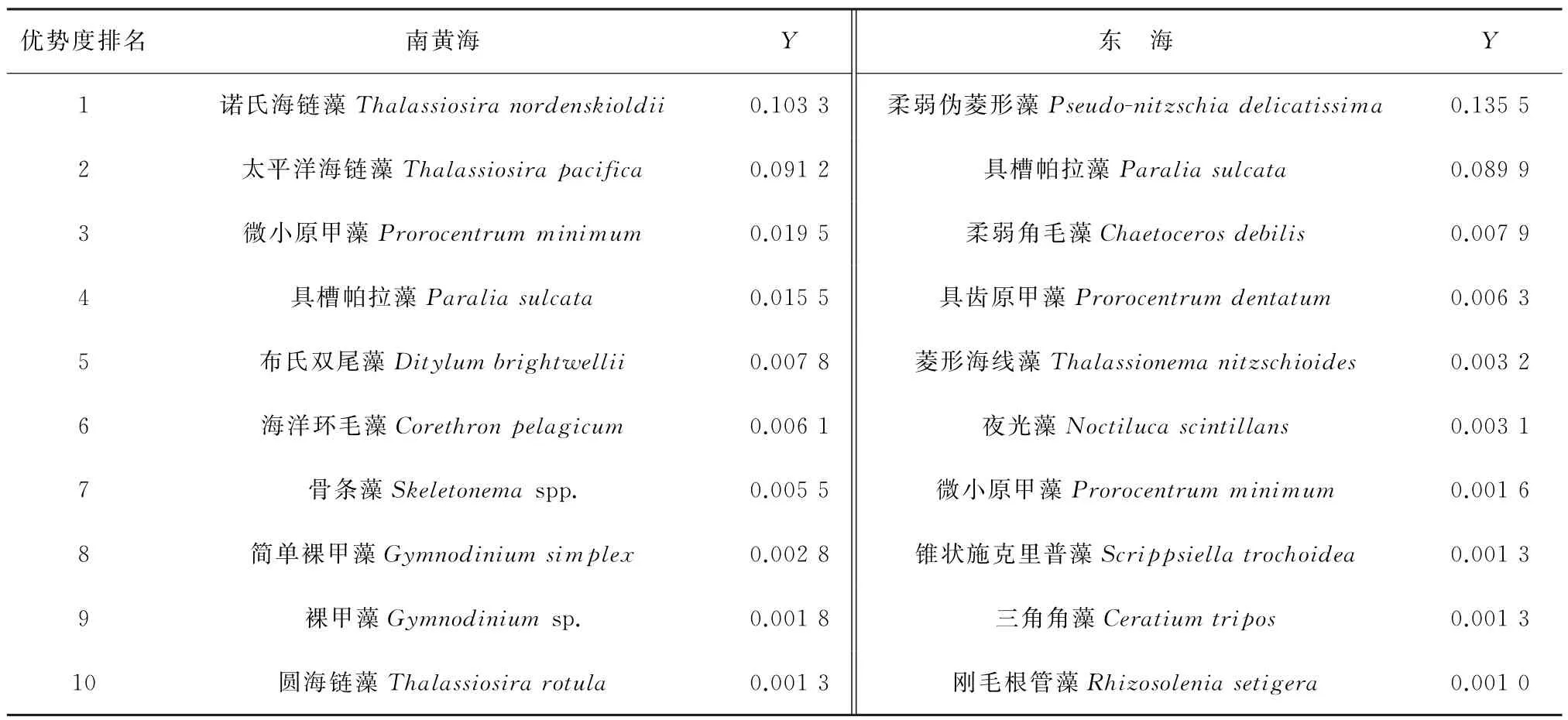
表1 调查区南黄海、东海优势度(Y)排名前10的浮游植物物种Table 1 Top 10 dominant phytoplankton species in the southern Yellow Sea and East China Sea(Ranked according to the dominance index(Y)of each species)
2.3 细胞丰度水平分布
调查区表层、20 m和50 m水层浮游植物细胞丰度分布见图4。调查区表层浮游植物总细胞丰度为(0.07~547.87)×103个/L,平均值为29.45×103个/L;硅藻细胞丰度为(0.07~544.81)×103个/L,平均值为22.45×103个/L;甲藻细胞丰度为(0.04~96.80)×103个/L,平均值为6.99×103个/L。整体来看,调查区表层有2个浮游植物细胞丰度高值区:南黄海中部区域和闽浙沿岸区域(图4a)。南黄海浮游植物细胞丰度高值集中在B断面外侧的3500-09站位,优势物种为诺氏海链藻和太平洋海链藻;闽浙沿岸区域优势物种为柔弱伪菱形藻。表层硅藻细胞丰度与浮游植物总细胞丰度的分布趋势一致(图4b);甲藻细胞丰度高值集中在南黄海中部和浙江沿岸区域(图4c),其中南黄海中部细胞丰度高值区优势物种为微小原甲藻和简单裸甲藻(Gymnodiniumsimplex),浙江沿岸区域优势物种为具齿原甲藻。调查区20 m水层浮游植物细胞丰度高值出现在南黄海中部区域,丰度高值主要由硅藻贡献,优势物种为诺氏海链藻和太平洋海链藻;另外在长江口外侧站位细胞丰度也较高,优势物种为具槽帕拉藻和骨条藻(图4d~图4e)。甲藻细胞丰度在B断面的3500-03站位较高,优势物种为微小原甲藻(图4f)。在50 m水层,浮游植物细胞丰度仍然在南黄海中部出现高值,优势物种为太平洋海链藻和诺氏海链藻(图4g);硅藻细胞丰度分布与浮游植物总细胞丰度分布一致(图4h);甲藻细胞丰度较低,最高值出现在B断面的3500-07站位,优势物种为裸甲藻(图4i)。
南黄海和东海海域主要优势物种细胞丰度在表层的分布趋势(图5)显示,在南黄海海域,诺氏海链藻、太平洋海链藻和微小原甲藻的细胞丰度高值区均集中在中部区域(图5a~图5c),具槽帕拉藻的细胞丰度高值区则集中在南黄海南部水深较浅的内侧站位(图5d)。在东海海域,柔弱伪菱形藻细胞丰度高值集中在闽浙沿岸区域,另外在DH04站位也出现高值(图5e);具槽帕拉藻细胞丰度高值区出现在长江口邻近的区域及DH06站位(图5f);柔弱角毛藻和具齿原甲藻的细胞丰度高值均集中于浙江近岸区域,分布趋势比较一致(图5h)。综上所述,南黄海海域优势物种的细胞丰度高值区集中在南黄海中部,其他区域分布较少;东海海域优势物种的细胞丰度高值区集中在长江口外侧及闽浙沿岸海域,从近岸到外海呈逐渐降低的趋势。
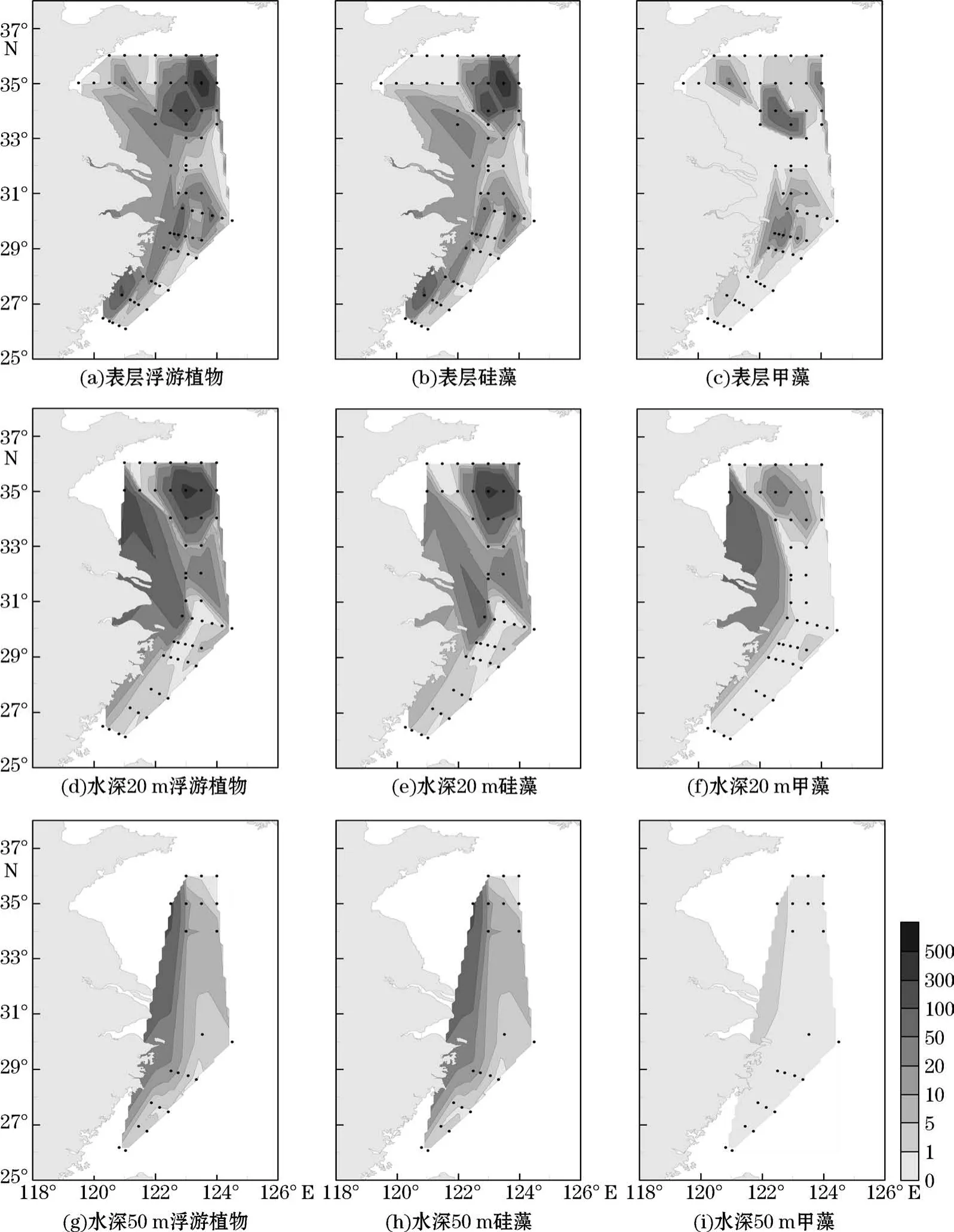
图4 调查区表层、水深20 m和50 m水层浮游植物细胞丰度(×103个·L-1)分布Fig.4 Horizontal distributions of phytoplankton cell abundance(×103 cells·L-1)in the surface,20 m and 50 m layer in the study area
2.4 细胞丰度断面分布
调查区各断面浮游植物细胞丰度分布见图6。断面A浮游植物细胞丰度高值区出现在断面外侧的3600-08站位的10~30 m水层,优势物种为布氏双尾藻(Ditylumbrightwellii)和太平洋海链藻。断面B浮游植物细胞丰度高值区出现在断面外侧的3500-08站位的20~30 m水层,优势物种为诺氏海链藻和太平洋海链藻。断面C浮游植物细胞丰度高值区出现在断面外侧的3400-08站位的30 m水层,优势物种为太平洋海链藻和菱形海线藻(Thalassionemanitzschioides);另外,位于断面中部的3400-06站位的0 m和20 m水层细胞丰度也较高,优势物种为太平洋海链藻和诺氏海链藻。可以看出,南黄海海域断面A、B和C浮游植物细胞丰度的高值均出现在外侧,并且在断面B外侧站位的10~30 m水层出现硅藻水华,最高值已超过106个/L,近岸区域细胞丰度则较低。
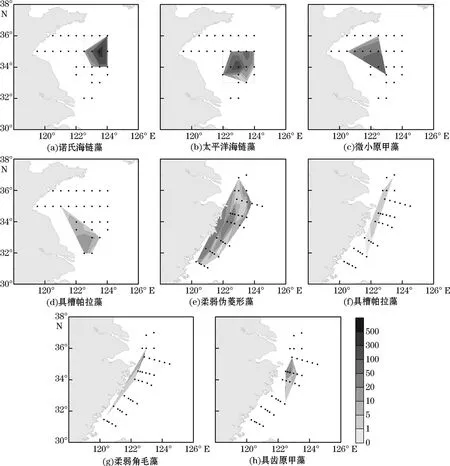
图5 调查区表层浮游植物优势物种细胞丰度(×103个·L-1)分布Fig.5 Horizontal distributions of cell abundance(×103 cells·L-1)of dominant species in the surface water in the study area
断面D浮游植物细胞丰度高值区出现在近岸DH-01站位的0~20 m水层,优势物种为柔弱角毛藻和具齿原甲藻。断面E浮游植物细胞丰度高值区出现在近岸Zb7站位的表层,优势物种为柔弱伪菱形藻。断面F浮游植物细胞丰度高值区出现在近岸DH8-1站位的表层,优势物种为柔弱伪菱形藻和微小细柱藻(Leptocylindrusminimus)。东海海域3条断面的细胞丰度高值区均集中在近岸区域,并呈从近岸到外海、从表层到底层呈逐渐降低的趋势。
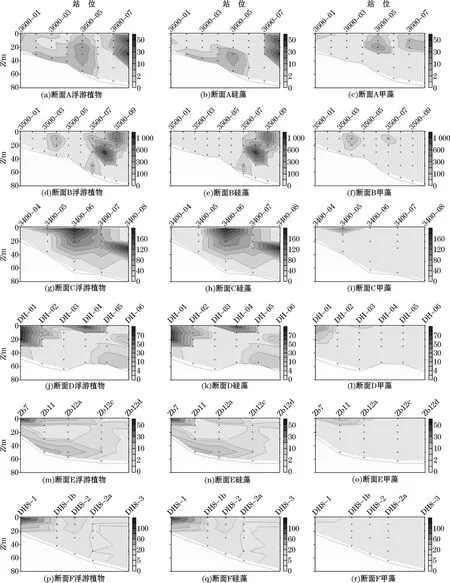
图6 调查区各断面浮游植物细胞丰度(×103个·L-1)分布Fig.6 Transectional distributions of phytoplankton cell abundance(×103 cells·L-1)in the study area
2.5 多样性指数和均匀度指数
调查区表层水体浮游植物群落香侬-威纳多样性指数(图7)显示表层为1.05~4.86,平均值为2.48,高值区出现在调查区的北部近岸站位,低值区则集中在南黄海南部区域。调查区表层Pielou均匀度指数(图7)为0.02~0.97,平均值为0.57,最低值出现在南黄海中部的3330-06站位,仅为0.02,优势物种为微小原甲藻,其细胞丰度占到该站位总细胞丰度的99%以上;高值区集中在南黄海北部近岸区域和东海外侧区域。
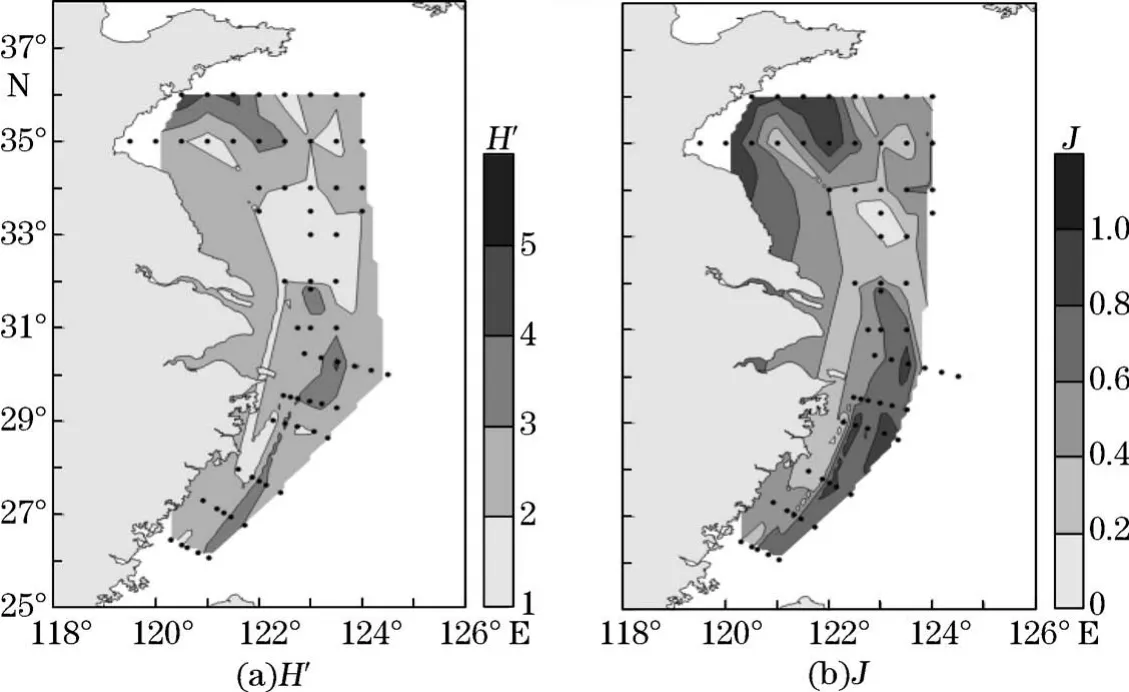
图7 调查区海域表层浮游植物群落Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)分布Fig.7 Distributions of Shannon-Wiener diversity index(H′)and Pielou′s eveness index(J)of phytoplankton community in the surface layer in the study area
2.6 浮游植物细胞丰度与环境因子的关系
调查区表层浮游植物细胞丰度与环境因子相关性分析(表2)表明,南黄海浮游植物细胞丰度与SiO3-Si和PO4-P浓度呈现显著正相关,而与其他环境因子相关性不明显;硅藻细胞丰度与SiO3-Si和NO3-N浓度呈现显著正相关,甲藻细胞丰度则与PO4-P和NO3-N浓度呈现显著负相关。东海浮游植物细胞丰度与温度和盐度呈现显著负相关,与SiO3-Si,NH4-N和NO3-N浓度呈现显著正相关;硅藻细胞丰度与温度和盐度呈现显著负相关,与NO3-N浓度呈现显著正相关;甲藻细胞丰度与盐度呈现显著负相关,与SiO3-Si和NO3-N浓度呈现显著正相关。

表2 调查区域表层浮游植物细胞丰度与环境因子的Pearson相关性分析Table 2 Pearson correlation analysis between phytoplankton cell abundance and environmental factors in the surface layer in the study area
3 讨 论
3.1 与历史资料比较
我们的调查结果与同时期历史调查资料的比较(表3)说明,南黄海海域本次调查主要优势种为诺氏海链藻、太平洋海链藻、微小原甲藻和具槽帕拉藻等(表1),其中2种优势种与ZHANG S等在2014年的调查结果[16]一致。2009年田伟和孙军调查发现具齿原甲藻是首要优势种,主要集中分布在南黄海南部,其次是柔弱伪菱形藻和具槽帕拉藻等[17],即总体上南黄海春季浮游植物优势物种还是以链状硅藻为主。东海海域本次调查主要优势种为柔弱伪菱形藻、具槽帕拉藻、柔弱角毛藻和具齿原甲藻等(表1),历史调查资料主要优势种则以具齿原甲藻和骨条藻居多(表3),这可能跟调查范围的差异有关。历史调查资料的采样范围多集中在长江口及其邻近海域,如赵冉等的调查范围为(121°00′~123°30′E,30°30′~32°30′N)[11],孙军和田伟的调查范围为(121°00′~123°50′E,30°50′~32°00′N)[18]。这些区域均是春季发生原甲藻和骨条藻水华的常见区域[19],而本次调查在长江口邻近区域并没有设置调查站位(图1)。
调查中南黄海和东海海域浮游植物优势物种均以链状硅藻为主(表1)。于其他类群比较,链状硅藻物种具有更好的环境适应性,在营养盐丰富的陆架区较容易成为优势类群[20]。我们认为,首先群体形态可以增加细胞的浮力,使细胞更容易抵御下沉,从而更好地停留在上层水体进行光合作用;其次相较于单细胞,群体形态的细胞在下沉过程中会有更大的细胞营养盐通量[21];另外群体形态的细胞可以有效抵御浮游动物的摄食[22]。因此在营养盐较丰富且水动力复杂的黄、东海陆架区域,链状硅藻较易成为优势类群。
3.2 浮游植物细胞丰度与环境因子的相关性
从浮游植物细胞丰度的平面分布来看,调查区春季细胞丰度高值区出现在南黄海中部区域和闽浙沿岸区域(图4a),这与调查区营养盐的分布趋势相一致(图2c~图2e)。研究发现,长江冲淡水和黑潮次表层水的涌升是黄、东海海域营养盐的2个主要来源[24]。春季在季风影响下,长江冲淡水有分支南下,形成闽浙沿岸的营养盐高值区(图2c~图2e);同时在南黄海中部区域,从断面A和B的营养盐分布可以看出次表层水的涌升(图3c和图3f),为该区域上层水体带来充足的营养盐,这与前人的研究结果相似[25-26]。因此,2个区域充足的营养盐供应导致了浮游植物细胞丰度的高值现象。
南黄海近岸区域浮游植物细胞丰度较低(图4a)。断面分布表明断面A和B近岸站位有硝酸盐浓度的高值(图3c和图3f),但同时水体盐度垂直混合也比较强烈(图3b和图3e)。程君等研究发现,春季南黄海近岸区域由于水深较浅、水体上下混合均匀,浊度较大,透明度不高,浮游植物的生长受到光限制[27]。因此虽然该区域营养盐浓度较高,浮游植物生长易受到光限制而导致细胞丰度较低。另外,东海海域外侧区域浮游植物细胞丰度随着等深线的加深迅速降低(图4a),这与营养盐浓度的分布趋势也比较一致(图2c~图2e)。研究还表明,东海水层50 m以深的陆架区,营养盐含量较低,浮游植物的生长易受到营养盐的限制[24]。
浮游植物细胞丰度与环境因子的Pearson相关性分析表明,南黄海海域硅藻细胞丰度跟SiO3-Si和NO3-N浓度呈显著正相关,甲藻细胞丰度则跟PO4-P和NO3-N浓度呈显著负相关(表2),这种差异可能跟硅藻和甲藻的生存策略差异有关。硅藻的营养型属于完全自养型,在营养盐充足的环境中可以凭借生长优势快速成为优势类群[28],研究断面A,B,C的硅藻细胞丰度的高值区也基本与营养盐浓度的高值区相吻合(图3c、图3f、图3i、图6b、图6e和图6h),因此硅藻细胞丰度与营养盐浓度呈现显著的正相关,在高营养盐浓度区域占据优势地位。与硅藻相比,甲藻则更能适应寡营养的环境。研究发现,很多甲藻物种具有异养营养行为,在寡营养环境中可以通过摄食其他小型生物补充自身营养需要[29]。因此当营养盐浓度较低时,甲藻通过异养方式仍具有一定的生存优势,而硅藻则往往由于营养盐限制而导致生物量和优势度降低,这便导致
两者在与营养盐的相关性上出现了差异(表2)。在东海海域,硅藻和甲藻细胞丰度则均与盐度呈显著负相关,与NO3-N浓度呈显著正相关(表2),这与文斐等的研究结果[30]基本一致。从细胞丰度的分布可以看出,硅藻和甲藻的细胞丰度高值均出现在受长江冲淡水影响明显的闽浙沿岸区域(图4b和图4c),邻近区域水体以低盐度和高营养盐浓度为显著特点(图2b~图2e)。因此,在调查区盐度越低、营养盐浓度越高的区域,浮游植物的细胞丰度也往往越高。东海甲藻与SiO3-Si的相关性比硅藻好(表2),这是因为甲藻丰度的高值区主要集中在长江口外及浙江沿岸区域(图4c),跟SiO3-Si浓度高值区的分布比较一致(图2e)。但是,这并不能说明是当地海域高的SiO3-Si浓度支撑了甲藻的生长,因为NO3-N在该区域的浓度也较高(图2c),很有可能是NO3-N的高浓度造成了该区域甲藻的高丰度。与甲藻不同,东海海域硅藻的丰度高值出现在南侧F断面的近岸区域(图4b),该区域SiO3-Si的浓度并不算太高(图2e),在SiO3-Si浓度最高的长江口外和浙江近岸区域硅藻丰度则相对较低,所以导致硅藻与SiO3-Si的相关性并不显著。
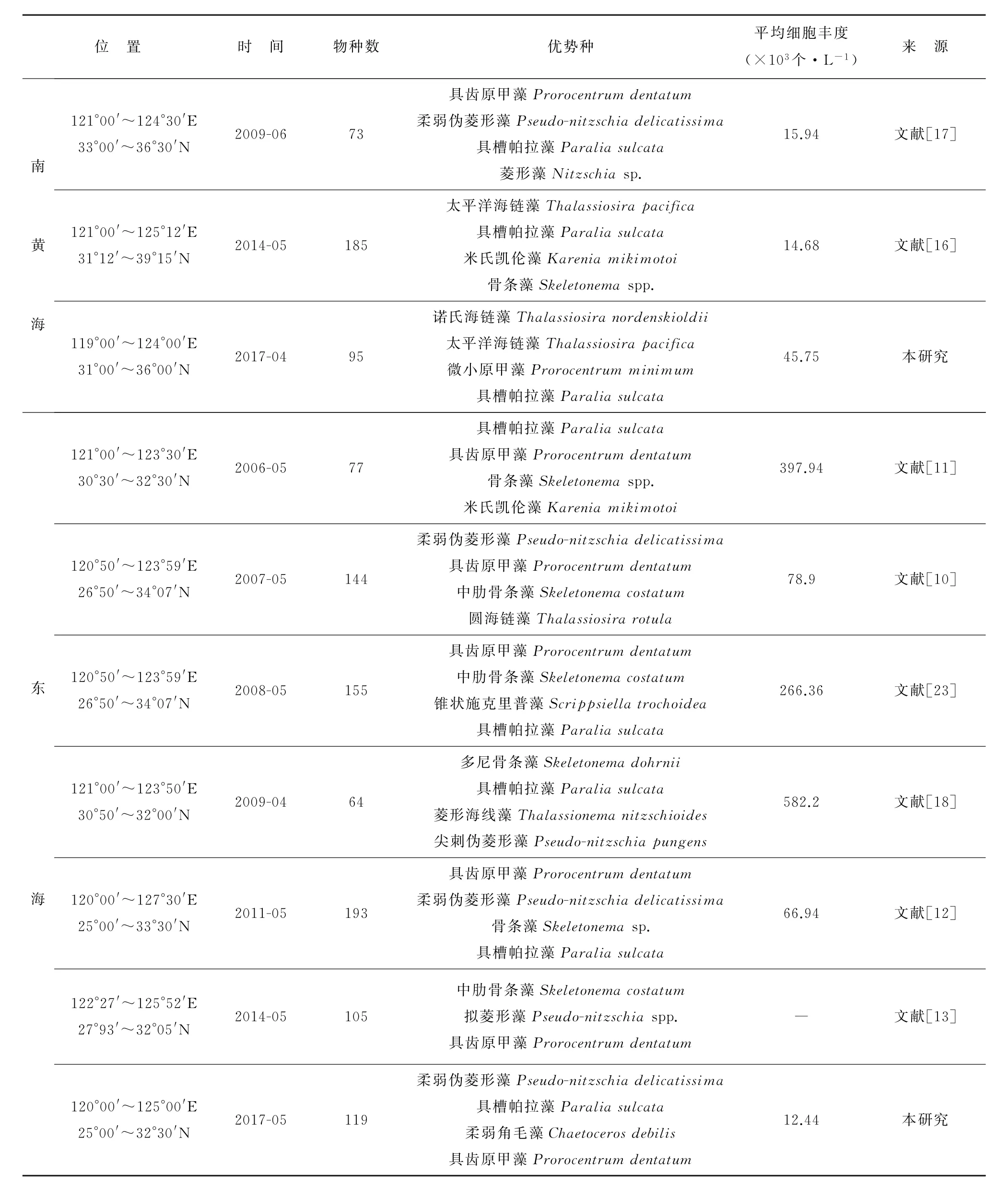
表3 调查区域物种丰富度、优势种组成及细胞丰度与历史水采方法调查资料的比较Table 3 Comparison of species richness,dominant species and cell abundance among historical data in the survey area
3.3 南黄海中部春季水华
春季水华在温带和亚极地海域是一个常见的现象,很大程度上影响着当地海域渔业生产量及生源要素的循环[31]。在南黄海中部,几乎每年春季都会爆发浮游植物水华[32]。本次调查中在南黄海中部也发现了硅藻水华,优势物种为诺氏海链藻和太平洋海链藻,其中在断面B的3500-08站位的30 m水层细胞丰度超过了106个/L(图6e)。水华发生的位置集中在南黄海水深超过50 m的中部区域(图6b、图6e和图6h),这与前人的观测结果也一致[33-34]。
从细胞丰度的垂直分布来看,水华区域浮游植物细胞丰度的最高值出现在各站位的10~30 m水层(图6a~图6c)。以断面C的3400-08站位为例,浮游植物细胞丰度最大值出现在30 m水层(图6g)。该站位表层的NO3-N浓度为0.36μmol/L,PO4-P浓度为0.087μmol/L。相关的营养盐吸收动力学研究发现,浮游植物生长所需的NO3-N浓度阈值为1.0μmol/L,PO4-P浓度阈值为0.1μmol/L[35]。因此,该区域表层水体的营养盐浓度较低,不适宜浮游植物的生长,导致浮游植物细胞丰度的最大值不是出现在表层,而是出现在营养盐浓度较高的30 m层。我们认为,春季南黄海中部次表层水的涌升携带来充分营养盐,支持了当地海域浮游植物的生长,形成水华;不过次表层水的涌升往往只能达到10~30 m的水层,导致浮游植物细胞丰度的最高值集中在水华区域的次表层。
调查中南黄海中部水华区域的优势物种为诺氏海链藻、太平洋海链藻和菱形海线藻等硅藻物种(表4)。营养盐数据分析表明,水华站位的Si/N比值均明显大于1。
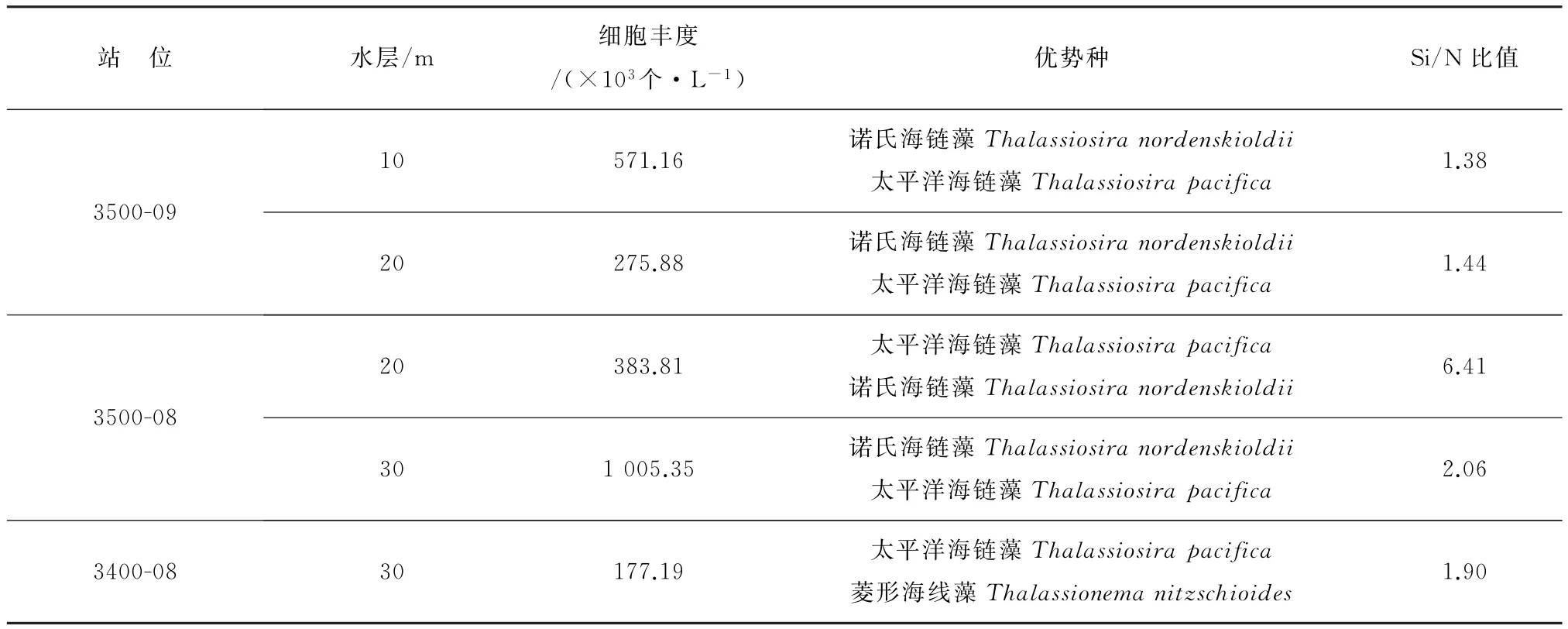
表4 南黄海中部水华区域各站位细胞丰度、优势物种及Si/N比值Table 4 Cell abundance,dominant species and Si/N values at the bloom stations in the central southern Yellow Sea
根据Redfield理论,Si在南黄海中部不是硅藻生长的限制性因素,这也与前人的研究结论[36]一致。因此,在硅酸盐供应充足的情况下,硅藻可以迅速繁殖,成为南黄海中部春季水华的优势物种。鉴于硅藻细胞死亡后具有较高的沉降速率,形成碳汇。因此,南黄海中部春季硅藻水华及后续的浮游植物碳沉降在该海域碳汇中的贡献值得探索。
4 结 论
我们用水采样品系统地研究2017年春季南黄海、东海海域的浮游植物群落结构,对其与环境因子关系进行了分析:
对306个浮游植物样品的分析共鉴定浮游植物4门55属146种,硅藻和甲藻是调查区域的主要浮游植物功能群。生态类型方面,调查区浮游植物物种以温带近岸性和广布性物种为主,也包含部分暖水性或大洋性物种。南黄海和东海海域浮游植物优势物种均以链状硅藻为主,其中南黄海主要优势种为诺氏海链藻(Thalassiosiranordenskioldii)、太平洋海链藻(Thalassiosirapacifica)、微小原甲藻(Prorocentrumminimum)和具槽帕拉藻(Paraliasulcata)等,东海主要优势种为柔弱伪菱形藻(Pseudo-nitzschiadelicatissima)、具槽帕拉藻(Paraliasulcata)、柔弱角毛藻(Chaetocerosdebilis)和具齿原甲藻(Prorocentrumdentatum)等。调查区浮游植物细胞丰度存在2个高值区:南黄海中部区域和闽浙沿岸区域。断面分布上,南黄海浮游植物细胞丰度的高值均出现在外侧区域,并且在断面B外侧站位的10~30 m水层发现硅藻水华;东海浮游植物细胞丰度高值区则集中在近岸区域,且从近岸到外海、从表层到底层逐渐降低。调查区浮游植物细胞丰度的分布趋势主要受营养盐分布影响所致:长江冲淡水南下分支和南黄海次表层水的涌升分别为闽浙沿岸和南黄海中部带来充足的营养盐,从而导致了2个区域浮游植物细胞丰度的高值。调查中发现南黄海中部水华区域的浮游植物优势物种以链状硅藻为主,鉴于其较高的沉降速率和碳输出效率,南黄海中部春季硅藻水华在该海域碳汇中的贡献值得后续的深入探索。
致谢:中国科学院海洋研究所“科学三号”科研调查船提供采样平台,中国科学院海洋研究所研究生刘华健、田梓杨和李庆洁进行现场采样。
