苏州东山茶树种质资源遗传多样性的ISSR分析
2020-08-04李浩宇杨大强王利芬张忠新郑俊华韩兆岚朱旭君房婉萍尹江海
李浩宇,杨大强,王利芬,张忠新,郑俊华,韩兆岚,朱旭君,房婉萍,尹江海
(1.苏州市吴中区东山镇农林服务站,江苏 苏州 215107;2.苏州大学金螳螂建筑学院,江苏 苏州 215325;3.南京农业大学茶叶科学研究所,江苏 南京 210095;4.苏州农业职业技术学院,江苏 苏州 216000)
苏州是我国最古老的茶区之一,早在明朝便有苏州茶为“天下之冠”,以及“饮遍天下”之说。苏州地区茶树(Camellia sinensis)主要以来自洞庭东、西山地区的地方群体种为代表。以洞庭山茶树为原料制作的洞庭碧螺春是历史名茶,并以独特的自然品质和悠久的帝王传说闻名海内外。根据朱鸿寿等[1]的记载,洞庭山茶树品种较杂,品种来源无从考证,除白茶树外,还有少数紫芽种,大部分以中小型灌木为主。各单株间性状差异较大,有早、晚芽,有中、小叶,有茸毛多少,有紫色芽叶或碧绿芽叶;内在生化成分也有差异。东、西山群体种以‘柳叶条’、‘酱板头’、‘柴茶’等小叶茶树品种为代表,这些品种适制洞庭碧螺春,是目前洞庭碧螺春原产地主导品种。除群体种茶树品种外,还有福鼎大白茶、槠叶种、福云5号、福云6号、鸠坑种、迎霜、浙农113、乌牛早、龙井43等,其中大多数茶树品种并不适合制作洞庭碧螺春,所制茶叶质量无论外形条索、色泽、香气还是滋味、汤色,都不具备洞庭碧螺春原产地茶叶外形“条索纤细,卷曲成螺,茸毛遍体,银绿隐翠”及内质“汤色碧绿,清香高雅,入口爽甜”的传统品质风格特色,不利于洞庭碧螺春品质声誉[2]。因此,从适制洞庭碧螺春的东、西山群体种中选育出生物学性状优异的茶树无性系良种作为洞庭碧螺春未来的主导茶树品种是亟待解决的关键问题之一。
简单重复间序列标记 (Inter-Simple Sequence Repeat,ISSR)是一种基于微卫星序列发展起来的十分有效的分子标记技术[3],与AFLP、RAPD和SSR等其他分子标记技术相比,它具有引物设计简单、遗传多样性丰富、稳定性强和重复性好等优点,适用于亲缘关系较近的材料[4—8]。ISSR分子标记广泛应用于茶树种质资源鉴定和亲缘关系分析[4—5,9—18]。苏州洞庭地区茶树栽培品种或品系十分丰富,但没有当地选育的、适制洞庭碧螺春的茶树无性系品种,导致洞庭地区茶树品种无法统一,洞庭碧螺春茶叶质量难于一致[2]。为探明洞庭地区已有茶树品种间的遗传多样性,本研究采用ISSR标记对洞庭地区的茶树种质资源进行分子标记研究,以期为充分利用茶树资源和创新提供依据。
1 材料与方法
1.1 材料
供试茶树种质材料51份,具体茶树品种(材料)和来源见表1。取样地点为苏州东山和无锡。取1芽2叶,液氮速冻后于-80 ℃冰箱保存备用。
1.2 方法
1.2.1 DNA提取
采用改良CTAB法提取茶树叶片基因组DNA[17]。取茶树1芽2叶用液氮迅速研磨至细末,加入1 mL 65 ℃预热的DNA提取缓冲液(100 mmol·L-1Tris·HCl,50 mmol·L-1EDTA,1.5 mol·L-1NaCl,2%十六烷基三乙基溴化铵,1%聚乙烯吡咯烷酮,2%β-巯基乙醇,pH 8.0),转入2 mL离心管。65 ℃水浴90 min,每15 min上下轻轻颠倒混匀;10 000×g离心10 min,取上清,加入等体积氯仿:异戊醇(24:1)混合液,轻轻颠倒 10 min 后静置3 min;10 000×g离心15 min,取上清;再加入等体积氯仿:异戊醇(24:1)抽提一次,离心后取上清450 μL,加入2/3体积预冷的异丙醇混合均匀,置于-20 ℃冰箱中30 min;10 000×g离心10 min;弃上清,用1 mL 75%乙醇洗涤沉淀2次,无菌风吹干,加50 μL无菌双蒸水溶解。提取所有样品的基因组DNA,用1% (W/V)琼脂糖凝胶电泳检测DNA纯度。
1.2.2 ISSR-PCR扩增和产物检测
茶树 ISSR-PCR反应体系总体积为20 μL,PCR组分:2 μL 10×PCR buffer (不含Mg2+),1.6 μL Mg2+(25 mmol·L-1),1.6 μL dNTP (2.5 mmol·L-1),引物0.8 μL (10 mmol·L-1),0.2 μLTaq酶 (5 U·μL-1),1 μL模板DNA(50 ng)。扩增程序:94 ℃预变性 7 min;94 ℃变性1 min,退火45 s,72 ℃延伸 90 s,35个循环; 72 ℃延伸 7 min。ISSR-PCR引物来自加拿大哥伦比亚大学(UBC)提供的100条序列,结合相关研究报道[7,19—22],筛选出12条引物(表2),由上海生工生物工程有限公司合成。PCR产物用1.5%琼脂糖凝胶120 V电泳1 h,凝胶成像系统检测。
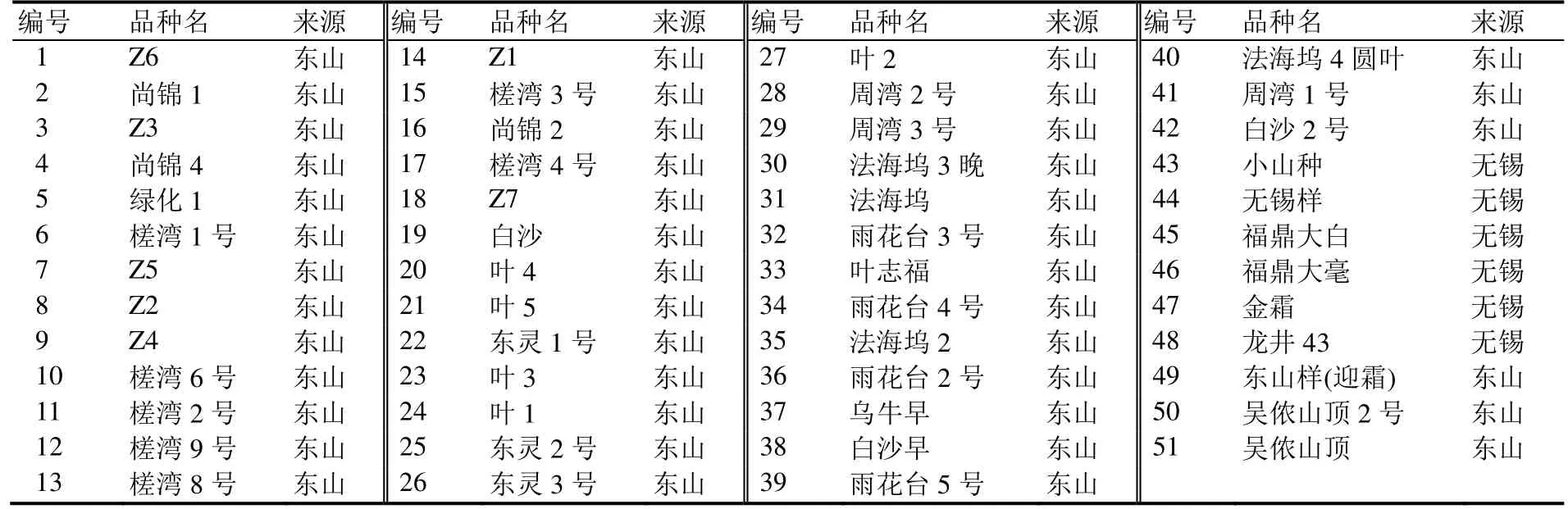
表1 供试材料Table 1 Materials used for ISSR analysis

表2 ISSR引物Table 2 Information of 12 pairs of ISSR primer
1.2.3 数据处理与分析
将凝胶成像的图片采用人工计数,同一位置有条带记为1,无则记为0,应用NTsys-pc2.01对供试材料(品种)进行聚类分析,先用Similarity程序的Qualitative data计算相似系数矩阵;再利用Clustering程序的SHAN计算,聚类分法选用UPGMA。
2 结果与分析
2.1 ISSR扩增结果
对12条引物进一步ISSR扩增筛选,选出 6条重现性好、多态性明显的引物对51个茶树品种的基因组DNA进行扩增,共扩增出76条DNA条带,其中多态性条带70条,平均多态性频率达92.11%(表3);其中UBC815引物扩增的条带为14条,多态性频率100.00%。所有引物扩增的条带大小在100~2000 bp之间,尤其以300~1500 bp居多。图1为引物UBC835扩增不同茶树材料得到的多态性条带。

表3 供试材料ISSR扩增结果Table 3 ISSR amplification result of selected primers

图 1 引物UBC835对茶树基因组DNA的ISSR PCR扩增结果Fig.1 Amplified results using primer UBC835
2.2 遗传相似性系数
利用ISSR所获得的76条扩增条带计算材料间的遗传相似性系数(GS值)。结果表明,51份茶树种质间的GS值为0.37~0.89。其中,白沙和东灵1号GS值最大,遗传相似程度最高,遗传距离最近;吴侬山顶2号和尚锦4之间的GS值最小,遗传相似程度最低,遗传距离最远(图2)。
2.3 聚类分析
利用ISSR标记数据计算材料间的遗传相似性系数矩阵,采用UPGMA法构建茶树材料间的遗传关系聚类图。聚类图在相似系数为0.57处可划分为2大组(图2)。第I组包括27份材料,组内又可划分4个亚组(相似系数0.67处)。第1亚组包含Z5、Z6、槎湾1号、周湾3号、吴侬山顶2号、法海坞3晚和法海坞2共7份材料;第2亚组为Z3;第3亚组包含尚锦1、金霜、福鼎大白、Z4、东灵3号和雨花台4号6份材料;第4亚组有槎湾2号、槎湾6号、槎湾9号、Z1、尚锦2、叶2、东灵2号、乌牛早、福鼎大毫、法海坞、小山种、无锡样和叶志福共13份材料。
第Ⅱ组包括24份材料,组内又可划分4个亚组(相似系数0.64处)。第1亚组只有1份材料,为尚锦4;第2亚组有12份材料,分别为绿化1、Z7、白沙、东灵1号、雨花台5号、东山样(迎霜)、Z2、叶5、周湾2号、雨花台3号、白沙早和法海坞4圆叶;第3亚组共有9份材料,分别是槎湾3号、槎湾4号、槎湾8号、龙井43、叶1、叶3、叶4、吴侬山顶和周湾1号;第4亚组有2份材料,即雨花台2号和白沙2号。
由图2可知,当相似性系数为0.57时,所有茶树材料都属于一个居群,说明苏州51个种质资源有较高的亲缘性,当相似性系数为0.64时可分为8个居群。其中,部分来自同一品种的不同品系在同一亚组且位置相邻,说明有较高的遗传相似性系数。

图2 供试的51份茶树材料遗传关系树状图Fig.2 The dendrogram of 51 tea cultivars based on UPGMA clustering method
3 讨论
聚类分析显示,苏州茶树种质资源中部分来自相同地区的种质能与其他地区的聚类在同一亚组,还有部分来自同一品种的不同品系有较高的遗传相似性系数,说明茶树种质资源间存在较复杂的亲缘关系。本研究从12条ISSR引物中筛选出6条多态性高、分辨力强的引物,对51份来自苏州洞庭地区的茶树种质DNA进行扩增,共扩增出76条条带,其中多态性条带70条,多态性比例达92.11%。此外,遗传相似性系数介于0.37~0.89,宽于福建53份茶树资源[23]、云南134份茶树资源[13]、我国茶树无性系品种[24]、印度栽培品种[25],表明苏州茶树种质资源间遗传基础较宽,遗传背景差异较大。白沙和东灵1号遗传相似性系数最高,为0.89,表明亲缘关系较近,可能由于种植位置较近等环境因素或亲本相似;吴侬山顶2号和尚锦4之间的遗传相似性系数为0.37,亲缘关系较远。本研究表明ISSR分子标记是评估苏州地区茶树种质资源遗传多样性的有效方法,同时也验证其具有稳定性强、操作简单、遗传多样性丰富和多态性高等特点[26—28]。UPGMA聚类分析表明,在遗传相似系数为0.57的水平上供试材料可划分为两大类,大部分在同一亚组的茶树材料来源于同一地区,表明亲缘关系远近除了受遗传背景影响外,还受种植环境带来的基因改变的影响,这与林伟东等[10]的研究结果类似。
本研究表明,苏州地区的茶树具有丰富的遗传多样性,茶树分布广泛。其遗传多样性高,与不同茶树产区特有的生态环境,以及茶树长期异花授粉造成的遗传背景复杂等因素有关。
茶树本身为高度杂合体,又是异花授粉植物,在长期杂交演化过程中产生大量不连续变异[16,29—30]。同时,长时间移种后新环境对种质的遗传信息也可能产生一定影响。本研究通过ISSR分子标记从分子水平揭示了苏州茶树种质资源间的亲缘远近,为苏州地区茶树品种选育提供理论基础。
